Relationship between Photosynthetic Capacity and Microcystin Production in Toxic Microcystis Aeruginosa under Different Iron Regimes



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cyanobacteria Cultivation and Experimental Setup
2.2. Determination of Cyanobacterial Growth and Photosynthetic Capacity
2.3. Extraction and Measurement of MC–LR
2.4. Generalized Additive Model (GAM) Construction and Statistical Analysis
3. Results and Discussion
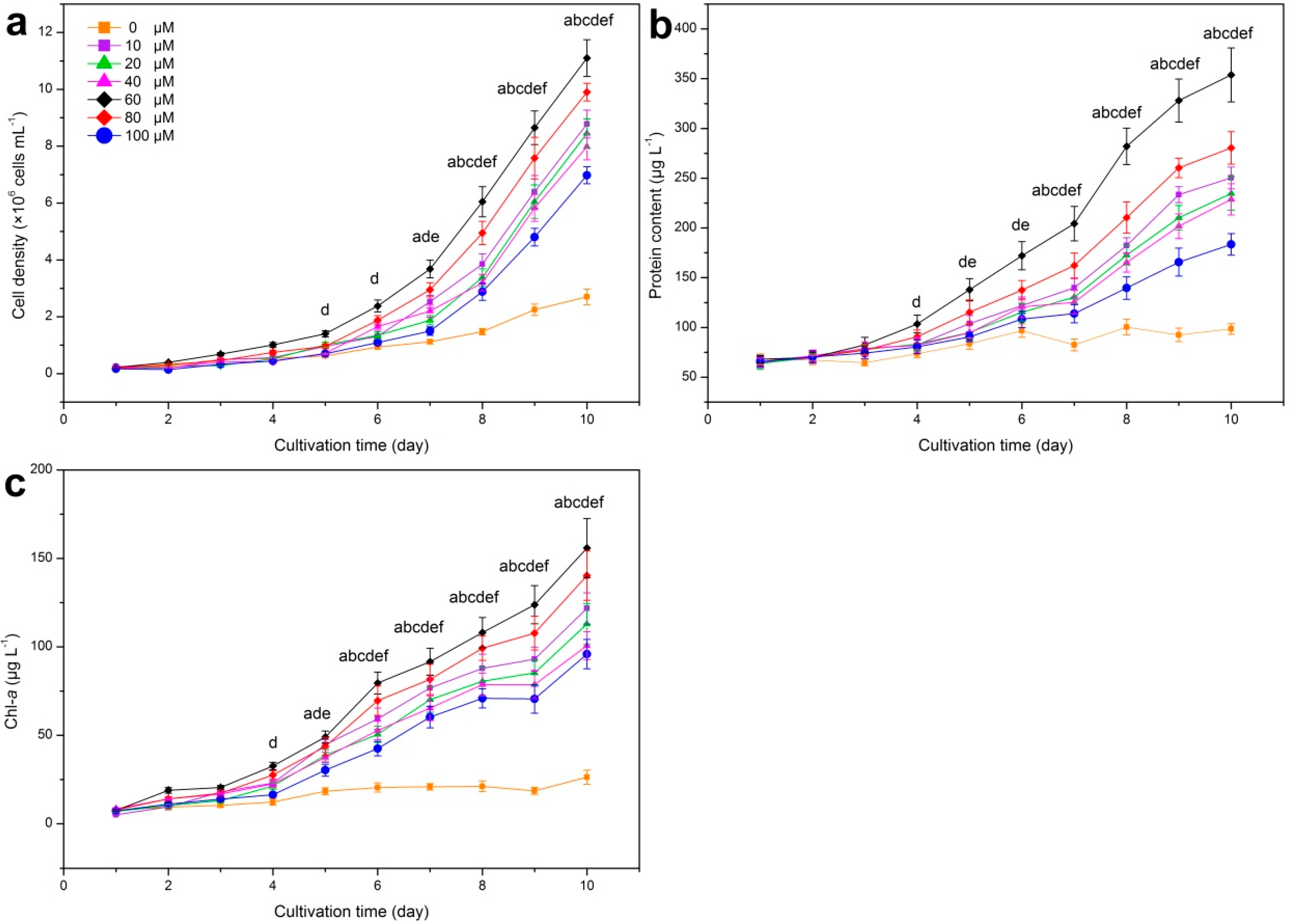
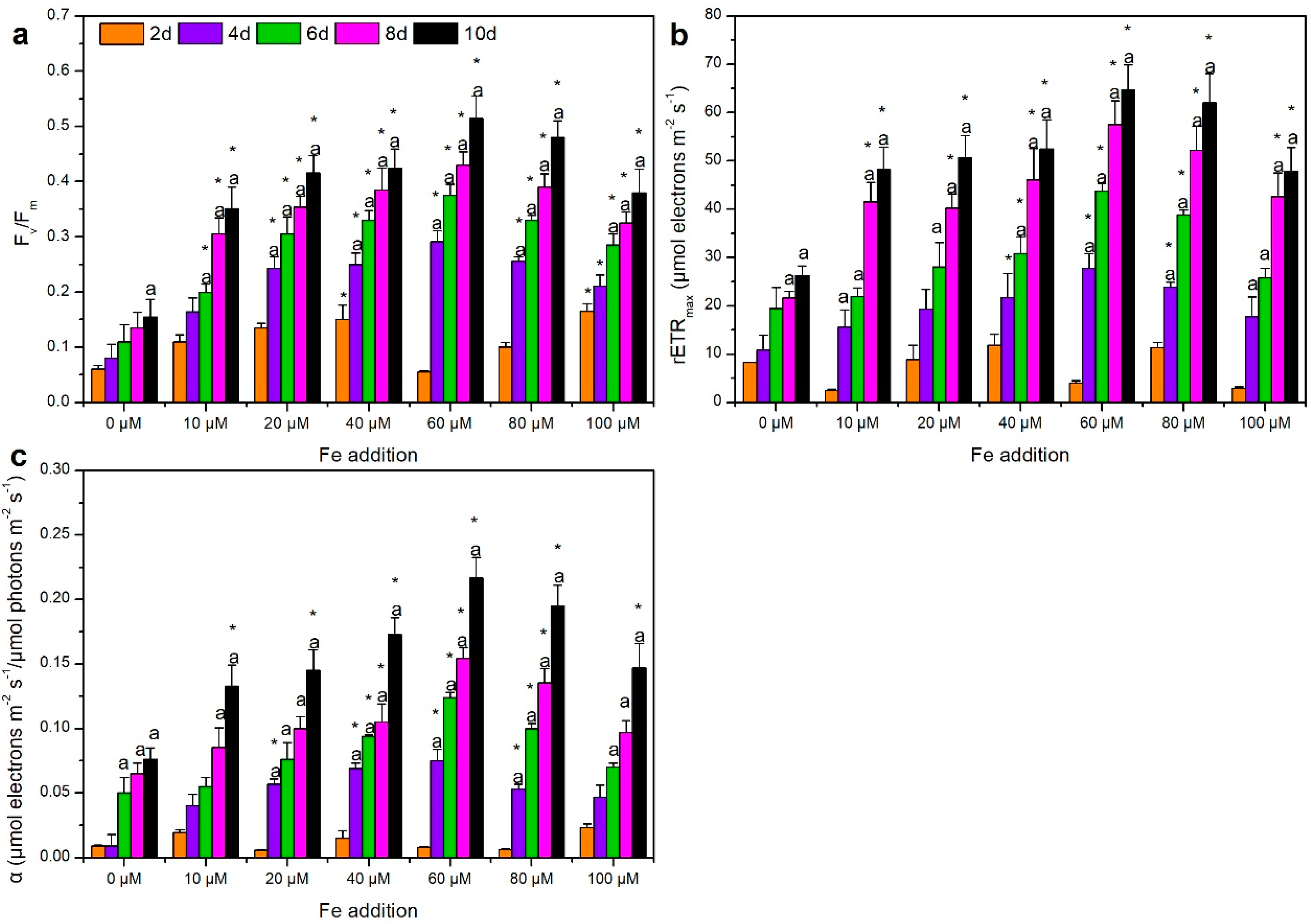
3.1. Response of Growth and Photosynthetic Capacity of M. aeruginosa to Iron Variation
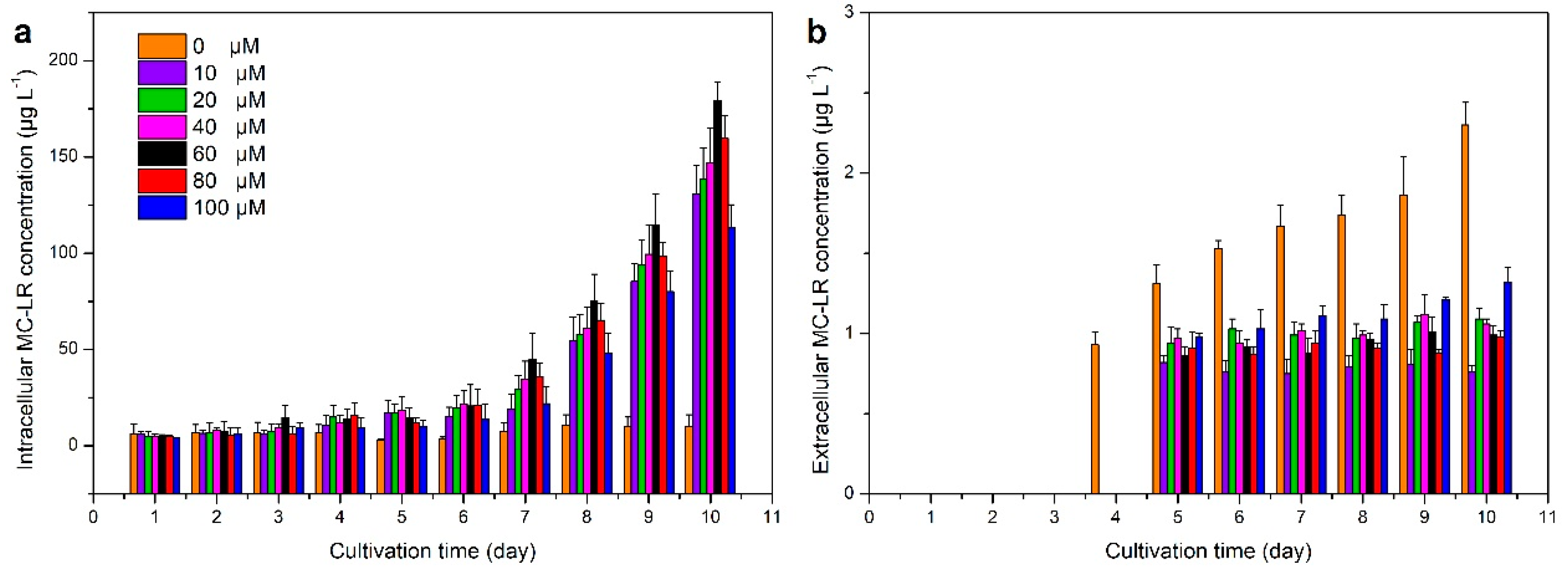
3.2. Iron Effect on MC Production in M. aeruginosa
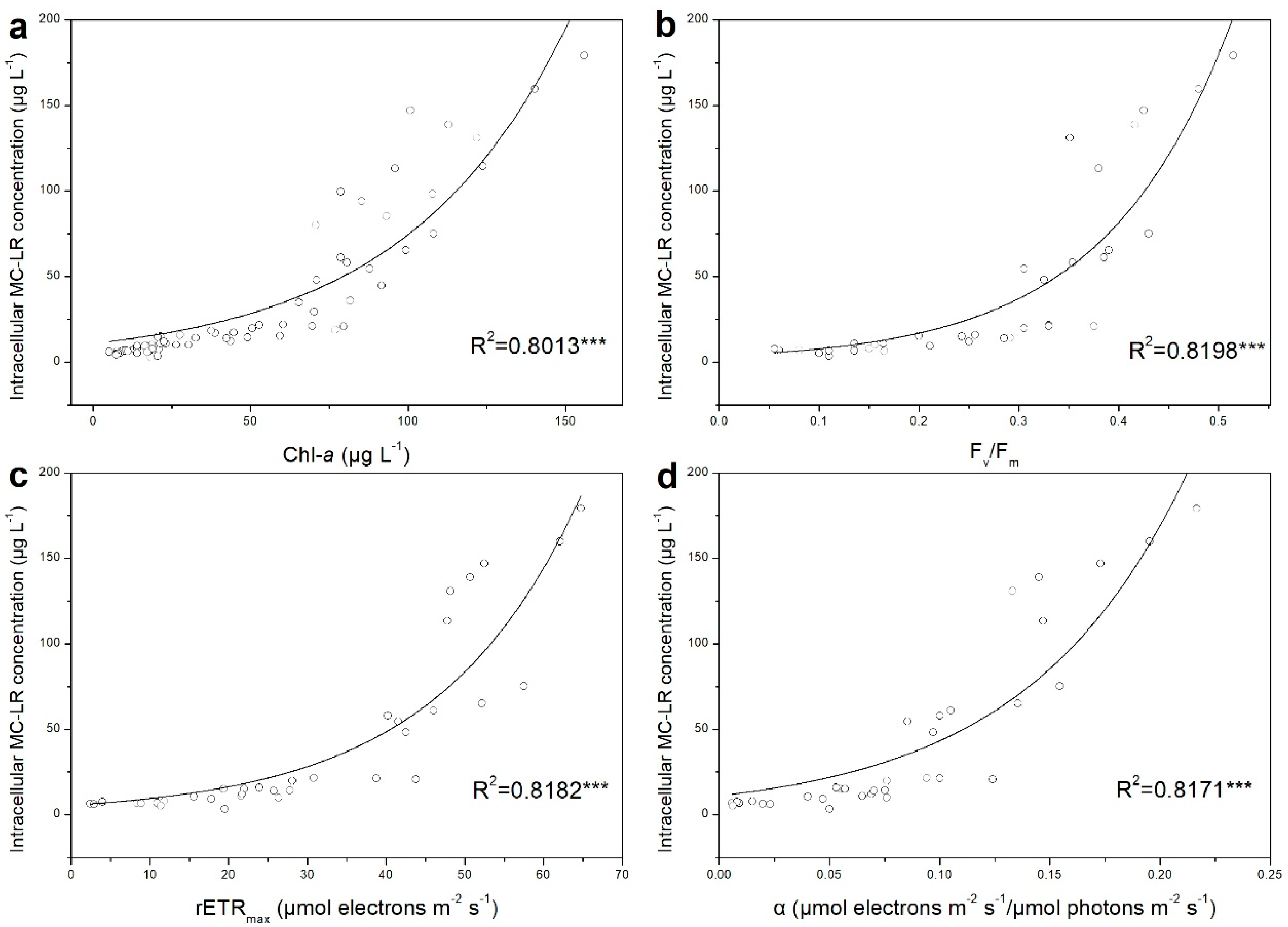
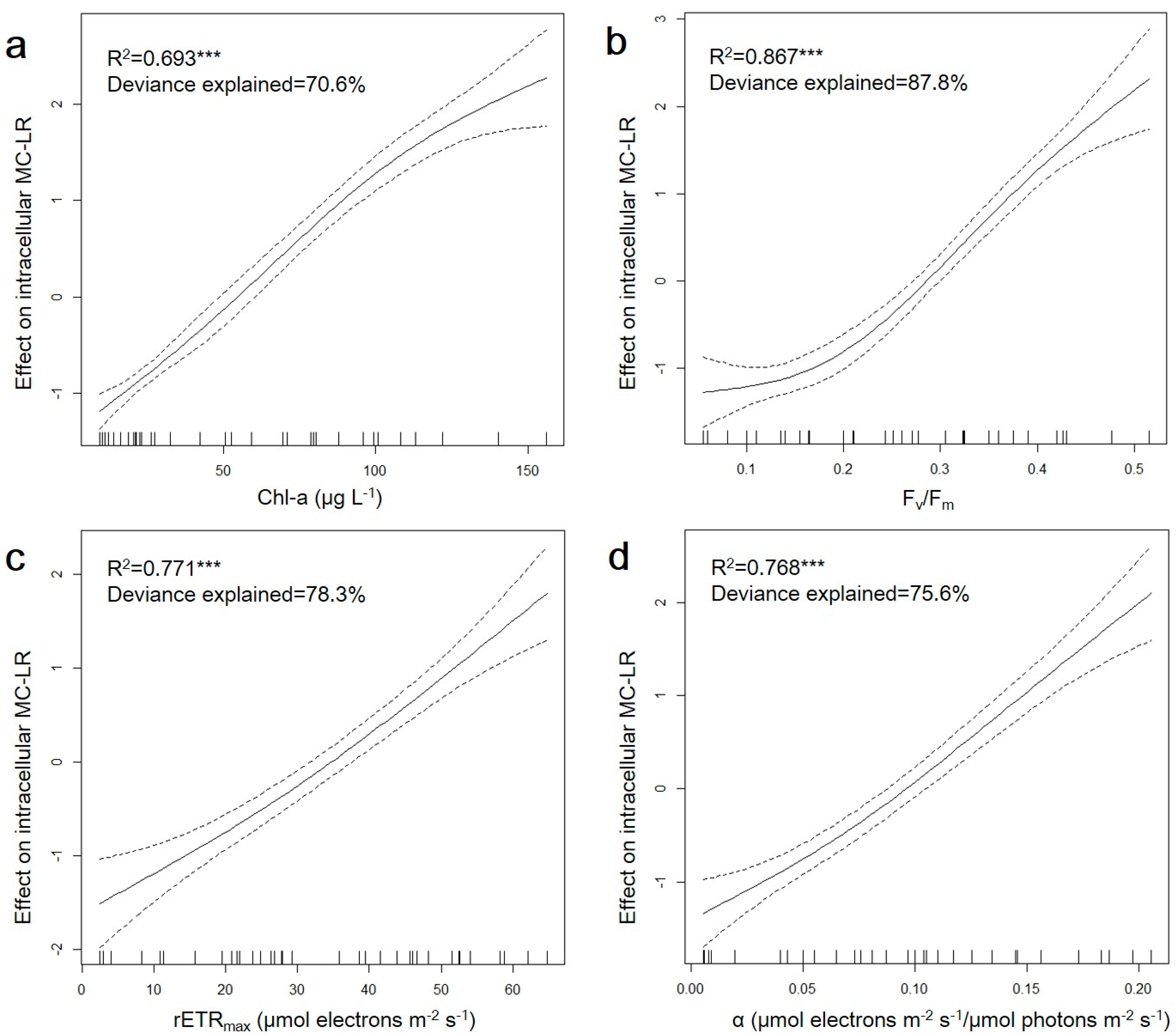
3.3. Relationship between Photosynthetic Capacity and MC–LR Production of M. aeruginosa under Iron Variation
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Waal, D.B.V.D.; Ferreruela, G.; Tonk, L.; Donk, E.V.; Huisman, J.; Visser, P.M.; Matthijs, H.C.P. Pulsed nitrogen supply induces dynamic changes in the amino acid composition and microcystin production of the harmful cyanobacterium planktothrix agardhii. FEMS Microbiol. Ecol. 2010, 74, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Sarnelle, O.; White, J.D.; Horst, G.P.; Hamilton, S.K. Phosphorus addition reverses the positive effect of zebra mussels (dreissena polymorpha) on the toxic cyanobacterium, microcystis aeruginosa. Water Res. 2012, 46, 3471–3478. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Rai, P.K.; Chau, R.; Ravi, A.K.; Neilan, B.A.; Asthana, R.K. Temporal variations in microcystin–producing cells and microcystin concentrations in two fresh water ponds. Water Res. 2015, 69, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Parker, D.L.; Rai, L.C.; Mallick, N.; Rai, P.K.; Kumar, H.D. Effects of cellular metabolism and viability on metal ion accumulation by cultured biomass from a bloom of the cyanobacterium microcystis aeruginosa. Appl. Environ. Microbiol. 1998, 64, 1545–1547. [Google Scholar] [PubMed]
- Schwarz, R.; Forchhammer, K. Acclimation of unicellular cyanobacteria to macronutrient deficiency: Emergence of a complex network of cellular responses. Microbiology 2005, 151, 2503–2514. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W.W. Health effects of toxin–producing cyanobacteria: “The cyanohabs”. Hum. Ecol. Risk Assess. Int. J. 2001, 7, 1393–1407. [Google Scholar] [CrossRef]
- Svirčev, Z.; Drobac, D.; Tokodi, N.; Mijović, B.; Codd, G.A.; Meriluoto, J. Toxicology of microcystins with reference to cases of human intoxications and epidemiological investigations of exposures to cyanobacteria and cyanotoxins. Arch. Toxicol. 2017, 91, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Banack, S.A.; Cox, P.A. Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Wiley: Hoboken, NJ, USA, 2017. [Google Scholar]
- Dixit, F.; Barbeau, B.; Mohseni, M. Characteristics of competitive uptake between microcystin–lr and natural organic matter (nom) fractions using strongly basic anion exchange resins. Water Res. 2018, 139, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Cyanobacterial Toxins: Microcystin–LR in Drinking–Water. Available online: http://www.who.int/water_sanitation_health/dwq/chemicals/cyanobactoxins.pdf (accessed on 5 September 2018).
- Alexandre, C.; Vitor, V. Molecular mechanisms of microcystin toxicity in animal cells. Int. J. Mol. Sci. 2010, 11, 268–287. [Google Scholar]
- Cheng, H.; Yan, W.; Wu, Q.; Lu, J.; Liu, C.; Hung, T.; Li, G. Adverse reproductive performance in zebrafish with increased bioconcentration of microcystin–lr in the presence of titanium dioxide nanoparticles. Environ. Sci. Nano 2018, 5, 1208–1217. [Google Scholar] [CrossRef]
- Amé, M.V.; Wunderlin, D.A. Effects of iron, ammonium and temperature on microcystin content by a natural concentrated microcystis aeruginosa population. Water Air Soil Pollt. 2005, 168, 235–248. [Google Scholar] [CrossRef]
- Dziallas, C.; Grossart, H.P. Increasing oxygen radicals and water temperature select for toxic microcystis sp. PLoS ONE 2011, 6, e25569. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Bu, L.; Shi, Z.; Deng, L.; Zhu, S.; Gao, N. Electrochemical inactivation of microcystis aeruginosa using bdd electrodes: Kinetic modeling of microcystins release and degradation. J. Hazard. Mater. 2017, 346, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Utkilen, H.; Gjølme, N. Iron–stimulated toxin production in microcystis aeruginosa. Appl. Environ. Microbiol. 1995, 61, 797–800. [Google Scholar] [PubMed]
- Alexova, R.; Fujii, M.; Birch, D.; Cheng, J.; Waite, T.D.; Ferrari, B.C.; Neilan, B.A. Iron uptake and toxin synthesis in the bloom–forming microcystis aeruginosa under iron limitation. Environ. Microbiol. 2011, 13, 1064–1077. [Google Scholar] [CrossRef] [PubMed]
- Emma, S.; Beatriz, M.L.; Laura, V.; Teresa, B.M.; Fillat, M.F.; Luisa, P.M. Iron availability affects mcyd expression and microcystin–lr synthesis in microcystis aeruginosa pcc7806. Environ. Microbiol. 2008, 10, 2476–2483. [Google Scholar]
- Alexova, R.; Dang, T.C.; Fujii, M.; Raftery, M.J.; Waite, T.D.; Ferrari, B.C.; Neilan, B.A. Specific global responses to n and fe nutrition in toxic and non-toxic microcystis aeruginosa. Environ. Microbiol. 2016, 18, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Holland, A.; Kinnear, S. Interpreting the possible ecological role (s) of cyanotoxins: Compounds for competitive advantage and/or physiological aide? Mar. Drugs 2013, 11, 2239–2258. [Google Scholar] [CrossRef] [PubMed]
- Zilliges, Y.; Kehr, J.C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Börner, T.; Dittmann, E. The cyanobacterial hepatotoxin microcystin binds to proteins and increases the fitness of microcystis under oxidative stress conditions. PLoS ONE 2012, 6, e17615. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, A.; Harel, M.; Kaplan–Levy, R.N.; Hadas, O.; Sukenik, A.; Dittmann, E. The languages spoken in the water body (or the biological role of cyanobacterial toxins). Front. Microbiol. 2012, 3, 138. [Google Scholar] [CrossRef] [PubMed]
- Lukac, M.; Aegerter, R. Influence of trace metals on growth and toxin production of microcystis aeruginosa. Toxicon 1993, 31, 293–305. [Google Scholar] [CrossRef]
- Fujii, M.; Rose, A.L.; Omura, T.; Waite, T.D. Effect of fe(ii) and fe(iii) transformation kinetics on iron acquisition by a toxic strain of microcystis aeruginosa. Environ. Sci. Technol. 2010, 44, 1980–1986. [Google Scholar] [CrossRef] [PubMed]
- Guidelines for Drinking–Water Quality. Available online: http://www.who.int/water_sanitation_health/dwq/fulltext.pdf (accessed on 5 September 2018).
- North, R.; Guildford, S.; Smith, R.; Havens, S.; Twiss, M. Evidence for phosphorus, nitrogen, and iron colimitation of phytoplankton communities in lake erie. Limnol. Oceanogr. 2007, 52, 315–328. [Google Scholar] [CrossRef]
- Hyenstrand, P.; Rydin, E.; Gunnerhed, M. Response of pelagic cyanobacteria to iron additions—Enclosure experiments from lake erken. J. Plankton Res. 2000, 22, 1113–1126. [Google Scholar] [CrossRef]
- Wang, C.; Kong, H.N.; Wang, X.Z.; Wu, H.D.; Lin, Y.; He, S.B. Effects of iron on growth and intracellular chemical contents of microcystis aeruginosa. Biomed. Environ. Sci. 2010, 23, 48–52. [Google Scholar] [CrossRef]
- Sinang, S.C.; Reichwaldt, E.S.; Ghadouani, A. Local nutrient regimes determine site–specific environmental triggers of cyanobacterial and microcystin variability in urban lakes. Hydrol. Earth Syst. Sci. 2015, 9, 2179–2195. [Google Scholar] [CrossRef]
- Xu, H.; Zhu, G.; Qin, B.; Paerl, H.W. Growth response of microcystis spp. To iron enrichment in different regions of lake taihu, China. Hydrobiologia 2013, 700, 187–202. [Google Scholar] [CrossRef]
- Mckay, R.M.L.; Bullerjahn, G.; Porta, D.; Brown, E.; Sherrell, R.; Smutka, T.; Sterner, R.; Twiss, M.; Wilhelm, S. Consideration of the bioavailability of iron in the north american great lakes: Development of novel approaches toward understanding iron biogeochemistry. Aquat. Ecosyst. Health 2004, 7, 475–490. [Google Scholar] [CrossRef]
- Li, H.; Murphy, T.; Guo, J.; Parr, T.; Nalewajko, C. Iron–stimulated growth and microcystin production of microcystis novacekii uam 250. Limnologica 2009, 39, 255–259. [Google Scholar] [CrossRef]
- Eid, R.; Arab, N.T.; Greenwood, M.T. Iron mediated toxicity and programmed cell death: A review and a re–examination of existing paradigms. BBA-MOL Cell Res. 2017, 1864, 399–430. [Google Scholar] [CrossRef] [PubMed]
- Cottet, M.; Descloux, S.; Guédant, P.; Godon, A.; Cerdan, P.; Vigouroux, R. Total iron concentrations in waters and fish tissues in the nam theun 2 reservoir area (lao PDR). Environ. Monit. Assess. 2015. [Google Scholar] [CrossRef] [PubMed]
- Oppong–Anane, A.B.; Quiñones, K.Y.D.; Harris, W.; Townsend, T.; Bonzongo, J.C.J. Iron reductive dissolution in vadose zone soils: Implication for groundwater pollution in landfill impacted sites. Appl. Geochem. 2018, 94, 21–27. [Google Scholar] [CrossRef]
- Xing, W.; Huang, W.M.; Li, D.H.; Liu, Y.D. Effects of iron on growth, pigment content, photosystem ii efficiency, and siderophores production of microcystis aeruginosa and microcystis wesenbergii. Curr. Microbiol. 2007, 55, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Geider, R.J.; Roche, J.L. The role of iron in phytoplankton photosynthesis, and the potential for iron–limitation of primary productivity in the sea. Photosynth. Res. 1994, 39, 275–301. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.; Gao, N.; Deng, Y.; Qiao, J.; Wang, H. Immediate and long–term impacts of uv–c irradiation on photosynthetic capacity, survival and microcystin–lr release risk of microcystis aeruginosa. Water Res. 2012, 46, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Legendre, L.; Jiao, N. Phytoplankton responses to nitrogen and iron limitation in the tropical and subtropical pacific ocean. J. Plankton Res. 2015, 37, 306–319. [Google Scholar] [CrossRef]
- Jiang, Y.; Ji, B.; Wong, R.N.S.; Wong, M.H. Statistical study on the effects of environmental factors on the growth and microcystins production of bloom–forming cyanobacterium—microcystis aeruginosa. Harmful Algae 2008, 7, 127–136. [Google Scholar] [CrossRef]
- Jähnichen, S.; Long, B.M.; Petzoldt, T. Microcystin production by microcystis aeruginosa: Direct regulation by multiple environmental factors. Harmful Algae 2011, 12, 95–104. [Google Scholar] [CrossRef]
- Jüttner, F.; Lüthi, H. Topology and enhanced toxicity of bound microcystins in microcystis pcc 7806. Toxicon 2008, 51, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.A.; Hisbergues, M.; Börner, T.; Dittmann, E.; Neilan, B.A. Inactivation of an abc transporter gene, mcyh, results in loss of microcystin production in the cyanobacterium microcystis aeruginosa pcc 7806. Appl. Environ. Microbiol. 2004, 70, 6370–6378. [Google Scholar] [CrossRef] [PubMed]
- Neilan, B.A.; Pearson, L.A.; Muenchhoff, J.; Moffitt, M.C.; Dittmann, E. Environmental conditions that influence toxin biosynthesis in cyanobacteria. Environ. Microbiol. 2013, 15, 1239–1253. [Google Scholar] [CrossRef] [PubMed]
- Ralph, P.J.; Gademann, R. Rapid light curves: A powerful tool to assess photosynthetic activity. Aquat. Bot. 2005, 82, 222–237. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.; Wang, P.; Chen, B.; Hou, J.; Jin, Q.; Yang, Y. Effects of iron on growth, antioxidant enzyme activity, bound extracellular polymeric substances and microcystin production of microcystis aeruginosa fachb–905. Ecotoxicol. Environ. Saf. 2016, 132, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.L.; Pires, J.C.M.; Simões, M. The effects of light and temperature on microalgal growth and nutrient removal: An experimental and mathematical approach. RSC Adv. 2016, 6, 22896–22907. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Garrido, M.; Cecchi, P.; Vaquer, A.; Pasqualini, V. Effects of sample conservation on assessments of the photosynthetic efficiency of phytoplankton using pam fluorometry. Deep Sea Res. Pt I 2013, 71, 38–48. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R. Generalized Additive Models; Chapman and Hall/CRC: Boca Raton, FL, USA, 1990; pp. 587–602. [Google Scholar]
- Yi, Y.; Sun, J.; Zhang, S. A habitat suitability model for chinese sturgeon determined using the generalized additive method. J. Hydrol. 2016, 534, 11–18. [Google Scholar] [CrossRef]
- Li, Y.; Ye, T.; Liu, W.; Gao, Y. Linking livestock snow disaster mortality and environmental stressors in the qinghai–tibetan plateau: Quantification based on generalized additive models. Sci. Total Envirn. 2018, 625, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Dang, T.C.; Fujii, M.; Rose, A.L.; Bligh, M.; Waite, T.D. Characteristics of the freshwater cyanobacterium microcystis aeruginosa grown in iron–limited continuous culture. Appl. Environ. Microbiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, S.; Zhang, J.; Gao, B. Growth, microcystin–production and proteomic responses of microcystis aeruginosa under long–term exposure to amoxicillin. Water Res. 2016, 93, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Qiu, Z.; Zhou, Y.; Du, Y.; Liu, C.; Ye, J.; Hu, X. Physiological effects of the herbicide glyphosate on the cyanobacterium microcystis aeruginosa. Aquat. Toxicol. 2016, 178, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Vermaas, W.F. Photosynthesis and Respiration in Cyanobacteria. Available online: https://pdfs.semanticscholar.org/a0f6/26ac1656b606f0cd3aa31318ec68d511c7aa.pdf (accessed on 7 September 2018).
- Zhou, W.; Juneau, P.; Qiu, B. Growth and photosynthetic responses of the bloom–forming cyanobacterium microcystis aeruginosa to elevated levels of cadmium. Chemosphere 2006, 65, 1738. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Shao, Y.; Gao, N.; Deng, Y.; Qiao, J.; Ou, H.; Deng, J. Effects of different algaecides on the photosynthetic capacity, cell integrity and microcystin–lr release of microcystis aeruginosa. Sci. Total Envirn. 2013, s463–464, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Gągała, I.; Izydorczyk, K.; Jurczak, T.; Pawełczyk, J.; Dziadek, J.; Wojtal–Frankiewicz, A.; Jóźwik, A.; Jaskulska, A.; Mankiewicz–Boczek, J. Role of environmental factors and toxic genotypes in the regulation of microcystins–producing cyanobacterial blooms. Microb. Ecol. 2014, 67, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.; Jie, X.; Wang, P.; Li, S.; Wang, G.; Li, Y.; Li, Y.; Acharya, K. Effect of linoleic acid sustained–release microspheres on microcystis aeruginosa antioxidant enzymes activity and microcystins production and release. Chemosphere 2015, 121, 110. [Google Scholar] [CrossRef] [PubMed]
- Spiller, S.C.; Castelfranco, A.M.; Castelfranco, P.A. Effects of iron and oxygen on chlorophyll biosynthesis i. In vivo observations on iron and oxygen–deficient plants. Plant Physiol. 1982, 69, 112–116. [Google Scholar] [CrossRef]
- Reinbothe, C.; Bakkouri, M.E.; Buhr, F.; Muraki, N.; Nomata, J.; Kurisu, G.; Fujita, Y.; Reinbothe, S. Chlorophyll biosynthesis: Spotlight on protochlorophyllide reduction. Trends Plant Sci. 2010, 15, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Chi, G.; Huang, B.; Ma, J.; Shi, Y.; Chen, X. Effects of iron on growth and reflectance spectrum of the bloom-forming cyanobacterium microcystis viridis. Phycol. Res. 2015, 63, 265–273. [Google Scholar] [CrossRef]
- Macintyre, H.L.; Kana, T.M.; Anning, T.; Geider, R.J. Photoacclimation of photosynthesis irradiance response curves and photosynthetic pigments in microalgae and cyanobacteria. J. Phycol. 2002, 38, 17–38. [Google Scholar] [CrossRef]
- Nogales, J.; Gudmundsson, S.; Knight, E.M.; Palsson, B.O.; Thiele, I. Detailing the optimality of photosynthesis in cyanobacteria through systems biology analysis. Bioengineered 2012, 109, 2678–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Bba-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Behrenfeld, M.J.; Prasil, O.; Babin, M.; Bruyant, F. In search of a physiological basis for covariations in light–limited and light–saturated photosynthesis 1. J. Phycol. 2004, 40, 4–25. [Google Scholar] [CrossRef]
- Greene, R.M.; Geider, R.J.; Kolber, Z.; Falkowski, P.G. Iron–induced changes in light harvesting and photochemical energy conversion processes in eukaryotic marine algae. Plant Physiol. 1992, 100, 565. [Google Scholar] [CrossRef] [PubMed]
- Fujii, M.; Dang, T.C.; Rose, A.L.; Omura, T.; Waite, T.D. Effect of light on iron uptake by the freshwater cyanobacterium microcystis aeruginosa. Environ. Sci. Technol. 2011, 45, 1391. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Li, J.; Pan, X.; Sun, Z.; Ye, C.; Jin, G.; Fu, Z. Effects of streptomycin on growth of algae chlorella vulgaris and microcystis aeruginosa. Environ. Toxicol. 2012, 27, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Polyak, Y.; Zaytseva, T.; Medvedeva, N. Response of toxic cyanobacterium microcystis aeruginosa to environmental pollution. Water Air Soil Pollt. 2013, 224, 1494. [Google Scholar] [CrossRef]
- Waal, D.B.V.D.; Verspagen, J.M.H.; Lürling, M.; Donk, E.V.; Visser, P.M.; Huisman, J. The ecological stoichiometry of toxins produced by harmful cyanobacteria: An experimental test of the carbon-nutrient balance hypothesis. Ecol. Lett. 2009, 12, 1326–1335. [Google Scholar] [CrossRef] [PubMed]
- Yeung, A.C.Y.; D’Agostino, P.M.; Poljak, A.; Mcdonald, J.; Bligh, M.W.; Waite, T.D.; Neilan, B.A. Physiological and proteomic responses of continuous cultures of microcystis aeruginosa pcc 7806 to changes in iron bioavailability and growth rate. Appl. Environ. Microbiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Wiedner, C.; Visser, P.M.; Fastner, J.; Metcalf, J.S.; Codd, G.A.; Mur, L.R. Effects of light on the microcystin content of microcystis strain pcc 7806. Appl. Environ. Microbiol. 2003. [Google Scholar] [CrossRef]
- Gerbersdorf, S.U. An advanced technique for immuno–labelling of microcystins in cryosectioned cells of microcystis aeruginosa pcc 7806 (cyanobacteria): Implementations of an experiment with varying light scenarios and culture densities. Toxicon 2006, 47, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Latifi, A.; Ruiz, M.; Zhang, C.C. Oxidative stress in cyanobacteria. Fems Microbiol. Rev. 2009, 33, 258–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deblois, C.P.; Juneau, P. Relationship between photosynthetic processes and microcystin in microcystis aeruginosa grown under different photon irradiance. Harmful Algae 2010, 9, 18–24. [Google Scholar] [CrossRef]
- Gouvea, S.P.; Twiss, B.M.R. Influence of ultraviolet radiation, copper, and zinc on microcystin content in microcystis aeruginosa (cyanobacteria). Harmful Algae 2008, 7, 194–205. [Google Scholar] [CrossRef]
- Briand, E.; Bormans, M.; Quiblier, C.; Salençon, M.J.; Humbert, J.F. Evidence of the cost of the production of microcystins by microcystis aeruginosa under differing light and nitrate environmental conditions. PLoS ONE 2012, 7, e29981. [Google Scholar] [CrossRef]
- Hesse, K.; Dittmann, E.; Börner, T. Consequences of impaired microcystin production for light–dependent growth and pigmentation of microcystis aeruginosa pcc 7806. Fems Microbiol. Ecol. 2001, 37, 39–43. [Google Scholar] [CrossRef]
- Beecraft, L.; Watson, S.B.; Smith, R.E.H. Multi-wavelength pulse amplitude modulated fluorometry (phyto-pam) reveals differential effects of ultraviolet radiation on the photosynthetic physiology of phytoplankton pigment groups. Freshw. Biol. 2017. [Google Scholar] [CrossRef]
- Wang, Z.; Li, D.; Li, G.; Liu, Y. Mechanism of photosynthetic response in microcystis aeruginosa pcc7806 to low inorganic phosphorus. Harmful Algae 2010, 9, 613–619. [Google Scholar] [CrossRef]
- Meissner, S.; Fastner, J.; Dittmann, E. Microcystin production revisited: Conjugate formation makes a major contribution. Environ. Microbiol. 2013, 15, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Ceballos–Laita, L.; Calvo–Begueria, L.; Lahoz, J.; Bes, M.-T.; Fillat, M.F.; Peleato, M.-L. γ–lindane increases microcystin synthesis in microcystis aeruginosa pcc7806. Mar. Drugs 2015, 13, 5666–5680. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, E.; Martinluna, B.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. An active photosynthetic electron transfer chain required for mcyd transcription and microcystin synthesis in microcystis aeruginosa pcc7806. Ecotoxicology 2012, 21, 811–819. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Wang, P.; Wang, C.; Qian, J.; Feng, T.; Yang, Y. Relationship between Photosynthetic Capacity and Microcystin Production in Toxic Microcystis Aeruginosa under Different Iron Regimes. Int. J. Environ. Res. Public Health 2018, 15, 1954. https://doi.org/10.3390/ijerph15091954
Wang X, Wang P, Wang C, Qian J, Feng T, Yang Y. Relationship between Photosynthetic Capacity and Microcystin Production in Toxic Microcystis Aeruginosa under Different Iron Regimes. International Journal of Environmental Research and Public Health. 2018; 15(9):1954. https://doi.org/10.3390/ijerph15091954
Chicago/Turabian StyleWang, Xun, Peifang Wang, Chao Wang, Jin Qian, Tao Feng, and Yangyang Yang. 2018. "Relationship between Photosynthetic Capacity and Microcystin Production in Toxic Microcystis Aeruginosa under Different Iron Regimes" International Journal of Environmental Research and Public Health 15, no. 9: 1954. https://doi.org/10.3390/ijerph15091954
APA StyleWang, X., Wang, P., Wang, C., Qian, J., Feng, T., & Yang, Y. (2018). Relationship between Photosynthetic Capacity and Microcystin Production in Toxic Microcystis Aeruginosa under Different Iron Regimes. International Journal of Environmental Research and Public Health, 15(9), 1954. https://doi.org/10.3390/ijerph15091954