Effects of Sub-Acute Manganese Exposure on Thyroid Hormone and Glutamine (Gln)/Glutamate (Glu)-γ- Aminobutyric Acid (GABA) Cycle in Serum of Rats

Abstract
:1. Introduction
2. Methods and Materials
2.1. Experimental Animals
2.2. Experimental Design
2.3. Determination of Mn Levels in Serum
2.4. Measurement of Triiodothyronine (T3) and Thyroxin (T4) Levels in Serum
2.5. Determination of Amino Acids Neurotransmitters Levels in Serum
2.6. Statistical Analysis
3. Results
3.1. Mn Levels in Serum of Mn-exposed Rats Were Increased
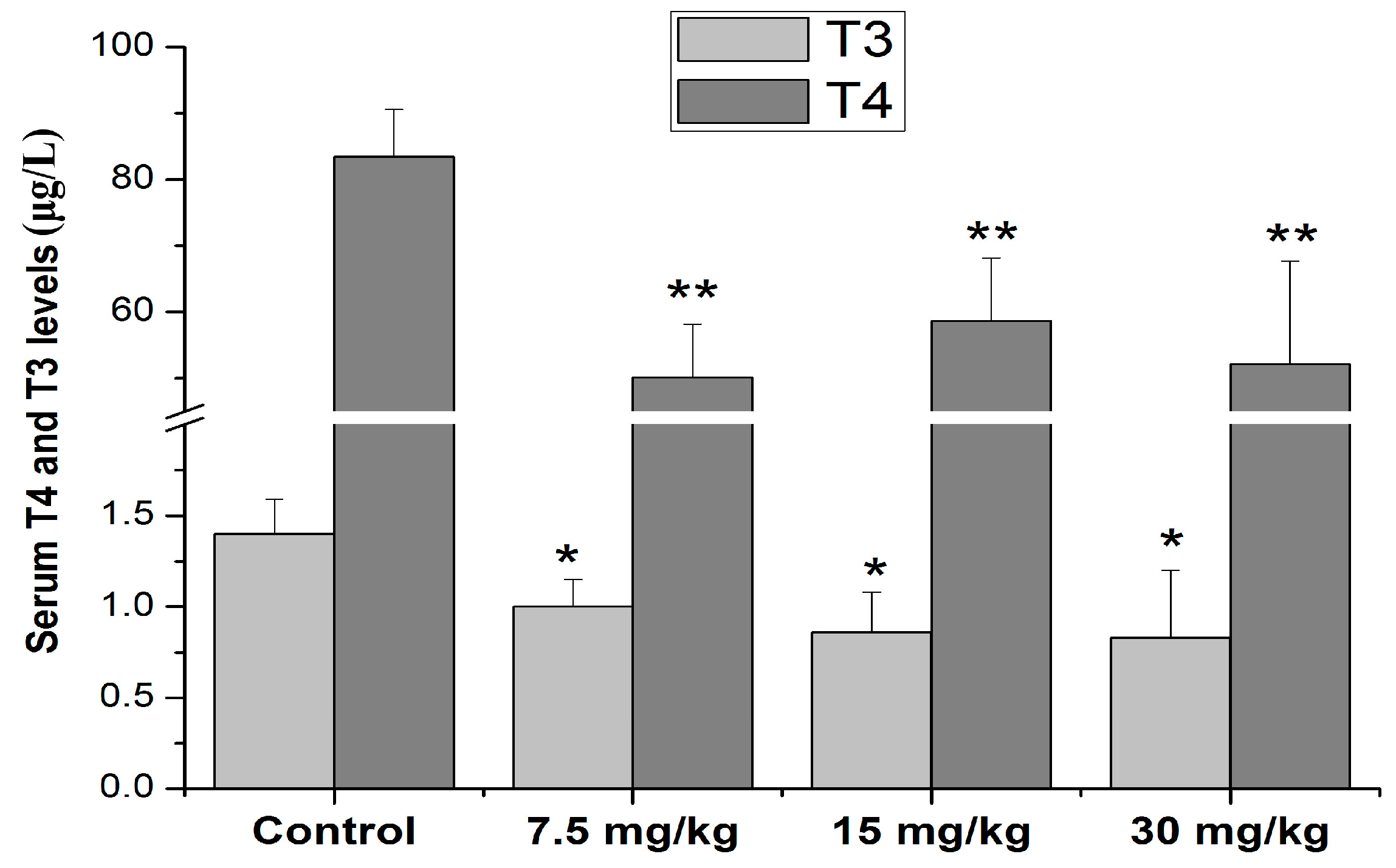
3.2. Effects of Mn Treatment on T4 and T3 Levels in Serum of Rats
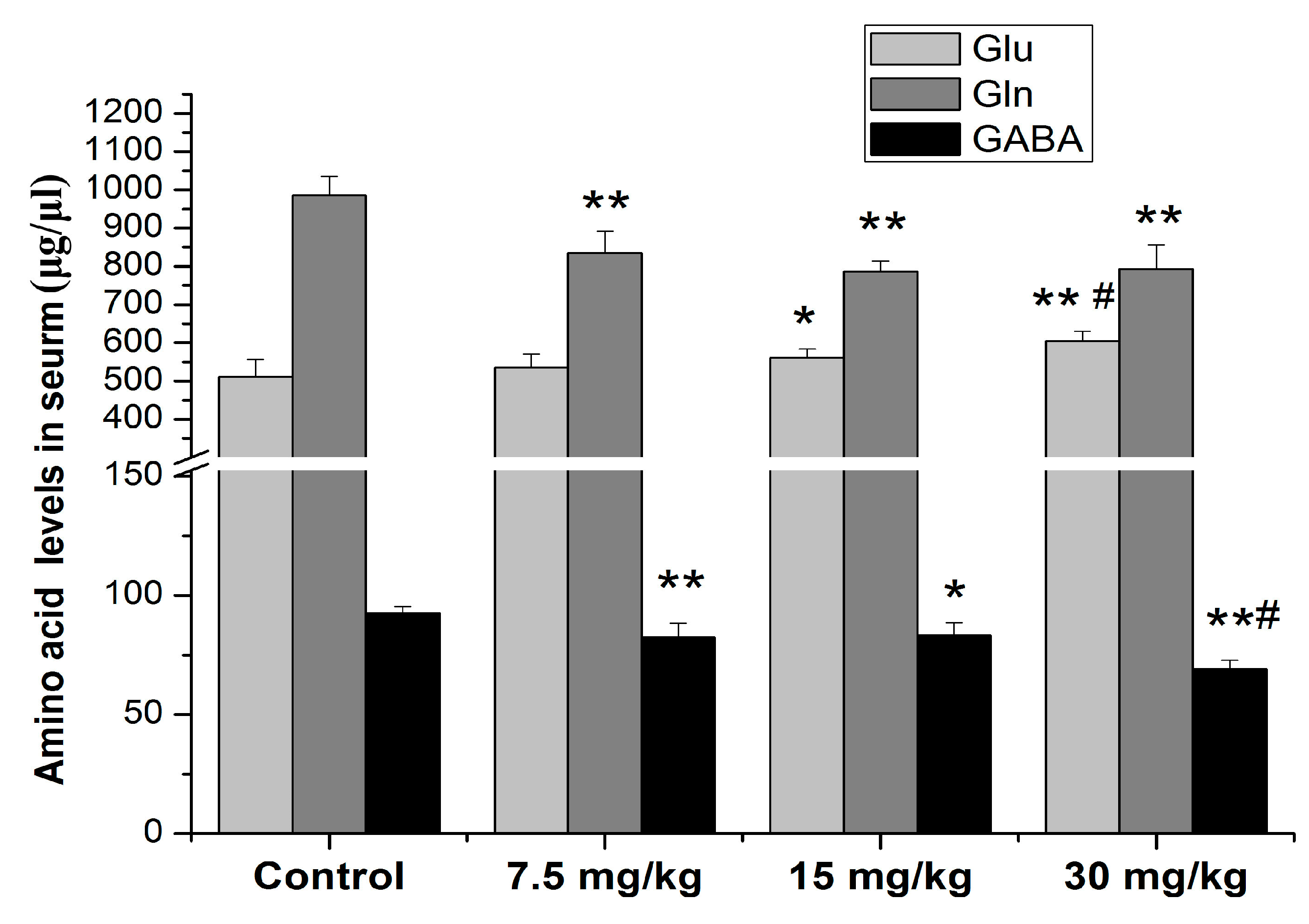
3.3. Effects of Mn Treatment on Serum Amino Acid Neurotransmitter Levels of Rats
3.4. Partial Correlation between Serum Mn Levels and Other Indicators
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Aschner, J.L.; Aschner, M. Nutritional aspects of manganese homeostasis. Mol. Aspects Med. 2005, 26, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Brazhnik, E.; Shah, F.; Tepper, J.M. GABAergic afferents activate both GABAA and GABAB receptors in mouse substantia nigra dopaminergic neurons in vivo. J. Neurosci. 2008, 28, 10386–10398. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Finley, E.J.; Gavin, C.E.; Aschner, M.; Gunter, T.E. Manganese neurotoxicity and the role of reactive oxygen species. Free Radic. Biol. Med. 2013, 62, 65–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pajarillo, E.; Johnson, J., Jr.; Kim, J.; Karki, P.; Son, D.S.; Aschner, M.; Lee, E. 17beta-estradiol and tamoxifen protect mice from manganese-induced dopaminergic neurotoxicity. Neurotoxicology 2018, 65, 280–288. [Google Scholar] [CrossRef]
- Chen, H.; Copes, R. Manganese in drinking water and intellectual impairment in school-age children. Environ. Health Perspect. 2011, 119, A240–A241. [Google Scholar] [CrossRef] [PubMed]
- Criswell, S.R.; Warden, M.N.; Nielsen, S.S.; Perlmutter, J.S.; Moerlein, S.M.; Sheppard, L.; Lenox-Krug, J.; Checkoway, H.; Racette, B.A. Selective D2 receptor PET in manganese-exposed workers. Neurology 2018, 91, e1022–e1030. [Google Scholar] [CrossRef] [PubMed]
- Lakhan, S.E.; Abboud, H. Teaching neuroimages: Manganese neurotoxicity of the basal ganglia and thalamus. Neurology 2013, 81, e111. [Google Scholar] [CrossRef]
- Konig, S.; Neto, V.M. Thyroid hormone actions on neural cells. Cell Mol. Neurobiol. 2002, 22, 517–544. [Google Scholar] [CrossRef]
- Edwards, N.J.; Tejeda, H.A.; Pignatelli, M.; Zhang, S.; McDevitt, R.A.; Wu, J.; Bass, C.E.; Bettler, B.; Morales, M.; Bonci, A. Circuit specificity in the inhibitory architecture of the VTA regulates cocaine-induced behavior. Nat. Neurosci. 2017, 20, 438–448. [Google Scholar] [CrossRef]
- Chen, A.; Kim, S.S.; Chung, E.; Dietrich, K.N. Thyroid hormones in relation to lead, mercury, and cadmium exposure in the National Health and Nutrition Examination Survey, 2007–2008. Environ. Health Perspect. 2013, 121, 181–186. [Google Scholar] [CrossRef]
- Barysheva, E.S. Experimental Simulation of the Effects of Essential and Toxic Trace Elements on Thyroid Function. Bull. Exp. Biol. Med. 2018, 164, 439–441. [Google Scholar] [CrossRef]
- Hanif, S.; Ilyas, A.; Shah, M.H. Statistical Evaluation of Trace Metals, TSH and T4 in Blood Serum of Thyroid Disease Patients in Comparison with Controls. Biol. Trace Elem. Res. 2018, 183, 58–70. [Google Scholar] [CrossRef]
- Struzynska, L.; Sulkowski, G. Relationships between glutamine, glutamate, and GABA in nerve endings under Pb-toxicity conditions. J. Inorg. Biochem. 2004, 98, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.F.; Zhang, L.M.; Zhang, Z.; Liu, N.; Xu, S.W.; Lin, H.J. Manganese-induced effects on testicular trace element levels and crucial hormonal parameters of Hyline cocks. Biol. Trace Elem. Res. 2013, 151, 217–224. [Google Scholar] [CrossRef]
- Ou, S.Y.; Luo, H.L.; Mailman, R.B.; Li, Z.C.; Zhang, Y.W.; Cai, M.; Huang, X.W.; Li, S.J.; Jiang, Y.M. Effect of manganese on neural endocrine hormones in serum of welders and smelters. J. Trace Elem. Med. Biol. 2018, 50, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bowler, R.M.; Beseler, C.L.; Gocheva, V.V.; Gocheva, M. Environmental exposure to manganese in air: Associations with tremor and motor function. Sci. Total Environ. 2016, 541, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xing, R.; Liu, S.; Yu, H.; Li, P. gamma-Aminobutyric acid ameliorates fluoride-induced hypothyroidism in male Kunming mice. Life Sci. 2016, 146, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wiens, S.C.; Trudeau, V.L. Thyroid hormone and gamma-aminobutyric acid (GABA) interactions in neuroendocrine systems. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2006, 144, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.E.; Sui, L.; Walker, M.J.; Anderson, W.; Thomas, S.; Smoller, S.N.; Schon, J.P.; Phani, S.; Goodman, J.H. Thyroid hormone insufficiency during brain development reduces parvalbumin immunoreactivity and inhibitory function in the hippocampus. Endocrinology 2007, 148, 92–102. [Google Scholar] [CrossRef]
- Anderson, J.G.; Cooney, P.T.; Erikson, K.M. Brain manganese accumulation is inversely related to gamma-amino butyric acid uptake in male and female rats. Toxicol. Sci. 2007, 95, 188–195. [Google Scholar] [CrossRef]
- Kwakye, G.F.; Paoliello, M.M.; Mukhopadhyay, S.; Bowman, A.B.; Aschner, M. Manganese-Induced Parkinsonism and Parkinson’s Disease: Shared and Distinguishable Features. Int. J. Environ. Res. Public Health 2015, 12, 7519–7540. [Google Scholar] [CrossRef] [PubMed]
- Li, S.J.; Ou, C.Y.; He, S.N.; Huang, X.W.; Luo, H.L.; Meng, H.Y.; Lu, G.D.; Jiang, Y.M.; Peres, T.V.; Luo, Y.N.; et al. Sodium p-Aminosalicylic Acid Reverses Sub-Chronic Manganese-Induced Impairments of Spatial Learning and Memory Abilities in Rats, but Fails to Restore gamma-Aminobutyric Acid Levels. Int. J. Environ. Res. Public Health 2017, 14, 400. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, Q.; Tong, S.-Y.; Ding, L.; Liu, N.; Gao, D.-S. Changes of gamma-aminobutyric acid in peripheral blood of patients with mild cognitive impairment in Parkinson’s disease. Shandong Pharm. 2018, 58, 75–78. [Google Scholar]
- Zeng, Z.-X.; Liu, X.-M.; Liang, X.; Chen, B.-X.; Liu, X.-H.; Wen, B.-Y.; Xu, Z.-M.; Fang, X.-L.; Zhang, Y.-L. Effects of Kudiezi injection on serum neurotransmitters in patients with acute cerebral infarction. J. Peking Univ. Trad. Chin. Med. 2018, 41, 523–528. [Google Scholar]
- Liu, J.-L.; Zhu, P.-X.; Han, X.-J.; Fu, H.; Zhao, Y.; Liu, X. Treatment of acupuncture method of draining yin and nourishing yang combined with Huatan Tongluo decoction in spastic paralysis after stroke. Jilin J. Chin. Med. 2017, 37, 1275–1279. [Google Scholar]
- Ou, C.Y.; Luo, Y.N.; He, S.N.; Deng, X.F.; Luo, H.L.; Yuan, Z.X.; Meng, H.Y.; Mo, Y.H.; Li, S.J.; Jiang, Y.M. Sodium P-Aminosalicylic Acid Improved Manganese-Induced Learning and Memory Dysfunction via Restoring the Ultrastructural Alterations and gamma-Aminobutyric Acid Metabolism Imbalance in the Basal Ganglia. Biol. Trace Elem. Res. 2017, 176, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, X.; Yang, D.; Zhang, H.; Zhao, P.; Fu, J.; Yao, B.; Zhou, Z. ER stress and ER stress-mediated apoptosis are involved in manganese-induced neurotoxicity in the rat striatum in vivo. Neurotoxicology 2015, 48, 109–119. [Google Scholar] [CrossRef]
- Lazrishvili, I.L.; Shukakidze, A.A.; Chkhartishvili, N.N.; Bikashvili, T.Z. Morphological changes and manganese content in the brains of rat pups subjected to subchronic poisoning with manganese chloride. Neurosci. Behav. Physiol. 2009, 39, 7–12. [Google Scholar] [CrossRef]
- Li, S.J.; Qin, W.X.; Peng, D.J.; Yuan, Z.X.; He, S.N.; Luo, Y.N.; Aschner, M.; Jiang, Y.M.; Liang, D.Y.; Xie, B.Y.; et al. Sodium P-aminosalicylic acid inhibits sub-chronic manganese-induced neuroinflammation in rats by modulating MAPK and COX-2. Neurotoxicology 2018, 64, 219–229. [Google Scholar] [CrossRef]
- Li, S.J.; Li, Y.; Chen, J.W.; Yuan, Z.X.; Mo, Y.H.; Lu, G.D.; Jiang, Y.M.; Ou, C.Y.; Wang, F.; Huang, X.W.; et al. Sodium Para-aminosalicylic Acid Protected Primary Cultured Basal Ganglia Neurons of Rat from Manganese-Induced Oxidative Impairment and Changes of Amino Acid Neurotransmitters. Biol. Trace Elem. Res. 2015, 170, 357–365. [Google Scholar] [CrossRef]
- Cigan, S.S.; McGovern, P.M.; Choudhary, K.; Miller, N.C.; Georgieff, M.; Rao, R.; Stepanov, I. Toenail manganese as a potential biomarker for in utero and early childhood exposure studies. Sci. Rep. 2018, 8, 17034. [Google Scholar] [CrossRef] [PubMed]
- Butler, L.; Gennings, C.; Peli, M.; Borgese, L.; Placidi, D.; Zimmerman, N.; Hsu, H.L.; Coull, B.A.; Wright, R.O.; Smith, D.R.; et al. Assessing the contributions of metals in environmental media to exposure biomarkers in a region of ferroalloy industry. J. Expo. Sci. Environ. Epidemiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.L.G.; Araujo, C.F.S.; Santos, N.R.D.; Bandeira, M.J.; Anjos, A.L.S.; Carvalho, C.F.; Lima, C.S.; Abreu, J.N.S.; Mergler, D.; Menezes-Filho, J.A. Airborne manganese exposure and neurobehavior in school-aged children living near a ferro-manganese alloy plant. Environ. Res. 2018, 167, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Hassani, H.; Golbabaei, F.; Shirkhanloo, H.; Tehrani-Doust, M. Relations of biomarkers of manganese exposure and neuropsychological effects among welders and ferroalloy smelters. Ind. Health 2016, 54, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.; Kim, H.; Zhao, Q. Comparative toxicokinetics of manganese chloride and methylcyclopentadienyl manganese tricarbonyl (MMT) in Sprague-Dawley rats. Toxicol. Sci. 2000, 54, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Newland, M.C.; Cox, C.; Hamada, R.; Oberdorster, G.; Weiss, B. The clearance of manganese chloride in the primate. Fundam. Appl. Toxicol. 1987, 9, 314–328. [Google Scholar] [CrossRef]
- Apostoli, P.; Lucchini, R.; Alessio, L. Are current biomarkers suitable for the assessment of manganese exposure in individual workers? Am. J. Ind. Med. 2000, 37, 283–290. [Google Scholar] [CrossRef]
- Greger, J.L.; Davis, C.D.; Suttie, J.W.; Lyle, B.J. Intake, serum concentrations, and urinary excretion of manganese by adult males. Am. J. Clin. Nutr. 1990, 51, 457–461. [Google Scholar] [CrossRef]
- Williams, M.; Todd, G.D.; Roney, N.; Crawford, J.; Coles, C.; McClure, P.R.; Garey, J.D.; Zaccaria, K.; Citra, M. Toxicological Profile for Manganese; ATSDR: Atlanta, GA, USA, 2012.
- Mirmohammadi, S.; Moghaddasi, Y.; Yazdani, J.; Yousefinejad, R.; Esfandyari, Y.; Gorgani, M.; Habibpour, M. Correlation between airborne manganese concentration at the workstations in the iron foundry and manganese concentration in workers’ blood. Med. Pr. 2017, 68, 449–458. [Google Scholar] [CrossRef]
- Huang, C.C.; Lu, C.S.; Chu, N.S.; Hochberg, F.; Lilienfeld, D.; Olanow, W.; Calne, D.B. Progression after chronic manganese exposure. Neurology 1993, 43, 1479–1483. [Google Scholar] [CrossRef]
- Chan, S.; Kilby, M.D. Thyroid hormone and central nervous system development. J. Endocrinol. 2000, 165, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, M.E.; Mack, C.M.; Lasley, S.M. Chronic developmental lead exposure and hippocampal long-term potentiation: Biphasic dose-response relationship. Neurotoxicology 1999, 20, 71–82. [Google Scholar] [PubMed]
- Wu, C.Y.; Liu, B.; Wang, H.L.; Ruan, D.Y. Levothyroxine rescues the lead-induced hypothyroidism and impairment of long-term potentiation in hippocampal CA1 region of the developmental rats. Toxicol. Appl. Pharmacol. 2011, 256, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Sui, L.; Anderson, W.L.; Gilbert, M.E. Impairment in short-term but enhanced long-term synaptic potentiation and ERK activation in adult hippocampal area CA1 following developmental thyroid hormone insufficiency. Toxicol. Sci. 2005, 85, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Zoeller, R.T.; Rovet, J. Timing of thyroid hormone action in the developing brain: Clinical observations and experimental findings. J. Neuroendocrinol. 2004, 16, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Stoker, T.E.; Laws, S.C.; Guidici, D.L.; Cooper, R.L. The effect of atrazine on puberty in male wistar rats: An evaluation in the protocol for the assessment of pubertal development and thyroid function. Toxicol. Sci. 2000, 58, 50–59. [Google Scholar] [CrossRef]
- Guo, J.; Lv, N.; Tang, J.; Zhang, X.; Peng, L.; Du, X.; Li, S.; Luo, Q.; Zhang, D.; Chen, G. Associations of blood metal exposure with thyroid hormones in Chinese pregnant women: A cross-sectional study. Environ. Int. 2018, 121 Pt 2, 1185–1192. [Google Scholar] [CrossRef]
- Kaellis, E. Effect of manganous ions on thyroidal iodine metabolism in the rat. Proc. Soc. Exp. Biol. Med. 1970, 135, 216–218. [Google Scholar] [CrossRef] [PubMed]
- Buthieau, A.M.; Autissier, N. The effect of Mn2+ on thyroid iodine metabolism in rats. C. R. Seances Soc. Biol. Fil. 1977, 171, 1024–1028. [Google Scholar]
- Hoseini, S.M.; Hedayati, A.; Ghelichpour, M. Plasma metabolites, ions and thyroid hormones levels, and hepatic enzymes activity in Caspian roach (Rutilus rutilus caspicus) exposed to waterborne manganese. Ecotoxicol. Environ. Saf. 2014, 107, 84–89. [Google Scholar] [CrossRef]
- Modaberi, S.; Heysieattalab, S.; Shahbazi, M.; Naghdi, N. Combination Effects of Forced Mild Exercise and GABAB Receptor Agonist on Spatial Learning, Memory, and Motor Activity in Striatum Lesion Rats. J. Mot. Behav. 2018, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kolasinski, J.; Hinson, E.L.; Divanbeighi Zand, A.P.; Rizov, A.; Emir, U.E.; Stagg, C.J. The dynamics of cortical GABA in human motor learning. J. Physiol. 2019, 597, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.T.; Chang, Y.G.; Chern, Y. Insights into GABAAergic system alteration in Huntington’s disease. Open Biol. 2018, 8, 180165. [Google Scholar] [CrossRef] [PubMed]
- Giovannetti, E.A.; Fuhrmann, M. Unsupervised excitation: GABAergic dysfunctions in Alzheimer’s disease. Brain Res. 2018, 1707, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Mehan, S.; Singh, S. Understanding multifactorial architecture of Parkinson’s disease: Pathophysiology to management. Neurol. Sci. 2019, 40, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Diana, A.; Pillai, R.; Bongioanni, P.; O’Keeffe, A.G.; Miller, R.G.; Moore, D.H. Gamma aminobutyric acid (GABA) modulators for amyotrophic lateral sclerosis/motor neuron disease. Cochrane Database Syst. Rev. 2017, 1, CD006049. [Google Scholar] [CrossRef] [PubMed]
- Lipe, G.W.; Duhart, H.; Newport, G.D.; Slikker, W., Jr.; Ali, S.F. Effect of manganese on the concentration of amino acids in different regions of the rat brain. J. Environ. Sci. Health B 1999, 34, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.E.; Ward, E.J.; Yeh, C.L.; Snyder, S.; Long, Z.; Gokalp Yavuz, F.; Zauber, S.E.; Dydak, U. Thalamic GABA levels and occupational manganese neurotoxicity: Association with exposure levels and brain MRI. Neurotoxicology 2018, 64, 30–42. [Google Scholar] [CrossRef]
- Bonilla, E.; Arrieta, A.; Castro, F.; Dávila, J.O.; Quiroz, I. Manganese toxicity: Free amino acids in the striatum and olfactory bulb of the mouse. Investig. Clin. 1994, 35, 175–181. [Google Scholar]
- Santos, D.; Batoreu, M.C.; Almeida, I.; Ramos, R.; Sidoryk-Wegrzynowicz, M.; Aschner, M.; Marreilha dos Santos, A.P. Manganese alters rat brain amino acids levels. Biol. Trace Elem. Res. 2012, 150, 337–341. [Google Scholar] [CrossRef]
- Soldin, O.P.; Aschner, M. Effects of manganese on thyroid hormone homeostasis: Potential links. Neurotoxicology 2007, 28, 951–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzano, J.; Cuadrado, M.; Morte, B.; Bernal, J. Influence of thyroid hormone and thyroid hormone receptors in the generation of cerebellar gamma-aminobutyric acid-ergic interneurons from precursor cells. Endocrinology 2007, 148, 5746–5751. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Honda, S.; Okada, A.; Ikeda, A.; Kinoshita, S.; Tomooka, Y. A bipotent neural progenitor cell line cloned from a cerebellum of an adult p53-deficient mouse generates both neurons and oligodendrocytes. Eur. J. Neurosci. 2005, 21, 2903–2911. [Google Scholar] [CrossRef]
- Yi, J.; Zheng, J.Y.; Zhang, W.; Wang, S.; Yang, Z.F.; Dou, K.F. Decreased pain threshold and enhanced synaptic transmission in the anterior cingulate cortex of experimental hypothyroidism mice. Mol. Pain 2014, 10, 38. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Indicator | Mn | Glu | Gln | GABA | T3 |
|---|---|---|---|---|---|
| Mn | - | - | - | - | - |
| Glu | 0.756 ** | - | −0.484 ** | −0.601 ** | - |
| Gln | −0.679 ** | - | - | 0.652 ** | - |
| GABA | −0.782 ** | - | - | - | - |
| T3 | −0.618 ** | 0.225 | −0.120 | 0.423 * | - |
| T4 | −0.719 ** | 0.393 | 0.490 * | 0.274 | −0.322 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ou, C.-Y.; He, Y.-H.; Sun, Y.; Yang, L.; Shi, W.-X.; Li, S.-J. Effects of Sub-Acute Manganese Exposure on Thyroid Hormone and Glutamine (Gln)/Glutamate (Glu)-γ- Aminobutyric Acid (GABA) Cycle in Serum of Rats. Int. J. Environ. Res. Public Health 2019, 16, 2157. https://doi.org/10.3390/ijerph16122157
Ou C-Y, He Y-H, Sun Y, Yang L, Shi W-X, Li S-J. Effects of Sub-Acute Manganese Exposure on Thyroid Hormone and Glutamine (Gln)/Glutamate (Glu)-γ- Aminobutyric Acid (GABA) Cycle in Serum of Rats. International Journal of Environmental Research and Public Health. 2019; 16(12):2157. https://doi.org/10.3390/ijerph16122157
Chicago/Turabian StyleOu, Chao-Yan, Yong-Hua He, Yi Sun, Lin Yang, Wen-Xiang Shi, and Shao-Jun Li. 2019. "Effects of Sub-Acute Manganese Exposure on Thyroid Hormone and Glutamine (Gln)/Glutamate (Glu)-γ- Aminobutyric Acid (GABA) Cycle in Serum of Rats" International Journal of Environmental Research and Public Health 16, no. 12: 2157. https://doi.org/10.3390/ijerph16122157
APA StyleOu, C. -Y., He, Y. -H., Sun, Y., Yang, L., Shi, W. -X., & Li, S. -J. (2019). Effects of Sub-Acute Manganese Exposure on Thyroid Hormone and Glutamine (Gln)/Glutamate (Glu)-γ- Aminobutyric Acid (GABA) Cycle in Serum of Rats. International Journal of Environmental Research and Public Health, 16(12), 2157. https://doi.org/10.3390/ijerph16122157