Genetic Characterization of Antimicrobial-Resistant Escherichia coli Isolated from a Mixed-Use Watershed in Northeast Georgia, USA



Abstract
:1. Introduction
2. Materials and Methods
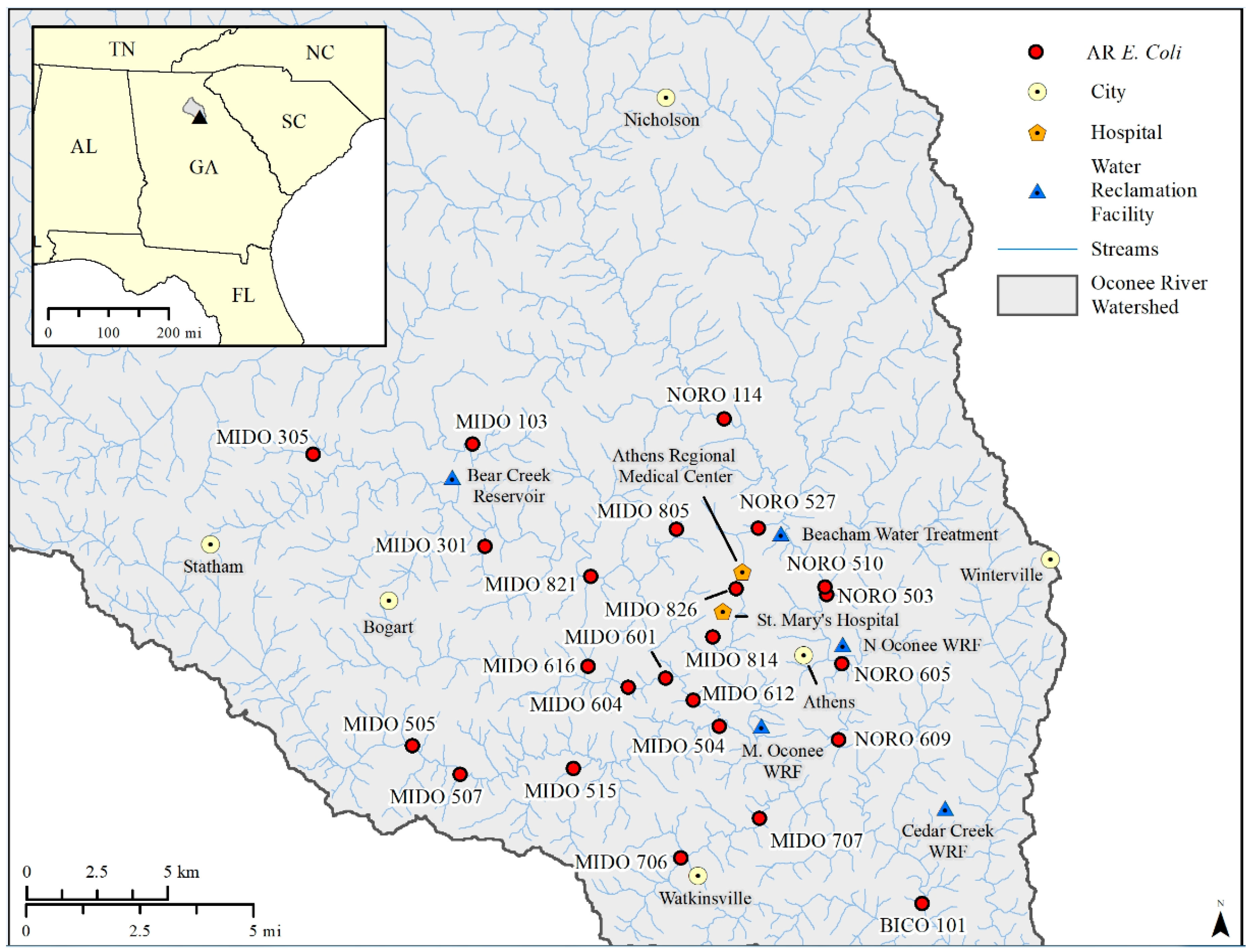
2.1. E. coli Isolates
2.2. PCR Detection of AR Genes and Sequence Analysis
2.3. Phenotypic and Genotypic Detection of ESBL
2.4. Pulsed-Field Gel Electrophoresis (PFGE)
2.5. Multilocus Sequence Typing (MLST)
2.6. Replicon Typing and Integron Analysis
3. Results
3.1. AR Genes
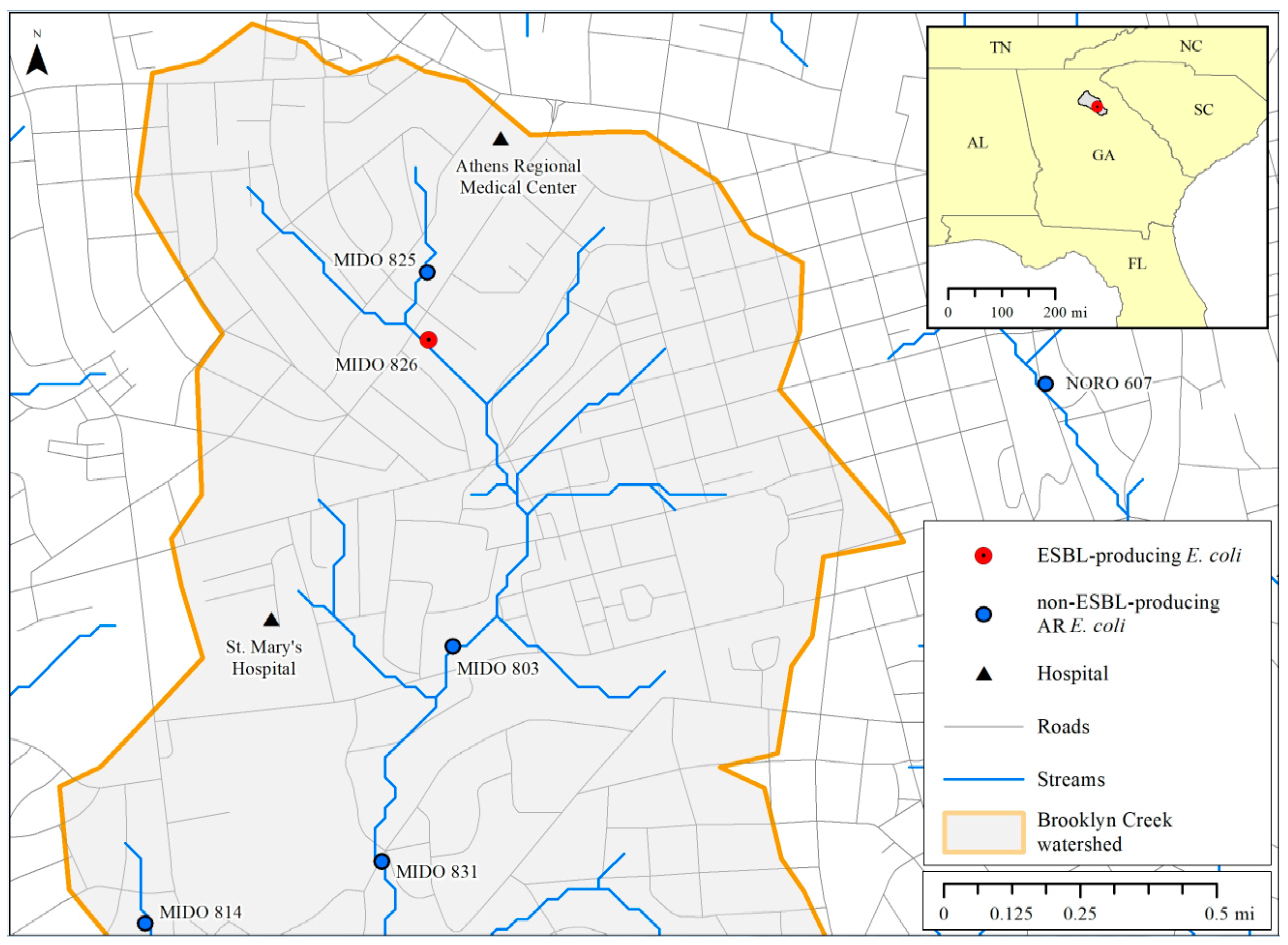
3.2. ESBL Detection
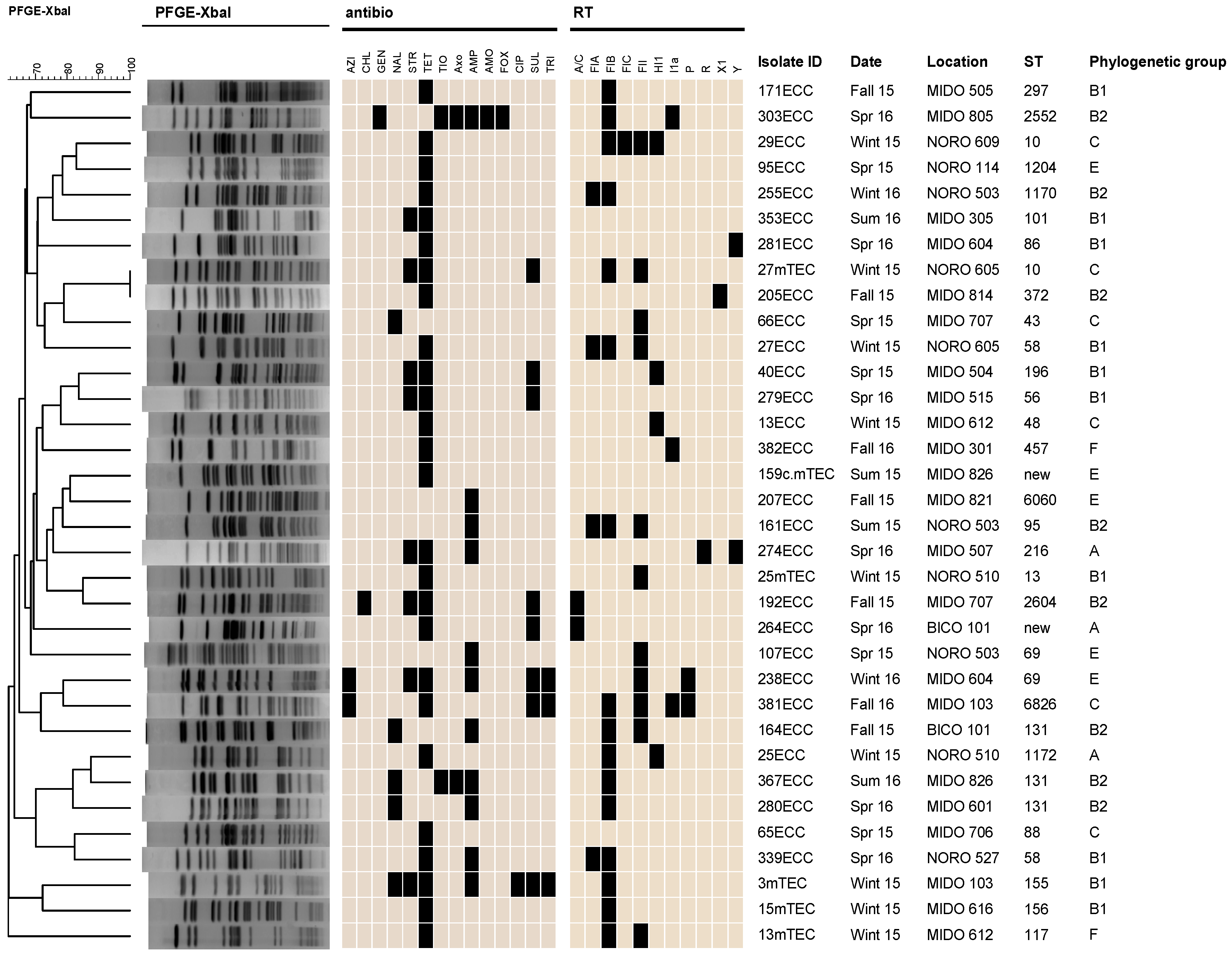
3.3. Molecular Characteristics
3.4. Mobile Genetic Elements
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Llor, C.; Bjerrum, L. Antimicrobial resistance: Risk associated with antibiotic overuse and initiatives to reduce the problem. Adv. Drug Saf. 2014, 5, 229–241. [Google Scholar] [CrossRef] [PubMed]
- von Baum, H.; Marre, R. Antimicrobial resistance of Escherichia coli and therapeutic implications. Int. J. Med. Microbiol. 2005, 295, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.P.; Bu, D.P.; Carrique-Mas, J.; Fevre, E.M.; Gilbert, M.; Grace, D.; Hay, S.I.; Jiwakanon, J.; Kakkar, M.; Kariuki, S.; et al. Antibiotic resistance is the quintessential One Health issue. Trans. R Soc. Trop. Med. Hyg. 2016, 110, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Finley, R.L.; Collignon, P.; Larsson, D.G.; McEwen, S.A.; Li, X.Z.; Gaze, W.H.; Reid-Smith, R.; Timinouni, M.; Graham, D.W.; Topp, E. The scourge of antibiotic resistance: The important role of the environment. Clin. Infect. Dis. 2013, 57, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Hiott, L.M.; Barrett, J.B.; McMillan, E.A.; House, S.L.; Humayoun, S.B.; Adams, E.S.; Jackson, C.R.; Frye, J.G. Prevalence and characterization of Escherichia coli isolated from the Upper Oconee Watershed in Northeast Georgia. Plos ONE 2018, 13, e0197005. [Google Scholar] [CrossRef] [PubMed]
- Brinas, L.; Zarazaga, M.; Saenz, Y.; Ruiz-Larrea, F.; Torres, C. β-Lactamases in Ampicillin-Resistant Escherichia coli Isolates from Foods, Humans, and Healthy Animals. Antimicrob. Agents Chemother. 2002, 46, 3156–3163. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhao, S.; White, D.G.; Schroeder, C.M.; Lu, R.; Yang, H.; McDermott, P.F.; Ayers, S.; Meng, J. Characterization of Multiple-Antimicrobial-Resistant Salmonella Serovars Isolated from Retail Meats. Appl. Environ. Microbiol. 2004, 70, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.K.; Martin, I.; Alfa, M.; Mulvey, M. Multiplex PCR for the detection of tetracycline resistant genes. Mol. Cell Probes 2001, 15, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Maynard, C.; Fairbrother, J.M.; Bekal, S.; Sanschagrin, F.; Levesque, R.C.; Brousseau, R.; Masson, L.; Lariviere, S.; Harel, J. Antimicrobial Resistance Genes in Enterotoxigenic Escherichia coli O149:K91 Isolates Obtained over a 23-Year Period from Pigs. Antimicrob. Agents Chemother. 2003, 47, 3214–3221. [Google Scholar] [CrossRef] [Green Version]
- Randall, L.P.; Cooles, S.W.; Osborn, M.K.; Piddock, L.J.; Woodward, M.J. Antibiotic resistance genes, integrons and multiple antibiotic resistance in thirty-five serotypes of Salmonella enterica isolated from humans and animals in the UK. J. Antimicrob. Chemother. 2004, 53, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Mattiello, S.P.; Drescher, G.; Barth, V.C., Jr.; Ferreira, C.A.; Oliveira, S.D. Characterization of antimicrobial resistance in Salmonella enterica strains isolated from Brazilian poultry production. Antonie Leeuwenhoek 2015, 108, 1227–1238. [Google Scholar] [CrossRef] [PubMed]
- Phuc Nguyen, M.C.; Woerther, P.L.; Bouvet, M.; Andremont, A.; Leclercq, R.; Canu, A. Escherichia coli as reservoir for macrolide resistance genes. Emerg. Infect. Dis. 2009, 15, 1648–1650. [Google Scholar] [CrossRef] [PubMed]
- Linda, M.; Weigel, C.D.S.; Fred, C. Tenover, gyrA Mutations Associated with Fluoroquinolone Resistance in Eight Species of Enterobacteriaceae. Antimicrob. Agents Chemother. 1998, 42, 2661–2667. [Google Scholar]
- Komp Lindgren, P.; Karlsson, A.; Hughes, D. Mutation Rate and Evolution of Fluoroquinolone Resistance in Escherichia coli Isolates from Patients with Urinary Tract Infections. Antimicrob. Agents Chemother. 2003, 47, 3222–3232. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 26th ed.; CLSI Supplement M100S; Clinical Laboratory Standards Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Ribot, E.M.; Fair, M.A.; Gautom, R.; Cameron, D.N.; Hunter, S.B.; Swaminathan, B.; Barrett, T.J. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 2006, 3, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Tenover, A.R.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by PFGE. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [PubMed]
- Tartof, S.Y.; Solberg, O.D.; Manges, A.R.; Riley, L.W. Analysis of a uropathogenic Escherichia coli clonal group by multilocus sequence typing. J. Clin. Microbiol. 2005, 43, 5860–5864. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Ponten, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed]
- Tankson, J.D.; Fedorka-Cray, P.J.; Jackson, C.R.; Headrick, M. Genetic relatedness of a rarely isolated Salmonella: Salmonella enterica serotype Niakhar from NARMS animal isolates. J. Antimicrob. Chemother. 2006, 57, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Hamelin, K.; Bruant, G.; El-Shaarawi, A.; Hill, S.; Edge, T.A.; Fairbrother, J.; Harel, J.; Maynard, C.; Masson, L.; Brousseau, R. Occurrence of virulence and antimicrobial resistance genes in Escherichia coli isolates from different aquatic ecosystems within the St. Clair River and Detroit River areas. Appl. Environ. Microbiol. 2007, 73, 477–484. [Google Scholar] [CrossRef]
- Titilawo, Y.; Obi, L.; Okoh, A. Antimicrobial resistance determinants of Escherichia coli isolates recovered from some rivers in Osun State, South-Western Nigeria: Implications for public health. Sci. Total Environ. 2015, 523, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Hamelin, K.; Bruant, G.; El-Shaarawi, A.; Hill, S.; Edge, T.A.; Bekal, S.; Fairbrother, J.M.; Harel, J.; Maynard, C.; Masson, L.; et al. A virulence and antimicrobial resistance DNA microarray detects a high frequency of virulence genes in Escherichia coli isolates from Great Lakes recreational waters. Appl. Environ. Microbiol. 2006, 72, 4200–4206. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, K.L.; Davies, R.H.; Threlfall, E.J. Mechanisms of quinolone resistance in Escherichia coli and Salmonella: Recent developments. Int. J. Antimicrob. Agents 2005, 25, 358–373. [Google Scholar] [CrossRef] [PubMed]
- Nicolas-Chanoine, M.H.; Bertrand, X.; Madec, J.Y. Escherichia coli ST131, an intriguing clonal group. Clin. Microbiol. Rev. 2014, 27, 543–574. [Google Scholar] [CrossRef] [PubMed]
- Canton, R.; Coque, T.M. The CTX-M beta-lactamase pandemic. Curr. Opin. Microbiol. 2006, 9, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.A. AmpC beta-lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef] [PubMed]
- Accogli, M.; Fortini, D.; Giufrè, M.; Graziani, C.; Dolejska, M.; Carattoli, A.; Cerquetti, M. IncI1 plasmids associated with the spread of CMY-2, CTX-M-1 and SHV-12 in Escherichia coli of animal and human origin. Clin. Microbiol. Infect. 2013, 19, E238–E240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Sallem, R.; Ben Slama, K.; Rojo-Bezares, B.; Porres-Osante, N.; Jouini, A.; Klibi, N.; Boudabous, A.; Saenz, Y.; Torres, C. IncI1 plasmids carrying bla(CTX-M-1) or bla(CMY-2) genes in Escherichia coli from healthy humans and animals in Tunisia. Microb. Drug Resist. 2014, 20, 495–500. [Google Scholar] [CrossRef]
- Heider, L.C.; Hoet, A.E.; Wittum, T.E.; Khaitsa, M.L.; Love, B.C.; Huston, C.L.; Morley, P.S.; Funk, J.A.; Gebreyes, W.A. Genetic and phenotypic characterization of the bla(CMY) gene from Escherichia coli and Salmonella enterica isolated from food-producing animals, humans, the environment, and retail meat. Foodborne Pathog. Dis. 2009, 6, 1235–1240. [Google Scholar] [CrossRef]
- Blaak, H.; de Kruijf, P.; Hamidjaja, R.A.; van Hoek, A.H.; de Roda Husman, A.M.; Schets, F.M. Prevalence and characteristics of ESBL-producing E. coli in Dutch recreational waters influenced by wastewater treatment plants. Vet. Microbiol. 2014, 171, 448–459. [Google Scholar] [CrossRef]
- Vivant, A.L.; Boutin, C.; Prost-Boucle, S.; Papias, S.; Hartmann, A.; Depret, G.; Ziebal, C.; Le Roux, S.; Pourcher, A.M. Free water surface constructed wetlands limit the dissemination of extended-spectrum beta-lactamase producing Escherichia coli in the natural environment. Water Res. 2016, 104, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A. Importance of integrons in the diffusion of resistance. Vet. Res. 2001, 32, 243–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, T.J.; Logue, C.M.; Johnson, J.R.; Kuskowski, M.A.; Sherwood, J.S.; Barnes, H.J.; DebRoy, C.; Wannemuehler, Y.M.; Obata-Yasuoka, M.; Spanjaard, L.; et al. Associations between multidrug resistance, plasmid content, and virulence potential among extraintestinal pathogenic and commensal Escherichia coli from humans and poultry. Foodborne Pathog. Dis. 2012, 9, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Gillings, M.; Boucher, Y.; Labbate, M.; Holmes, A.; Krishnan, S.; Holley, M.; Stokes, H.W. The evolution of class 1 integrons and the rise of antibiotic resistance. J. Bacteriol. 2008, 190, 5095–5100. [Google Scholar] [CrossRef] [PubMed]
- Roe, M.T.; Vega, E.; Pillai, S.D. Antimicrobial Resistance Markers of Class 1 and Class 2 Integron-bearing Escherichia coli from Irrigation Water and Sediments. Emerg. Infect. Dis. 2003, 9, 822–826. [Google Scholar] [CrossRef] [PubMed]
- Laroche, E.; Pawlak, B.; Berthe, T.; Skurnik, D.; Petit, F. Occurrence of antibiotic resistance and class 1, 2 and 3 integrons in Escherichia coli isolated from a densely populated estuary (Seine, France). FEMS Microbiol. Ecol. 2009, 68, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Korzeniewska, E.; Korzeniewska, A.; Harnisz, M. Antibiotic resistant Escherichia coli in hospital and municipal sewage and their emission to the environment. Ecotoxicol. Environ. Saf. 2013, 91, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Agga, G.E.; Arthur, T.M.; Durso, L.M.; Harhay, D.M.; Schmidt, J.W. Antimicrobial-Resistant Bacterial Populations and Antimicrobial Resistance Genes Obtained from Environments Impacted by Livestock and Municipal Waste. PLoS ONE 2015, 10, e0132586. [Google Scholar] [CrossRef]
- McArthur, J.V.; Fletcher, D.E.; Tuckfield, R.C.; Baker-Austin, C. Patterns of Multi-Antibiotic-Resistant Escherichia Coli from Streams with No History of Antimicrobial Inputs. Microb. Ecol. 2016, 72, 840–850. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Antimicrobial(s)/Integron | Target Gene | F Primer Sequence (5′ to 3′) | R Primer Sequence (5′ to 3′) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|---|
| β-lactams | blaCMY | AACACACTGATTGCGTCTGA | GCCTCATCGTCAGTTATTGCA | 1222 | this study |
| blaCTX-M | CACACGTGGAATTTAGGGACT | GAATGAGTTTCCCCATTCCGT | 970 | this study | |
| blaTEM | TTCTTGAAGACGAAAGGGC | ACGCTCAGTGGAACGAAAAC | 1150 | [6] | |
| tetracycline | tetA | GCGCCTTTCCTTTGGGTTCT | CCACCCGTTCCACGTTGTTA | 831 | [7] |
| tetB | CCCAGTGCTGTTGTTGTCAT | CCACCACCAGCCAATAAAAT | 723 | [7] | |
| tetC | TTGCGGGATATCGTCCATTC | CATGCCAACCCGTTCCATGT | 1019 | [7] | |
| tetG | AGCAGGTCGCTGGACACTAT | CGCGGTGTTCCACTGAAAAC | 623 | [7] | |
| tetM | GTGGACAAAGGTACAACGAG | CGGTAAAGTTCGTCACACAC | 406 | [8] | |
| trimethoprim/ | dhfr1 | CGGTCGTAACACGTTCAAGT | CTGGGGATTTCAGGAAAGTA | 220 | [7] |
| sulphamethoxazole | dhfr5 | CTGCAAAAGCGAAAAACGG | AGCAATAGTTAATGTTTGAGCTAAAG | 432 | [9] |
| dhfr12 | AAATTCCGGGTGAGCAGAAG | CCCGTTGACGGAATGGTTAG | 429 | [7] | |
| dhfr13 | GCAGTCGCCCTAAAACAAAG | GATACGTGTGACAGCGTTGA | 294 | [7] | |
| sulfisoxazole | sul1 | TCACCGAGGACTCCTTCTTC | CAGTCCGCCTCAGCAATATC | 331 | [7] |
| sul2 | CCTGTTTCGTCCGACACAGA | GAAGCGCAGCCGCAATTCAT | 435 | [7] | |
| cat1 | CTTGTCGCCTTGCGTATAAT | ATCCCAATGGCATCGTAAAG | 508 | [7] | |
| cat2 | AACGGCATGATGAACCTGAA | ATCCCAATGGCATCGTAAAG | 547 | [7] | |
| floR | CTGAGGGTGTCGTCATCTAC | GCTCCGACAATGCTGACTAT | 673 | [7] | |
| aminoglycosides | aacC2 | GGCAATAACGGAGGCAATTCGA | CTCGATGGCGACCGAGCTTCA | 450 | [7] |
| aac(3)-IVa | GATGGGCCACCTGGACTGAT | GCGCTCACAGCAGTGGTCAT | 462 | [7] | |
| aadA1 | TATCAGAGGTAGTTGGCGTCAT | GTTCCATAGCGTTAAGGTTTCATT | 484 | [10] | |
| aadA2 | TGTTGGTTACTGTGGCCGTA | GATCTCGCCTTTCACAAAGC | 622 | [10] | |
| strA | CTTGGTGATAACGGCAATTC | CCAATCGCAGATAGAAGGC | 546 | [11] | |
| strB | ATCGTCAAGGGATTGAAACC | GGATCGTAGAACATATTGGC | 509 | [11] | |
| azithromycin | mph(A) | GTGAGGAGGAGCTTCGCGAG | TGCCGCAGGACTCGGAGGTC | 403 | [12] |
| ciprofloxacin, | gyrA | CGACCTTGCGAGAGAAAT | GTTCCATCAGCCCTTCAA | 626 | [13] |
| nalidixic acid | parC | AGCGCCTTGCGTACATGAAT | GTGGTAGCGAAGAGGTGGTT | 965 | [14] |
| class I integron | intI1 | ACATGTGATGGCGACGCACGA | ATTTCTGTCCTGGCTGGCGA | 568 | [20] |
| Resistance Phenotype (No. of Isolates Tested) | Resistance Gene Detected | No. of Resistance Gene Detected (%) |
|---|---|---|
| Ampicillin (n = 11) | blaTEM-1 | 9 (81.8) |
| Third generation cephalosporins | blaCMY-2 | 1 (50.0) |
| Ceftiofur, ceftriaxone (n = 2) | blaCTX-M-15 | 1 (50.0) |
| Azithromycin (n = 2) | mph(A) | 2 (100.0) |
| Chloramphenicol (n = 1) | floR | 1 (100.0) |
| Ciprofloxacin, nalidixic acid (n = 5) | gyrA | 5 (100.0) |
| parC | 1 (20.0) | |
| Streptomycin (n = 8) | aadA1 | 1 (12.5) |
| strA | 7 (87.5) | |
| strB | 7 (87.5) | |
| Sulfisoxazole (n = 8) | sul1 | 1 (12.5) |
| sul2 | 7 (87.5) | |
| Tetracycline (n = 26) | tetA | 16 (61.5) |
| tetB | 9 (34.6) | |
| tetC | 1 (3.8) | |
| Trimethoprim/sulphamethoxazole (n = 3) | dhfr5 | 2 (66.7) |
| dhfr12 | 1 (33.3) | |
| Class I integron (n = 34) | intI1 | 3 (8.8) |
| Isolate ID | Resistance to (MIC in µg/mL) | parC | gyrA |
|---|---|---|---|
| 3 mTEC | Ciprofloxacin (>4) | Ser-80 → Ile | Val-37 → Leu |
| Nalidixic acid (>32) | Ser-83 → Leu | ||
| Asp-87 → Asn | |||
| 66 ECC | Nalidixic acid (>32) | No mutation | Ser-83 → Leu |
| 164 ECC | Nalidixic acid (>32) | Lys-247 → Glu | Ser-83 → Leu |
| Lys-162 → Gln | |||
| 280 ECC | Nalidixic acid (>32) | Ala-192 → Gly | Ser-83 → Leu |
| 367 ECC | Nalidixic acid (>32) | Lys-247 → Glu | Ser-83 → Leu |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, S.; Nguyen, H.A.T.; McDonald, J.M.; Woodley, T.A.; Hiott, L.M.; Barrett, J.B.; Jackson, C.R.; Frye, J.G. Genetic Characterization of Antimicrobial-Resistant Escherichia coli Isolated from a Mixed-Use Watershed in Northeast Georgia, USA. Int. J. Environ. Res. Public Health 2019, 16, 3761. https://doi.org/10.3390/ijerph16193761
Cho S, Nguyen HAT, McDonald JM, Woodley TA, Hiott LM, Barrett JB, Jackson CR, Frye JG. Genetic Characterization of Antimicrobial-Resistant Escherichia coli Isolated from a Mixed-Use Watershed in Northeast Georgia, USA. International Journal of Environmental Research and Public Health. 2019; 16(19):3761. https://doi.org/10.3390/ijerph16193761
Chicago/Turabian StyleCho, Sohyun, Hoang Anh Thi Nguyen, Jacob M. McDonald, Tiffanie A. Woodley, Lari M. Hiott, John B. Barrett, Charlene R. Jackson, and Jonathan G. Frye. 2019. "Genetic Characterization of Antimicrobial-Resistant Escherichia coli Isolated from a Mixed-Use Watershed in Northeast Georgia, USA" International Journal of Environmental Research and Public Health 16, no. 19: 3761. https://doi.org/10.3390/ijerph16193761
APA StyleCho, S., Nguyen, H. A. T., McDonald, J. M., Woodley, T. A., Hiott, L. M., Barrett, J. B., Jackson, C. R., & Frye, J. G. (2019). Genetic Characterization of Antimicrobial-Resistant Escherichia coli Isolated from a Mixed-Use Watershed in Northeast Georgia, USA. International Journal of Environmental Research and Public Health, 16(19), 3761. https://doi.org/10.3390/ijerph16193761