Urban Air Pollution Particulates Suppress Human T-Cell Responses to Mycobacterium Tuberculosis

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Approval
2.2. Human Subjects
2.3. Preparation of PBMC
2.4. Collection of PM2.5 and Preparation for in Vitro Exposure of PBMC
2.5. Preparation of M.tb for in Vitro Infection
2.6. PM2.5 Exposures of PBMC
2.7. PM2.5 Exposures, Infection with M.tb, and CFU Assays
2.8. Lactate Dehydrogenase (LDH) Assay
2.9. Cell Surface and Intracellular Cytokine Immunostaining
2.10. Flow Cytometry Analysis
2.11. Apoptosis Assay by Annexin V Staining
2.12. Transmission Electron Microscopy
2.13. Evaluation of Transcription Factor T-Bet by Western Blot
2.14. Statistical Analysis
3. Results
3.1. Uptake of PM2.5 and M.tb in PBMC
3.2. Cytotoxic Effects of PM2.5 in Human PBMC
3.3. PM2.5 Exposure Effects on Apoptosis of Mononuclear Cells
3.4. PM2.5 Exposure Decreases Intracellular Growth Control of M.tb by PBMC
3.5. PM2.5 Exposure Downregulates the Expression of CD69 on T Cells
3.6. PM2.5 Exposure Decreases M.tb-induced IFN-γ Expression in PBMC
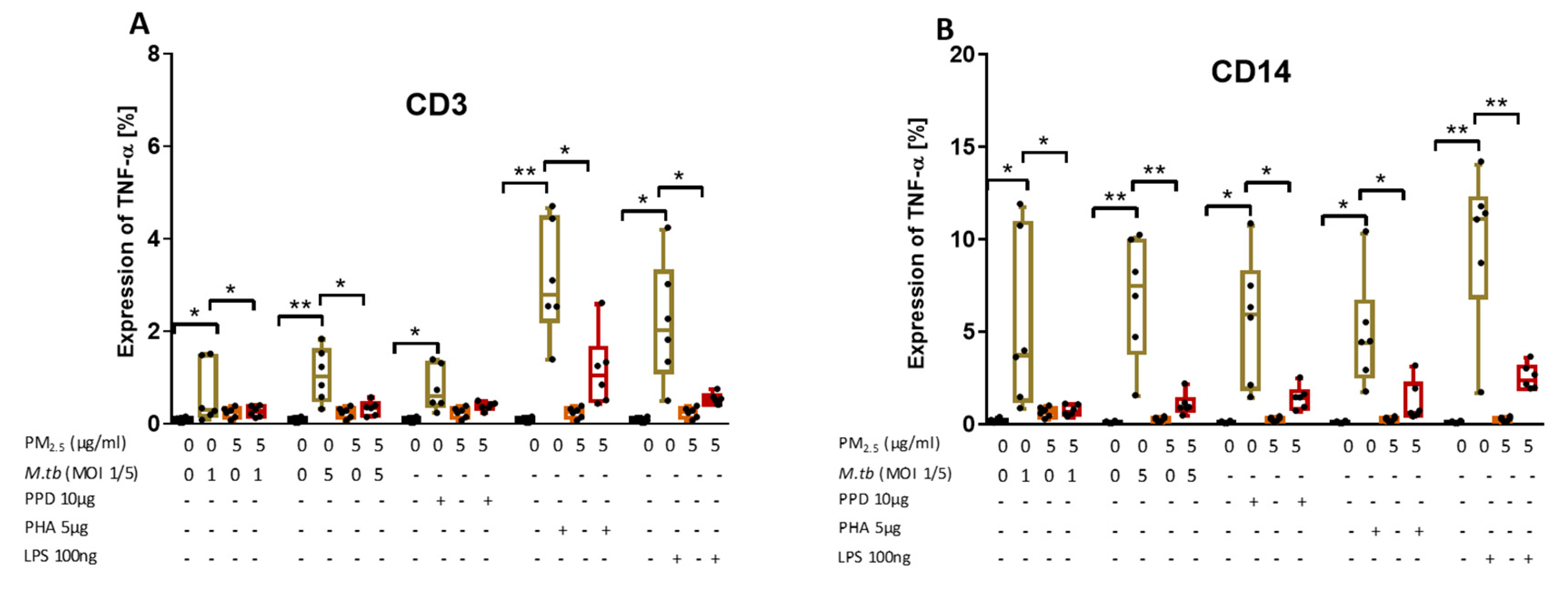
3.7. PM2.5 Exposure Decreases M.tb-Induced TNF-α Expression in CD3+ and CD14+ PBMC
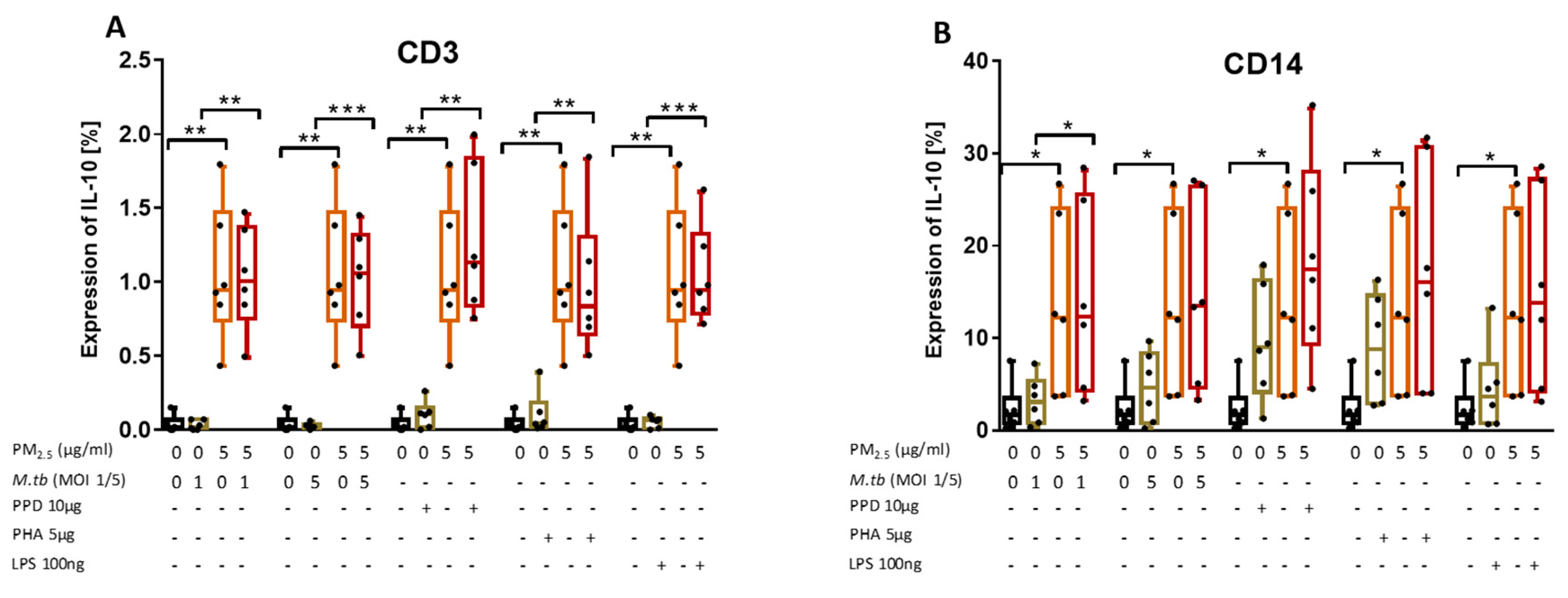
3.8. PM2.5 Exposure Induces the Production of IL-10 in M.tb-Infected PBMC
3.9. PM2.5 Exposure Reduces the Expression of Transcription Factor T-bet
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jasenosky, L.D.; Scriba, T.J.; Hanekom, W.A.; Goldfeld, A.E. T cells and adaptive immunity to Mycobacterium tuberculosis in humans. Immunol. Rev. 2015, 264, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.M. T cells in mycobacterial infection and disease. Curr. Opin. Immunol. 2009, 21, 378–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, C.G.; Bean, A.G.; Hooi, H.; Briscoe, H.; Britton, W.J. Increase in gamma interferon-secreting CD8(+), as well as CD4(+), T cells in lungs following aerosol infection with Mycobacterium tuberculosis. Infect. Immun. 1999, 67, 3242–3247. [Google Scholar] [PubMed]
- Allie, N.; Grivennikov, S.I.; Keeton, R.; Hsu, N.J.; Bourigault, M.L.; Court, N.; Fremond, C.; Yeremeev, V.; Shebzukhov, Y.; Ryffel, B.; et al. Prominent role for T cell-derived Tumour Necrosis Factor for sustained control of Mycobacterium tuberculosis infection. Sci. Rep. 2013, 3, 1809. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.T.; Modlin, R.L. Human macrophage host defense against Mycobacterium tuberculosis. Curr. Opin. Immunol. 2008, 20, 371–376. [Google Scholar] [CrossRef]
- Moreno, C.; Taverne, J.; Mehlert, A.; Bate, C.A.; Brealey, R.J.; Meager, A.; Rook, G.A.; Playfair, J.H. Lipoarabinomannan from Mycobacterium-Tuberculosis Induces the Production of Tumor Necrosis Factor from Human and Murine Macrophages. Clin. Exp. Immunol. 1989, 76, 240–245. [Google Scholar] [PubMed]
- Flesch, I.E.; Kaufmann, S.H. Activation of tuberculostatic macrophage functions by gamma interferon, interleukin-4, and tumor necrosis factor. Infect. Immun. 1990, 58, 2675–2677. [Google Scholar] [Green Version]
- Cavalcanti, Y.V.N.; Brelaz, M.C.A.; Neves, J.K.D.A.L.; Ferraz, J.C.; Pereira, V.R.A. Role of TNF-Alpha, IFN-Gamma, and IL-10 in the Development of Pulmonary Tuberculosis. Pulm. Med. 2012, 2012, 745483. [Google Scholar] [CrossRef]
- Zhang, M.; Gong, J.; Iyer, D.V.; Jones, B.E.; Modlin, R.L.; Barnes, P.F. T cell cytokine responses in persons with tuberculosis and human immunodeficiency virus infection. J. Clin. Investig. 1994, 94, 2435–2442. [Google Scholar] [CrossRef]
- Gong, J.H.; Zhang, M.; Modlin, R.L.; Linsley, P.S.; Iyer, D.; Lin, Y.; Barnes, P.F. Interleukin-10 downregulates Mycobacterium tuberculosis-induced Th1 responses and CTLA-4 expression. Infect. Immun. 1996, 64, 913–918. [Google Scholar] [Green Version]
- Liang, L.; Zhao, Y.L.; Yue, J.; Liu, J.F.; Han, M.; Wang, H.; Xiao, H. Interleukin-10 gene promoter polymorphisms and their protein production in pleural fluid in patients with tuberculosis. FEMS Immunol. Med. Microbiol. 2011, 62, 84–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mbow, M.; Santos, N.S.; Camara, M.; Ba, A.; Niang, A.; Daneau, G.; Wade, D.; Diallo, A.A.; Toupane, M.; DiakhatéM.;Lèye, N. HIV and Tuberculosis co-infection impacts T-cell activation markers but not the numbers subset of regulatory T-cells in HIV-1 infected patients. Afr. J. Lab. Med. 2013, 2, 76. [Google Scholar] [CrossRef] [PubMed]
- Anuradha, R.; Munisankar, S.; Bhootra, Y.; Kumar, N.P.; Dolla, C.; Kumaran, P.; Babu, S. Coexistent Malnutrition Is Associated with Perturbations in Systemic and Antigen-Specific Cytokine Responses in Latent Tuberculosis Infection. Clin. Vaccine Immunol. 2016, 23, 339–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anibarro, L.; Pena, A. Tuberculosis in patients with haematological malignancies. Medit. J. Hematol. Infect Dis. 2014, 6, e2014026. [Google Scholar] [CrossRef]
- Harris, J.; Keane, J. How tumour necrosis factor blockers interfere with tuberculosis immunity. Clin. Exp. Immunol. 2010, 161, 1–9. [Google Scholar] [CrossRef]
- O’Leary, S.M.; Coleman, M.M.; Chew, W.M.; Morrow, C.; McLaughlin, A.M.; Gleeson, L.E.; O’Sullivan, M.P.; Keane, J. Cigarette smoking impairs human pulmonary immunity to Mycobacterium tuberculosis. Am. J. Respir. Crit. Care Med. 2014, 190, 1430–1436. [Google Scholar] [CrossRef]
- García-Sancho, M.C.; García-García, L.; Báez-Saldaña, R.; Ponce-de-León, A.; Sifuentes-Osornio, J.; Bobadilla-del-Valle, M.; Ferreyra-Reyes, L.; Cano-Arellano, B.; Canizales-Quintero, S.; Palacios-Merino Ldel, C.; et al. Indoor pollution as an occupational risk factor for tuberculosis among women: A population-based, gender oriented, case-control study in Southern Mexico. Rev. Investig. Clin. 2009, 61, 392–398. [Google Scholar]
- Blount, R.J.; Pascopella, L.; Catanzaro, D.G.; Barry, P.M.; English, P.B.; Segal, M.R.; Flood, J.; Meltzer, D.; Jones, B.; Balmes, J.; et al. Traffic-Related Air Pollution and All-Cause Mortality during Tuberculosis Treatment in California. Environ. Health Perspect. 2017, 125, 097026. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Song, Y.; Sarkar, S.; Kipen, H.M.; Laumbach, R.J.; Zhang, J.; Strickland, P.A.O.; Gardner, C.R.; Schwander, S. Suppression of the NF-kappaB pathway by diesel exhaust particles impairs human antimycobacterial immunity. J. Immunol. 2012, 188, 2778–2793. [Google Scholar] [CrossRef]
- Rivas-Santiago, C.E.; Sarkar, S.; Cantarella, P.; Osornio-Vargas, Á.; Quintana-Belmares, R.; Meng, Q.; Kirn, T.J.; Strickland, P.O.; Chow, J.C.; Watson, J.G.; et al. Air pollution particulate matter alters antimycobacterial respiratory epithelium innate immunity. Infect. Immun. 2015, 83, 2507–2517. [Google Scholar] [CrossRef]
- Torres, M.; Carranza, C.; Sarkar, S.; Gonzalez, Y.; Vargas, A.O.; Black, K.; Meng, Q.; Quintana-Belmares, R.; Hernandez, M.; Garcia, J.J.F.A.; et al. Urban airborne particle exposure impairs human lung and blood Mycobacterium tuberculosis immunity. Thorax 2019, 74, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Carranza, C.; Juárez, E.; Torres, M.; Ellner, J.J.; Sada, E.; Schwander, S.K. Mycobacterium tuberculosis growth control by lung macrophages and CD8 cells from patient contacts. Am. J. Respir. Crit Care Med. 2006, 173, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Van Crevel, R.; Ottenhoff, T.H.; Van Der Meer, J.W. Innate immunity to Mycobacterium tuberculosis. Clin. Microbiol. Rev. 2002, 15, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Schwander, S.; Dheda, K. Human lung immunity against Mycobacterium tuberculosis: Insights into pathogenesis and protection. Am. J. Respir. Crit. Care Med. 2011, 183, 696–707. [Google Scholar] [CrossRef]
- Wolf, A.J.; Desvignes, L.; Linas, B.; Banaiee, N.; Tamura, T.; Takatsu, K.; Ernst, J.D. Initiation of the adaptive immune response to Mycobacterium tuberculosis depends on antigen production in the local lymph node, not the lungs. J. Exp. Med. 2008, 205, 105–115. [Google Scholar] [CrossRef]
- Xing, Z.; Wang, J.; Croitoru, K.; Wakeham, J. Protection by CD4 or CD8 T cells against pulmonary Mycobacterium bovis bacillus Calmette-Guerin infection. Infect. Immun. 1998, 66, 5537–5542. [Google Scholar]
- Moreira-Teixeira, L.; Redford, P.S.; Stavropoulos, E.; Ghilardi, N.; Maynard, C.L.; Weaver, C.T.; do Rosário, A.P.F.; Wu, X.; Langhorne, J.; O’Garra, A. T Cell-Derived IL-10 Impairs Host Resistance to Mycobacterium tuberculosis Infection. J. Immunol. 2017, 199, 613–623. [Google Scholar] [CrossRef]
- Boussiotis, V.A.; Tsai, E.Y.; Yunis, E.J.; Thim, S.; Delgado, J.C.; Dascher, C.C.; Berezovskaya, A.; Rousset, D.; Reynes, J.M.; Goldfeld, A.E. IL-10-producing T cells suppress immune responses in anergic tuberculosis patients. J. Clin. Investig. 2000, 105, 1317–1325. [Google Scholar] [CrossRef]
- Leung, C.C.; Lam, T.H.; Ho, K.S.; Yew, W.W.; Tam, C.M.; Chan, W.M.; Law, W.S.; Chan, C.K.; Chang, K.C.; Au, K.F. Passive smoking and tuberculosis. Arch. Intern. Med. 2010, 170, 287–292. [Google Scholar] [CrossRef]
- Patra, S.; Sharma, S.; Behera, D. Passive smoking, indoor air pollution and childhood tuberculosis: A case control study. Indian J. Tuberc. 2012, 59, 151–155. [Google Scholar]
- Smith, G.S.; Schoenbach, V.J.; Richardson, D.B.; Gammon, M.D. Particulate air pollution and susceptibility to the development of pulmonary tuberculosis disease in North Carolina: An ecological study. Int. J. Environ. Health Res. 2014, 24, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Furuyama, A.; Kanno, S.; Kobayashi, T.; Hirano, S. Extrapulmonary translocation of intratracheally instilled fine and ultrafine particles via direct and alveolar macrophage-associated routes. Arch. Toxicol. 2009, 83, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Saenen, N.D.; Bové, H.; Steuwe, C.; Roeffaers, M.B.; Provost, E.B.; Lefebvre, W.; Vanpoucke, C.; Ameloot, M.; Nawrot, T.S. Children’s Urinary Environmental Carbon Load. A Novel Marker Reflecting Residential Ambient Air Pollution Exposure? Am. J. Respir. Crit. Care Med. 2017, 196, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Gao, Z.; Tian, Z.; Xie, Y.; Xin, F.; Jiang, R.; Kan, H.; Song, W. The biological effects of individual-level PM(2.5) exposure on systemic immunity and inflammatory response in traffic policemen. Occup. Environ. Med. 2013, 70, 426–431. [Google Scholar] [CrossRef]
- Pierdominici, M.; Maselli, A.; Cecchetti, S.; Tinari, A.; Mastrofrancesco, A.; Alfè, M.; Gargiulo, V.; Beatrice, C.; Di Blasio, G.; Carpinelli, G.; et al. Diesel exhaust particle exposure in vitro impacts T lymphocyte phenotype and function. Part. Fibre Toxicol. 2014, 11, 74. [Google Scholar] [CrossRef]
- Bonay, M.; Chambellan, A.; Grandsaigne, M.; Aubier, M.; Soler, P. Effects of diesel particles on the control of intracellular mycobacterial growth by human macrophages in vitro. FEMS Immunol. Med. Microbiol. 2006, 46, 419–425. [Google Scholar] [CrossRef]
- Saxena, R.K.; Saxena, Q.B.; Weissman, D.N.; Simpson, J.P.; Bledsoe, T.A.; Lewis, D.M. Effect of diesel exhaust particulate on bacillus Calmette-Guerin lung infection in mice and attendant changes in lung interstitial lymphoid subpopulations and IFNgamma response. Toxicol. Sci. 2003, 73, 66–71. [Google Scholar] [CrossRef]
- Hiramatsu, K.; Saito, Y.; Sakakibara, K.; Azuma, A.; Kudoh, S.; Takizawa, H.; Sugawara, I. The effects of inhalation of diesel exhaust on murine mycobacterial infection. Exp. Lung Res. 2005, 31, 405–415. [Google Scholar] [CrossRef]
- Testi, R.; D’Ambrosio, D.; De Maria, R.; Santoni, A. The CD69 receptor: A multipurpose cell-surface trigger for hematopoietic cells. Immunol. Today 1994, 15, 479–483. [Google Scholar] [CrossRef]
- Avgustin, B.; Kotnik, V.; Škoberne, M.; Malovrh, T.; Skralovnik-Stern, A.; Terčelj, M. CD69 expression on CD4+ T lymphocytes after in vitro stimulation with tuberculin is an indicator of immune sensitization against Mycobacterium tuberculosis antigens. Clin. Diagn. Lab. Immunol. 2005, 12, 101–106. [Google Scholar] [CrossRef]
- Mackay, L.K.; Braun, A.; Macleod, B.L.; Collins, N.; Tebartz, C.; Bedoui, S.; Carbone, F.R.; Gebhardt, T. Cutting edge: CD69 interference with sphingosine-1-phosphate receptor function regulates peripheral T cell retention. J. Immunol. 2015, 194, 2059–2063. [Google Scholar] [CrossRef] [PubMed]
- Manzano-León, N.; Serrano-Lomelin, J.; Sánchez, B.N.; Quintana-Belmares, R.; Vega, E.; Vázquez-López, I.; Rojas-Bracho, L.; López-Villegas, M.T.; Vadillo-Ortega, F.; De Vizcaya-Ruiz, A.; et al. TNFalpha and IL-6 Responses to Particulate Matter in Vitro: Variation According to PM Size, Season, and Polycyclic Aromatic Hydrocarbon and Soil Content. Environ. Health Perspect. 2016, 124, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Manzano-León, N.; Quintana, R.; Sánchez, B.; Serrano, J.; Vega, E.; Vázquez-López, I.; Rojas-Bracho, L.; López-Villegas, T.; O’Neill, M.S.; Vadillo-Ortega, F.; et al. Variation in the composition and in vitro proinflammatory effect of urban particulate matter from different sites. J. Biochem. Mol. Toxicol. 2013, 27, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Rivas-Santiago, C.E.; Ibironke, O.A.; Carranza, C.; Meng, Q.; Osornio-Vargas, Á.; Zhang, J.; Torres, M.; Chow, J.C.; Watson, J.G.; et al. Season and size of urban particulate matter differentially affect cytotoxicity and human immune responses to Mycobacterium tuberculosis. PLoS ONE 2019, 14, e0219122. [Google Scholar] [CrossRef]
- Chan, R.C.F.; Wang, M.; Li, N.; Yanagawa, Y.; Onoé, K.; Lee, J.J.; Nel, A.E. Pro-oxidative diesel exhaust particle chemicals inhibit LPS-induced dendritic cell responses involved in T-helper differentiation. J. Allergy Clin. Immunol. 2006, 118, 455–465. [Google Scholar] [CrossRef]
- Lin, P.L.; Flynn, J.L. CD8 T cells and Mycobacterium tuberculosis infection. Semin. Immunopathol. 2015, 37, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Allen, M.; Bailey, C.; Cahatol, I.; Dodge, L.; Yim, J.; Kassissa, C.; Luong, J.; Kasko, S.; Pandya, S.; Venketaraman, V. Mechanisms of Control of Mycobacterium tuberculosis by NK Cells: Role of Glutathione. Front. Immunol. 2015, 6, 508. [Google Scholar] [CrossRef] [Green Version]
- Szabo, S.J.; Sullivan, B.M.; Stemmann, C.; Satoskar, A.R.; Sleckman, B.P.; Glimcher, L.H. Distinct effects of T-bet in TH1 lineage commitment and IFN-gamma production in CD4 and CD8 T cells. Science 2002, 295, 338–342. [Google Scholar] [CrossRef]
- Sullivan, B.M.; Jobe, O.; Lazarevic, V.; Vasquez, K.; Bronson, R.; Glimcher, L.H.; Kramnik, I. Increased susceptibility of mice lacking T-bet to infection with Mycobacterium tuberculosis correlates with increased IL-10 and decreased IFN-gamma production. J. Immunol. 2005, 175, 4593–4602. [Google Scholar] [CrossRef]
- Saraiva, M.; O’garra, A. The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Almeida, A.S.; Lago, P.M.; Boechat, N.; Huard, R.C.; Lazzarini, L.C.; Santos, A.R.; Nociari, M.; Zhu, H.; Perez-Sweeney, B.M.; Bang, H.; et al. Tuberculosis is associated with a down-modulatory lung immune response that impairs Th1-type immunity. J. Immunol. 2009, 183, 718–731. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Kong, Y.; Barnes, P.F.; Huang, F.F.; Klucar, P.; Wang, X.; Samten, B.; Sengupta, M.; Machona, B.; Donis, R.; et al. Exposure to cigarette smoke inhibits the pulmonary T-cell response to influenza virus and Mycobacterium tuberculosis. Infect. Immun. 2011, 79, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.P.; Scriba, T.J.; Joseph, S.; Harbacheuski, R.; Murray, R.A.; Gelderbloem, S.J.; Hawkridge, A.; Hussey, G.D.; Maecker, H.; Kaplan, G.; et al. Bacillus Calmette-Guerin vaccination of human newborns induces T cells with complex cytokine and phenotypic profiles. J. Immunol. 2008, 180, 3569–3577. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T Cell Subset | M.tb MOI | p-Value vs. Uninfected, Unstimulated (No PM2.5) PBMC |
|---|---|---|
| CD3+ | 1 | 0.0001 |
| CD3+ | 5 | <0.0001 |
| CD4+ | 1 | 0.0005 |
| CD4+ | 5 | <0.0001 |
| CD8+ | 1 | 0.0026 |
| CD8+ | 5 | 0.0002 |
| CD8+CD16+ | 1 | 0.0014 |
| CD8+CD16+ | 5 | 0.0002 |
| T Sell Subset | PM2.5 Pre-Exposure Followed by M.tb Infection | p-Value vs. MOI1 and MOI5 Combined |
|---|---|---|
| CD3+ | PM2.5 + M.tb | 0.0318 |
| CD4+ | PM2.5 + M.tb | 0.0268 |
| CD8+ | PM2.5 + M.tb | 0.0814 |
| CD8+CD16+ | PM2.5 + M.tb | 0.0522 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibironke, O.; Carranza, C.; Sarkar, S.; Torres, M.; Choi, H.T.; Nwoko, J.; Black, K.; Quintana-Belmares, R.; Osornio-Vargas, Á.; Ohman-Strickland, P.; et al. Urban Air Pollution Particulates Suppress Human T-Cell Responses to Mycobacterium Tuberculosis. Int. J. Environ. Res. Public Health 2019, 16, 4112. https://doi.org/10.3390/ijerph16214112
Ibironke O, Carranza C, Sarkar S, Torres M, Choi HT, Nwoko J, Black K, Quintana-Belmares R, Osornio-Vargas Á, Ohman-Strickland P, et al. Urban Air Pollution Particulates Suppress Human T-Cell Responses to Mycobacterium Tuberculosis. International Journal of Environmental Research and Public Health. 2019; 16(21):4112. https://doi.org/10.3390/ijerph16214112
Chicago/Turabian StyleIbironke, Olufunmilola, Claudia Carranza, Srijata Sarkar, Martha Torres, Hyejeong Theresa Choi, Joyce Nwoko, Kathleen Black, Raul Quintana-Belmares, Álvaro Osornio-Vargas, Pamela Ohman-Strickland, and et al. 2019. "Urban Air Pollution Particulates Suppress Human T-Cell Responses to Mycobacterium Tuberculosis" International Journal of Environmental Research and Public Health 16, no. 21: 4112. https://doi.org/10.3390/ijerph16214112
APA StyleIbironke, O., Carranza, C., Sarkar, S., Torres, M., Choi, H. T., Nwoko, J., Black, K., Quintana-Belmares, R., Osornio-Vargas, Á., Ohman-Strickland, P., & Schwander, S. (2019). Urban Air Pollution Particulates Suppress Human T-Cell Responses to Mycobacterium Tuberculosis. International Journal of Environmental Research and Public Health, 16(21), 4112. https://doi.org/10.3390/ijerph16214112