Different Associations between DC-SIGN Promoter-336G/A (rs4804803) Polymorphism with Severe Dengue in Asians and South-Central Americans: a Meta-Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy and Inclusion Criteria
2.2. Data Extraction
2.3. Quality Assessment
2.4. Statistical Analysis
3. Results
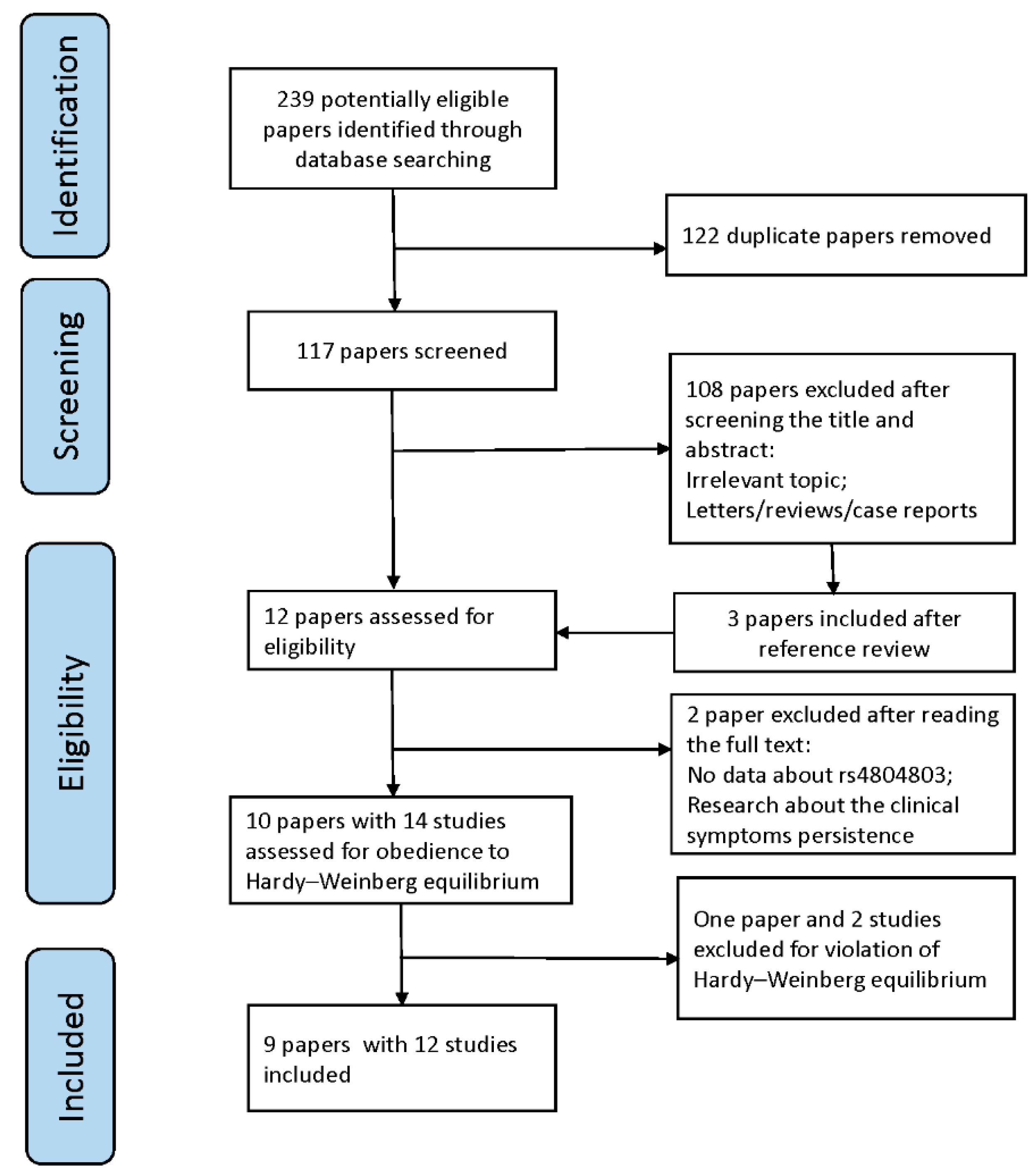
3.1. Study Inclusion and Characteristics
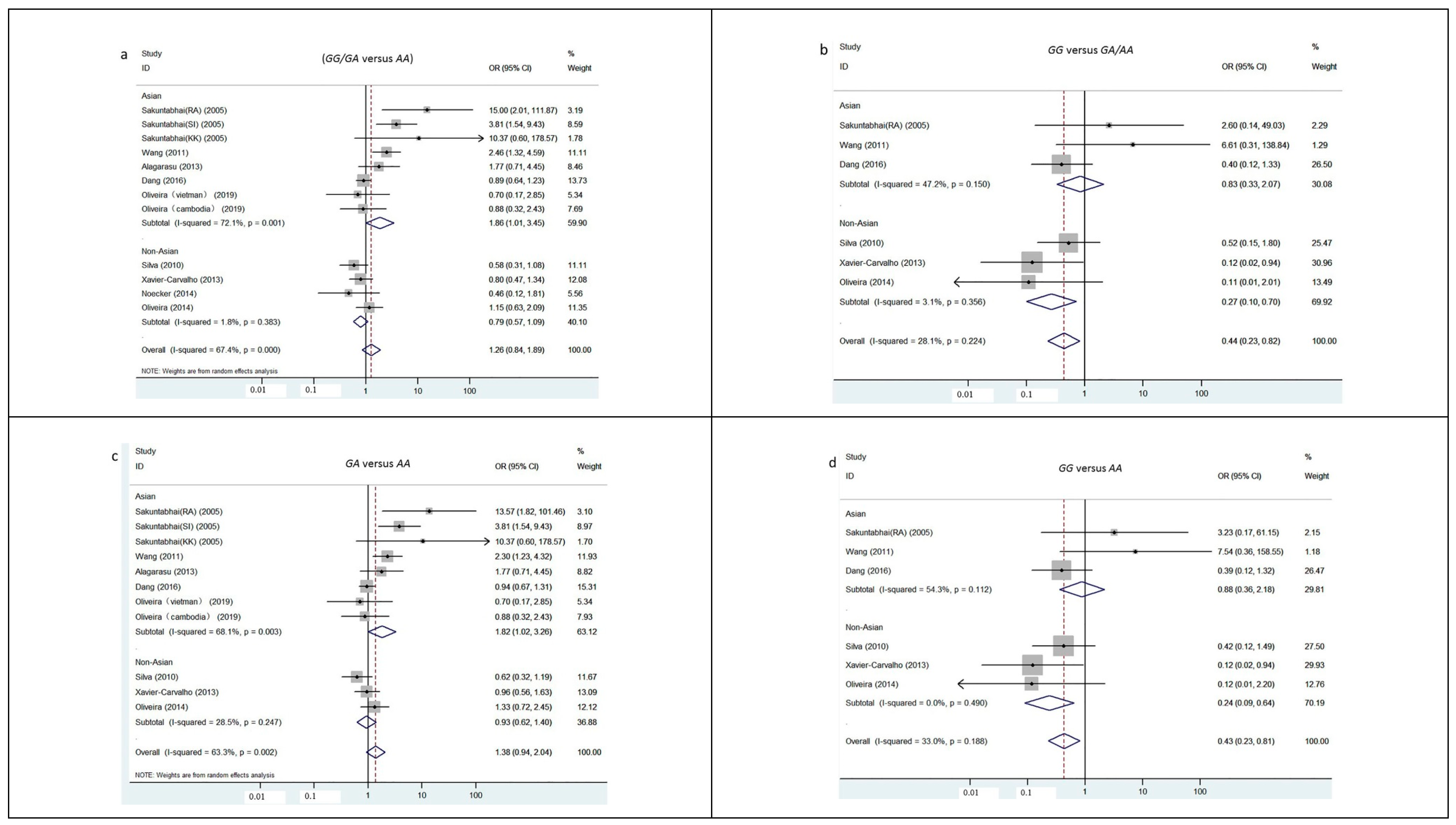
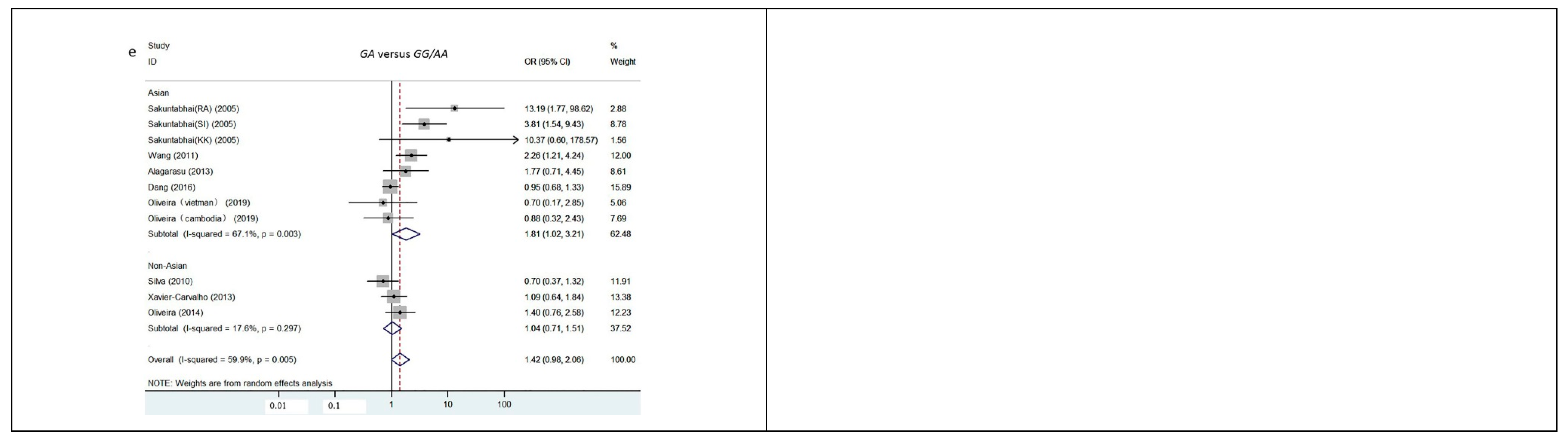
3.2. Quantitative Data Synthesis
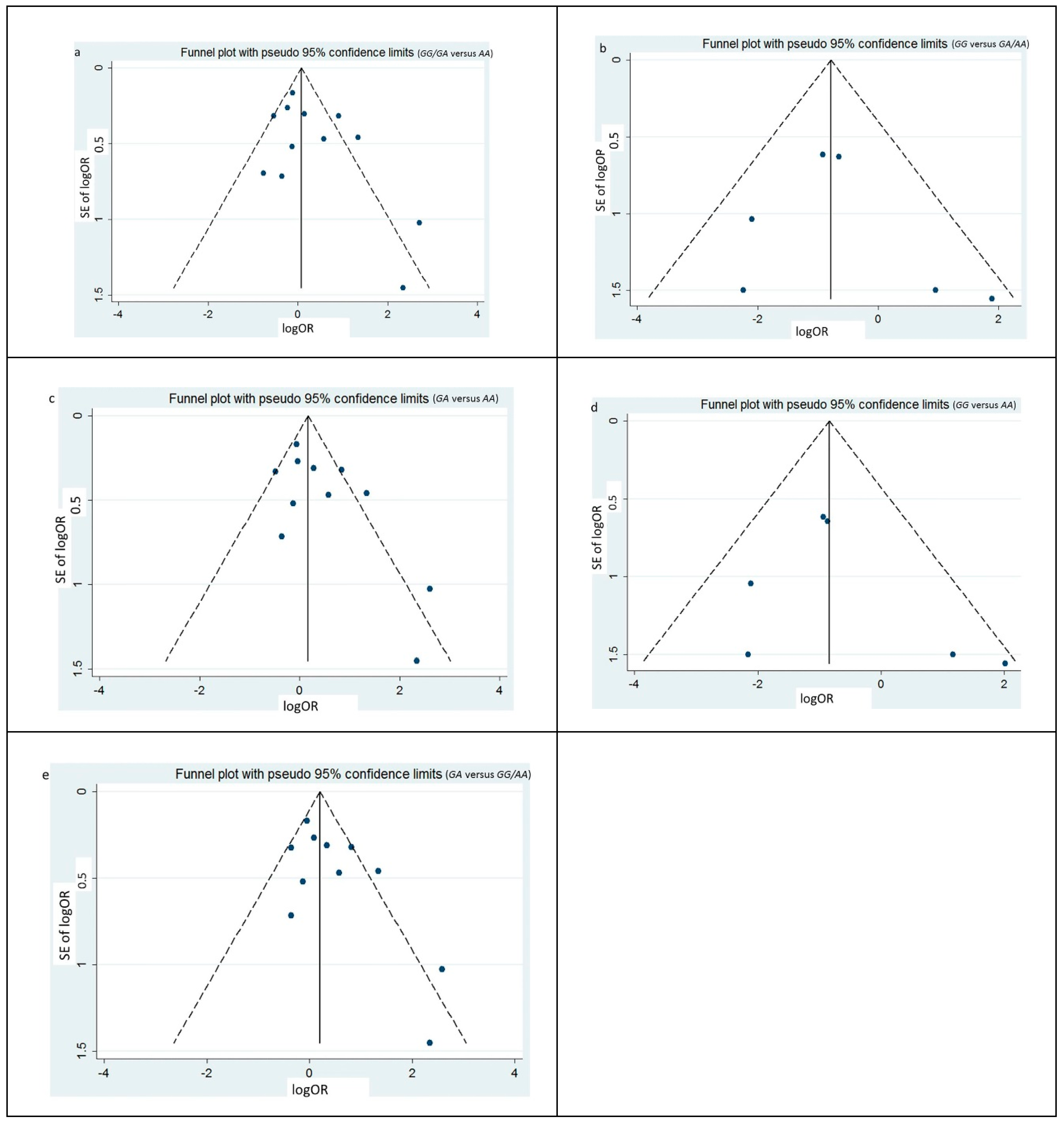
3.3. Heterogeneity and Publication Bias
3.4. Sensitivity Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. Dengue Guidelines for Diagnosis, Treatment, Prevention and Control [EB]. Available online: https://www.who.int/neglected_diseases/resources/9789241547871/en/ (accessed on 1 December 2018).
- Stanaway, J.D.; Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Coffeng, L.E.; Brady, O.J.; Hay, S.I.; Bedi, N.; Bensenor, I.M.; Castañeda-Orjuela, C.A.; et al. The global burden of dengue: An analysis from the global burden of disease study 2013. Lancet Infect. Dis. 2016, 6, 712–723. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [Green Version]
- Xavier-Carvalho, C.; Cardoso, C.C.; de Souza Kehdy, F.; Pacheco, A.G.; Moraes, M.O. Host genetics and dengue fever. Infect. Genet. Evol. 2017, 56, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Murugananthan, K.; Subramaniyam, S.; Kumanan, T.; Owens, L.; Ketheesan, N.; Noordeen, F. Blood group ab is associated with severe forms of dengue virus infection. VirusDisease 2018, 29, 103–105. [Google Scholar] [CrossRef] [PubMed]
- Nunes, P.C.G.; de Filippis, A.M.B.; Lima, M.Q.D.R.; Faria, N.R.D.C.; de Bruycker-Nogueira, F.; Santos, J.B.; Heringer, M.; Chouin-Carneiro, T.; Couto-Lima, D.; de Santis Gonçalves, B.; et al. 30 years of dengue fatal cases in Brazil: A laboratorial-based investigation of 1047 cases. BMC Infect. Dis. 2018, 18, 346. [Google Scholar] [CrossRef] [PubMed]
- Suppiah, J.; Ching, S.; Amin-Nordin, S.; Mat-Nor, L.; Ahmad-Najimudin, N.; Low, G.K.; Abdul-Wahid, M.; Thayan, R.; Chee, H. Clinical manifestations of dengue in relation to dengue serotype and genotype in Malaysia: A retrospective observational study. PLoS Negl. Trop. Dis. 2018, 12, e6817. [Google Scholar] [CrossRef]
- Huy, N.T.; Van Giang, T.; Thuy, D.H.; Kikuchi, M.; Hien, T.T.; Zamora, J.; Hirayama, K. Factors associated with dengue shock syndrome: A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2013, 7, e2412. [Google Scholar] [CrossRef]
- Moraes, G.H.; de Fatima Duarte, E.; Duarte, E.C. Determinants of mortality from severe dengue in Brazil: A population-based case-control study. Am. J. Trop. Med. Hyg. 2013, 88, 670–676. [Google Scholar] [CrossRef]
- Agrawal, V.K.; Prusty, B.; Reddy, C.S.; Mohan, R.G.; Agrawal, R.K.; Sekher, S.B.V. Clinical profile and predictors of severe dengue disease: A study from South India. Casp. J. Intern. Med. 2018, 9, 334–340. [Google Scholar]
- Pang, J.; Hsu, J.P.; Yeo, T.W.; Leo, Y.S.; Lye, D.C. Diabetes, cardiac disorders and asthma as risk factors for severe organ involvement among adult dengue patients: A matched case-control study. Sci. Rep. 2017, 7, 39872. [Google Scholar] [CrossRef] [Green Version]
- Thanachartwet, V.; Oer-areemitr, N.; Chamnanchanunt, S.; Sahassananda, D.; Jittmittraphap, A.; Suwannakudt, P.; Desakorn, V.; Wattanathum, A. Identification of clinical factors associated with severe dengue among Thai adults: A prospective study. BMC Infect. Dis. 2015, 15, 420. [Google Scholar] [CrossRef] [PubMed]
- Machado, C.R.; Machado, E.S.; Rohloff, R.D.; Azevedo, M.; Campos, D.P.; de Oliveira, R.B.; Brasil, P. Is pregnancy associated with severe dengue? A review of data from the Rio De Janeiro surveillance information system. PLoS Neglect. Trop. Dis. 2013, 7, e2217. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.C.; Castro, D.B.; Albuquerque, B.C.; Sampaio, V.D.S.; Passos, R.A.; Costa, C.F.; Sadahiro, M.; Braga, J.U. Mortality predictors in patients with severe dengue in the state of Amazonas, Brazil. PLoS ONE 2016, 11, e161884. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, U.; Nagar, R.; Shrivastava, R. Dengue and dengue haemorrhagic fever: Implications of host genetics. FEMS Immunol. Med. Microbiol. 2006, 47, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Carabali, M.; Hernandez, L.M.; Arauz, M.J.; Villar, L.A.; Ridde, V. Why are people with dengue dying? A scoping review of determinants for dengue mortality. BMC Infect. Dis. 2015, 15, 301. [Google Scholar] [CrossRef]
- Blanton, R.E.; Silva, L.K.; Morato, V.G.; Parrado, A.R.; Dias, J.P.; Melo, P.R.; Reis, E.A.; Goddard, K.A.; Nunes, M.R.; Rodrigues, S.G.; et al. Genetic ancestry and income are associated with dengue hemorrhagic fever in a highly admixed population. Eur. J. Hum. Genet. 2008, 16, 762–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Ren, S.; Zuo, Y. DC-SIGN, DC-SIGNR and LSECtin: C-type lectins for infection. Int. Rev. Immunol. 2014, 33, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, R.F.; Liu, J.W.; Lee, I.K.; Lee, C.P.; Kuo, H.C.; Huang, S.K.; Yang, K.D. DC-SIGN (CD209) promoter −336 A/G polymorphism is associated with dengue hemorrhagic fever and correlated to DC-SIGN expression and immune augmentation. PLoS Negl. Trop. Dis. 2011, 5, e934. [Google Scholar] [CrossRef]
- Mitchell, D.A.; Fadden, A.J.; Drickamer, K. A novel mechanism of carbohydrate recognition by the C-type lectins DC-SIGN and DC-SIGNR. J. Biol. Chem. 2001, 276, 28939–28945. [Google Scholar] [CrossRef]
- Boonnak, K.; Slike, B.M.; Burgess, T.H.; Mason, R.M.; Wu, S.; Sun, P.; Porter, K.; Rudiman, I.F.; Yuwono, D.; Puthavathana, P.; et al. Role of dendritic cells in antibody-dependent enhancement of dengue virus infection. J. Virol. 2008, 82, 3939–3951. [Google Scholar] [CrossRef]
- Hottz, E.D.; Oliveira, M.F.; Nunes, P.C.G.; Nogueira, R.M.R.; Valls-de-Souza, R.; Da Poian, A.T.; Weyrich, A.S.; Zimmerman, G.A.; Bozza, P.T.; Bozza, F.A. Dengue induces platelet activation, mitochondrial dysfunction and cell death through mechanisms that involve DC-SIGN and caspases. J. Thromb. Haemost. 2013, 11, 951–962. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.Y.; Sutherland, M.R.; Pryzdial, E.L.G. Dengue virus binding and replication by platelets. Blood 2015, 126, 378–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pabalan, N.; Chaisri, S.; Tabunhan, S.; Phumyen, A.; Jarjanazi, H.; Steiner, T.S. Associations of DC-SIGN (CD209) promoter-336G/A polymorphism (rs4804803) with dengue infection: A systematic review and meta-analysis. Acta Trop. 2018, 177, 186–193. [Google Scholar] [CrossRef]
- Noecker, C.A.; Amaya-Larios, I.Y.; Galeana-Hernandez, M.; Ramos-Castaneda, J.; Martinez-Vega, R.A. Contrasting associations of polymorphisms in FcγRIIa and DC-SIGN with the clinical presentation of dengue infection in a mexican population. Acta Trop. 2014, 138, 15–22. [Google Scholar] [CrossRef]
- Oliveira, L.F.; Lima, C.P.; Azevedo, R.S.; Mendonça, D.S.; Rodrigues, S.G.; Carvalho, V.L.; Pinto, E.V.; Maia, A.L.; Maia, M.H.; Vasconcelos, J.M.; et al. Polymorphism of DC-SIGN (CD209) promoter in association with clinical symptoms of dengue fever. Viral Immunol. 2014, 27, 245–249. [Google Scholar] [CrossRef]
- Alagarasu, K.; Damle, I.M.; Bachal, R.V.; Mulay, A.P.; Shah, P.S.; Dayaraj, C. Association of promoter region polymorphisms of CD209 gene with clinical outcomes of dengue virus infection in Western India. Infect. Genet. Evol. 2013, 17, 239–242. [Google Scholar] [CrossRef]
- Xavier-Carvalho, C.; Gibson, G.; Brasil, P.; Ferreira, R.X.; de Souza Santos, R.; Gonçalves Cruz, O.; de Oliveira, S.A.; de Sá Carvalho, M.; Pacheco, A.G.; Kubelka, C.F.; et al. Single nucleotide polymorphisms in candidate genes and dengue severity in children: A case–control, functional and meta-analysis study. Infect. Genet. Evol. 2013, 20, 197–205. [Google Scholar] [CrossRef]
- Silva, L.K.; Blanton, R.E.; Parrado, A.R.; Melo, P.S.; Morato, V.G.; Reis, E.A.; Dias, J.P.; Castro, J.M.; Vasconcelos, P.F.; Goddard, K.A.; et al. Dengue hemorrhagic fever is associated with polymorphisms in JAK1. Eur. J. Hum. Genet. 2010, 18, 1221–1227. [Google Scholar] [CrossRef]
- Sakuntabhai, A.; Turbpaiboon, C.; Casadémont, I.; Chuansumrit, A.; Lowhnoo, T.; Kajaste-Rudnitski, A.; Kalayanarooj, S.M.; Tangnararatchakit, K.; Tangthawornchaikul, N.; Vasanawathana, S.; et al. A variant in the CD209 promoter is associated with severity of dengue disease. Nat. Genet. 2005, 37, 507–513. [Google Scholar] [CrossRef]
- Lozach, P.; Burleigh, L.; Staropoli, I.; Navarro-Sanchez, E.; Harriague, J.; Virelizier, J.; Rey, F.A.; Desprès, P.; Arenzana-Seisdedos, F.; Amara, A. Dendritic cell-specific intercellular adhesion molecule 3-grabbing non-integrin (DC-SIGN)-mediated enhancement of dengue virus infection is independent of DC-SIGN internalization signals. J. Biol. Chem. 2005, 280, 23698–23708. [Google Scholar] [CrossRef]
- Gurevitch, J.; Koricheva, J.; Nakagawa, S.; Stewart, G. Meta-analysis and the science of research synthesis. Nature 2018, 555, 175–182. [Google Scholar] [CrossRef]
- Rodriguez, S.; Gaunt, T.R.; Day, I.N. Hardy-weinberg equilibrium testing of biological ascertainment for Mendelian randomization studies. Am. J. Epidemiol. 2009, 169, 505–514. [Google Scholar] [CrossRef]
- Wibawa, T.; Wijayanti, N.; Arguni, E.; Laksono, I.S. DC-SIGN (CD209) carbohydrate recognition domain is not polymorphic in dengue virus-infected Indonesian patients. Trop. Med. Health 2015, 43, 101–105. [Google Scholar] [CrossRef]
- Dettogni, R.S.; Tristao-Sa, R.; Dos, S.M.; Da, S.F.; Louro, I.D. Single nucleotide polymorphisms in immune system genes and their association with clinical symptoms persistence in dengue-infected persons. Hum. Immunol. 2015, 76, 717–723. [Google Scholar] [CrossRef]
- Dang, T.N.; Naka, I.; Sa-Ngasang, A.; Anantapreecha, S.; Wichukchinda, N.; Sawanpanyalert, P.; Patarapotikul, J.; Tsuchiya, N.; Ohashi, J. Association of BAK1 single nucleotide polymorphism with a risk for dengue hemorrhagic fever. BMC Med. Genet. 2016, 17, 43. [Google Scholar] [CrossRef]
- Oliveira, M.; Saraiva, D.P.; Cavadas, B.; Fernandes, V.; Pedro, N.; Casademont, I.; Koeth, F.; Alshamali, F.; Harich, N.; Cherni, L.; et al. Population genetics-informed meta-analysis in seven genes associated with risk to dengue fever disease. Infect. Genet. Evol. 2018, 62, 60–72. [Google Scholar] [CrossRef]
- Vannberg, F.O.; Chapman, S.J.; Khor, C.C.; Tosh, K.; Floyd, S.; Jackson-Sillah, D.; Crampin, A.; Sichali, L.; Bah, B.; Gustafson, P.; et al. CD209 genetic polymorphism and tuberculosis disease. PLoS ONE 2008, 3, e1388. [Google Scholar] [CrossRef]
- Chan, K.; Xu, M.; Ching, J.; So, T.; Lai, S.; Chu, C.; Yam, L.; Wong, A.; Chung, P.; Chan, V.; et al. CD209 (DC-SIGN) −336A>G promoter polymorphism and severe acute respiratory syndrome in Hong Kong Chinese. Hum. Immunol. 2010, 71, 702–707. [Google Scholar] [CrossRef]
- Selvaraj, P.; Alagarasu, K.; Swaminathan, S.; Harishankar, M.; Narendran, G. CD209 gene polymorphisms in south Indian HIV and HIV-TB patients. Infect. Genet. Evol. 2009, 9, 256–262. [Google Scholar] [CrossRef]
- Chaaithanya, I.K.; Muruganandam, N.; Surya, P.; Anwesh, M.; Alagarasu, K.; Vijayachari, P. Association of oligoadenylate synthetase gene cluster and DC-SIGN (CD209) gene polymorphisms with clinical symptoms in chikungunya virus infection. DNA Cell Biol. 2016, 35, 44–50. [Google Scholar] [CrossRef]
- Liu, P.; Ridilla, M.; Patel, P.; Betts, L.; Gallichotte, E.; Shahidi, L.; Thompson, N.L.; Jacobson, K. Beyond attachment: Roles of DC-SIGN in dengue virus infection. Traffic 2017, 18, 218–231. [Google Scholar] [CrossRef]
- Richter, M.K.; Da, S.V.J.; Torres, P.S.; Hoornweg, T.E.; van de Pol, D.P.; Rodenhuis-Zybert, I.A.; Wilschut, J.; Smit, J.M. Immature dengue virus is infectious in human immature dendritic cells via interaction with the receptor molecule DC-SIGN. PLoS ONE 2014, 9, e98785. [Google Scholar] [CrossRef] [PubMed]
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-dependent enhancement of severe dengue disease in humans. Science 2017, 358, 929–932. [Google Scholar] [CrossRef] [PubMed]
- Kuczera, D.; Assolini, J.P.; Tomiotto-Pellissier, F.; Pavanelli, W.R.; Silveira, G.F. Highlights for dengue immunopathogenesis: Antibody-dependent enhancement, cytokine storm, and beyond. J. Interferon Cytokine Res. 2018, 38, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Tomo, S.; Mohan, S.; Ramachandrappa, V.S.; Samadanam, D.M.; Suresh, S.; Pillai, A.B.; Tamilarasu, K.; Ramachandran, R.; Rajendiran, S. Dynamic modulation of DC-SIGN and FcYR2A receptors expression on platelets in dengue. PLoS ONE 2018, 13, e206346. [Google Scholar] [CrossRef] [PubMed]
- Barkhash, A.V.; Perelygin, A.A.; Babenko, V.N.; Brinton, M.A.; Voevoda, M.I. Single nucleotide polymorphism in the promoter region of the CD209 gene is associated with human predisposition to severe forms of tick-borne encephalitis. Antivir. Res. 2012, 93, 64–68. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| First Author | Publish Year | Ethnicity | Country | WHO Criteria | Matching Criteria of Controls | Method | N | Score NOS | |
|---|---|---|---|---|---|---|---|---|---|
| Severe Dengue | DF | ||||||||
| Sakuntabhai [30] | 2005 | Asian | Thailand (RA) | 1997 | Gender | TaqMan | 144 | 50 | 6 |
| Sakuntabhai [30] | 2005 | Asian | Thailand (SI) | 1997 | Gender | TaqMan | 124 | 66 | 6 |
| Sakuntabhai [30] | 2005 | Asian | Thailand (KK) | 1997 | Gender | TaqMan | 87 | 27 | 6 |
| Silva [29] | 2010 | SCA | Brazil | 1997 | Gender and age | BeadArray | 31 | 128 | 9 |
| Wang [19] | 2011 | Asian | China | 1997 | Gender | TaqMan | 106 | 157 | 6 |
| Xavier-Carvalho [28] | 2013 | SCA | Brazil | 2009 | Age | TaqMan. | 52 | 122 | 6 |
| Alagarasu [27] | 2013 | Asian | India | 1999 | Unmentioned | PCR-RFLP | 19 | 64 | 5 |
| Noecker [25] | 2014 | SCA | Mexican | 2009 | Gender | TaqMan | 33 | 51 | 6 |
| Oliveira [26] | 2014 | SCA | Brazil | 2009 | Gender and age | TaqMan | 67 | 74 | 8 |
| Dang [36] | 2016 | Asian | Thailand | 1997 | Gender | TaqMan | 415 | 331 | 6 |
| Oliveira [37] | 2018 | Asian | Vietnam | 1997 | Unmentioned | TaqMan | 20 | 21 | 6 |
| Oliveira [37] | 2018 | Asian | Cambodia | 1997 | Unmentioned | TaqMan | 93 | 57 | 6 |
| Model of inheritance | Factor | Β (95%CI) | t | p | |
|---|---|---|---|---|---|
| Dominant | GG/GA versus AA | Matching criteria of controls | 0.53 (−0.44, 1.49) | 1.23 | 0.249 |
| Ethnicity | 1.35 (0.04, 2.66) | 2.33 | 0.044 | ||
| Recessive | GG versus GA/AA | Matching criteria of controls | 0.98 (−4.41, 6.37) | 0.58 | 0.603 |
| Ethnicity | 2.20 (−3.00, 7.39) | 1.35 | 0.271 | ||
| Codominant | GG versus AA | Matching criteria of controls | 0.86 (−4.69, 6.40) | 0.49 | 0.657 |
| Ethnicity | 2.31 (−3.02, 7.65) | 1.38 | 0.262 | ||
| GA versus AA | Matching criteria of controls | 0.47 (−0.60, 1.55) | 1.02 | 0.338 | |
| Ethnicity | 1.13 (−0.39, 2.66) | 1.71 | 0.125 | ||
| Superdominant | GA versus GG/AA | Matching criteria of controls | 0.44 (−0.61, 1.50) | 0.97 | 0.359 |
| Ethnicity | 0.99 (−0.50, 2.49) | 1.53 | 0.165 | ||
| Model of inheritance | Egger’s | Begg’s | |||||
|---|---|---|---|---|---|---|---|
| t | p | z | p | ||||
| Dominant | GG/GA versus AA | 1.56 | 0.150 | 1.17 | 0.244 | ||
| Recessive | GG versus GA/AA | 0.54 | 0.616 | 1.13 | 0.260 | ||
| Codominant | GG versus AA | 0.74 | 0.499 | 0.75 | 0.452 | ||
| GA versus AA | 1.96 | 0.081 | 1.25 | 0.213 | |||
| Superdominant | GA versus GG/AA | 2.05 | 0.071 | 1.09 | 0.276 | ||
| Model of inheritance | Group | I2 (%) | Fixed effect model | Random effect model | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| z | p | OR(95%CI) | z | p | OR(95%CI) | |||||
| Dominant | GG /GA versus AA | Overall | 67.4 | 1.67 | 0.096 | 1.17 (0.97, 1.42) | 1.12 | 0.263 | 1.26 (0.84, 1.89) | |
| Asian | 72.1 | 3.12 | 0.002 | 1.46 (1.15, 1.85) | 1.98 | 0.048 | 1.86 (1.01, 3.45) | |||
| SCA | 1.8 | 1.48 | 0.140 | 0.79 (0.57, 1.08) | 1.43 | 0.153 | 0.79 (0.57, 1.09) | |||
| Recessive | GG versus GA/AA | Overall | 28.1 | 2.57 | 0.010 | 0.44 (0.23, 0.82) | 1.59 | 0.112 | 0.47 (0.19, 1.19) | |
| Asian | 47.2 | 0.40 | 0.691 | 0.83 (0.33, 2.07) | 0.21 | 0.834 | 1.22 (0.20, 7.51) | |||
| SCA | 3.1 | 2.67 | 0.008 | 0.27 (0.10, 0.70) | 2.29 | 0.022 | 0.30 (0.11, 0.84) | |||
| Codominant | GG versus AA | Overall | 33.0 | 2.63 | 0.009 | 0.43 (0.23, 0.81) | 1.54 | 0.122 | 0.46 (0.18, 1.23) | |
| Asian | 54.3 | 0.27 | 0.786 | 0.88 (0.36, 2.18) | 0.34 | 0.732 | 1.41 (0.20, 10.26) | |||
| SCA | 0.0 | 2.84 | 0.005 | 0.24 (0.09, 0.64) | 2.57 | 0.010 | 0.27 (0.10, 0.73) | |||
| GA versus AA | Overall | 63.3 | 2.46 | 0.014 | 1.28 (1.05, 1.56) | 1.62 | 0.104 | 1.38 (0.94, 2.04) | ||
| Asian | 68.1 | 3.24 | 0.001 | 1.49 (1.17, 1.90) | 2.02 | 0.043 | 1.82 (1.02, 3.26) | |||
| SCA | 28.5 | 0.37 | 0.709 | 0.94 (0.67, 1.32) | 0.32 | 0.746 | 0.94 (0.62, 1.40) | |||
| Superdominant | GA versus GG/AA | Overall | 59.9 | 2.80 | 0.005 | 1.32 (1.09, 1.61) | 1.84 | 0.065 | 1.42 (0.98, 2.06) | |
| Asian | 67.1 | 3.26 | 0.001 | 1.50 (1.17, 1.91) | 2.04 | 0.042 | 1.81 (1.02, 3.21) | |||
| SCA | 17.6 | 0.21 | 0.833 | 1.04 (0.74, 1.45) | 0.19 | 0.850 | 1.04 (0.71, 1.51) | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, J.; Wang, Z.; Chen, E. Different Associations between DC-SIGN Promoter-336G/A (rs4804803) Polymorphism with Severe Dengue in Asians and South-Central Americans: a Meta-Analysis. Int. J. Environ. Res. Public Health 2019, 16, 1475. https://doi.org/10.3390/ijerph16081475
Ren J, Wang Z, Chen E. Different Associations between DC-SIGN Promoter-336G/A (rs4804803) Polymorphism with Severe Dengue in Asians and South-Central Americans: a Meta-Analysis. International Journal of Environmental Research and Public Health. 2019; 16(8):1475. https://doi.org/10.3390/ijerph16081475
Chicago/Turabian StyleRen, Jiangping, Zhengting Wang, and Enfu Chen. 2019. "Different Associations between DC-SIGN Promoter-336G/A (rs4804803) Polymorphism with Severe Dengue in Asians and South-Central Americans: a Meta-Analysis" International Journal of Environmental Research and Public Health 16, no. 8: 1475. https://doi.org/10.3390/ijerph16081475
APA StyleRen, J., Wang, Z., & Chen, E. (2019). Different Associations between DC-SIGN Promoter-336G/A (rs4804803) Polymorphism with Severe Dengue in Asians and South-Central Americans: a Meta-Analysis. International Journal of Environmental Research and Public Health, 16(8), 1475. https://doi.org/10.3390/ijerph16081475