Mitigation of Nickel Toxicity and Growth Promotion in Sesame through the Application of a Bacterial Endophyte and Zeolite in Nickel Contaminated Soil

 ,
, 
 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Acquisition of Endophytic Bacterial Strain and Minimum Inhibitory Concentration (MIC)
2.2. Analysis of Plant Growth-Promoting (PGP) Attributes of Caulobacter sp. MN13
2.3. Experimental Design and Treatment Plan
2.4. Preparation of Inoculum
2.5. Treatment of Seeds Before Sowing
2.6. Measurement of Gas Exchange Attributes
2.7. Plant Biomass Measurement
2.8. Measurement of Physiological Attributes
2.9. Grain Biochemical and Micronutrients Analysis of Shoot
2.10. Measurement of Antioxidant Enzymes (AEs)
2.11. Persistence of Caulobacter sp. MN13 in the Rhizospheric Soil and Parts of Sesame Plant
2.12. Chemical Analysis of Metal
2.13. Statistical Analysis of Data
3. Results
3.1. MIC and PGP Characteristics
3.2. Agronomic Parameters
3.3. Gaseous Exchange Attributes
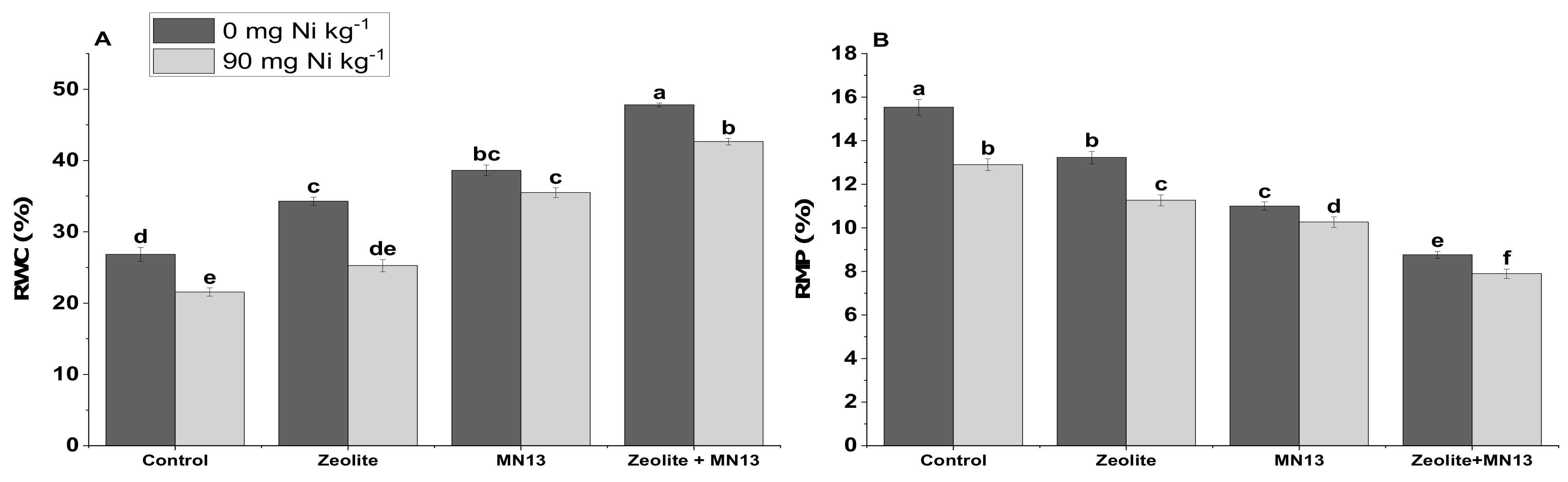
3.4. Physiological Measurements
3.5. Activities of AEs
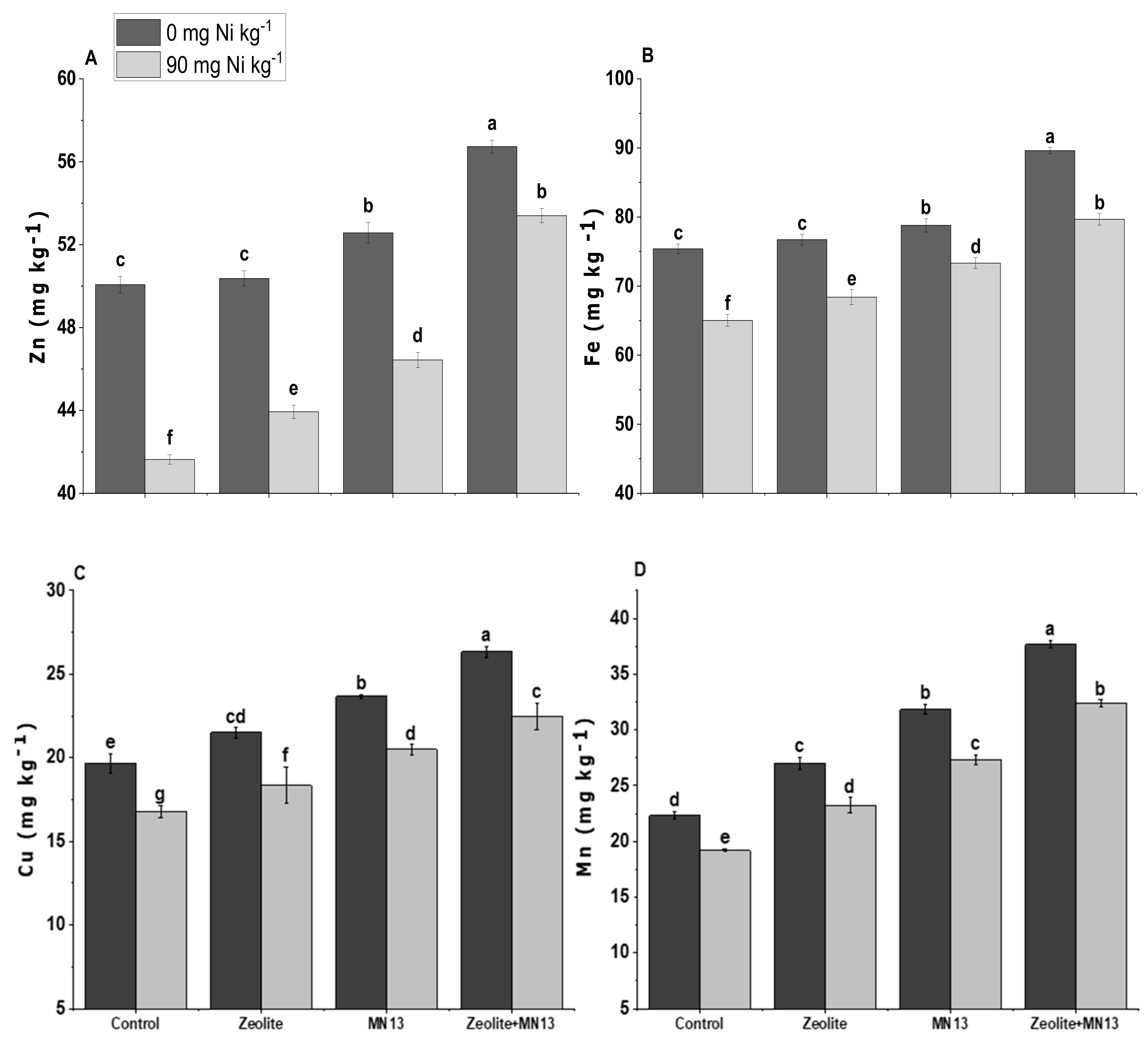
3.6. Grain Biochemical and Micronutrients Analysis of Shoot
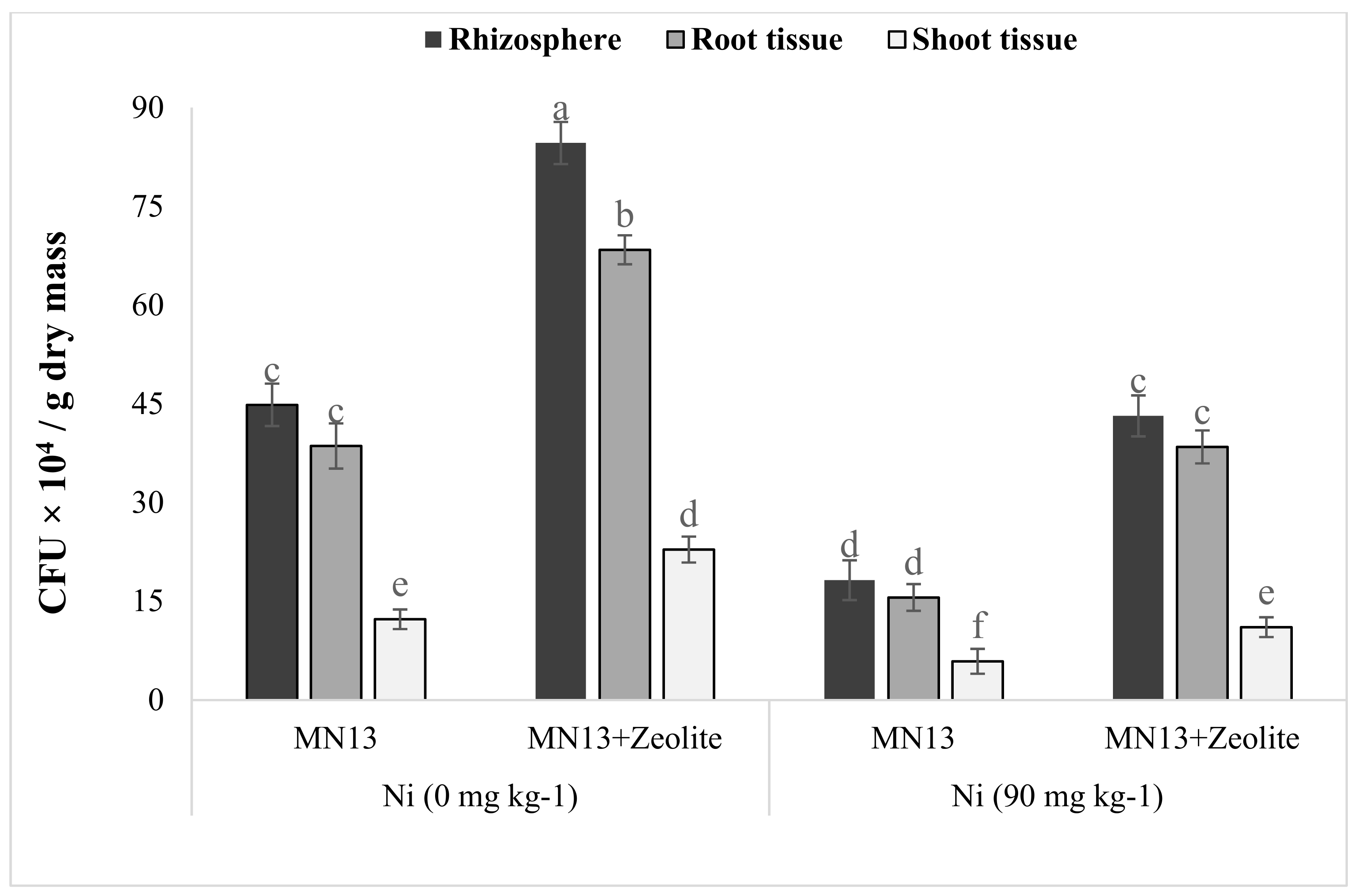
3.7. Persistence of Inoculant Strain in Rhizosphere Soil, and Sesame Root and Shoot Tissues
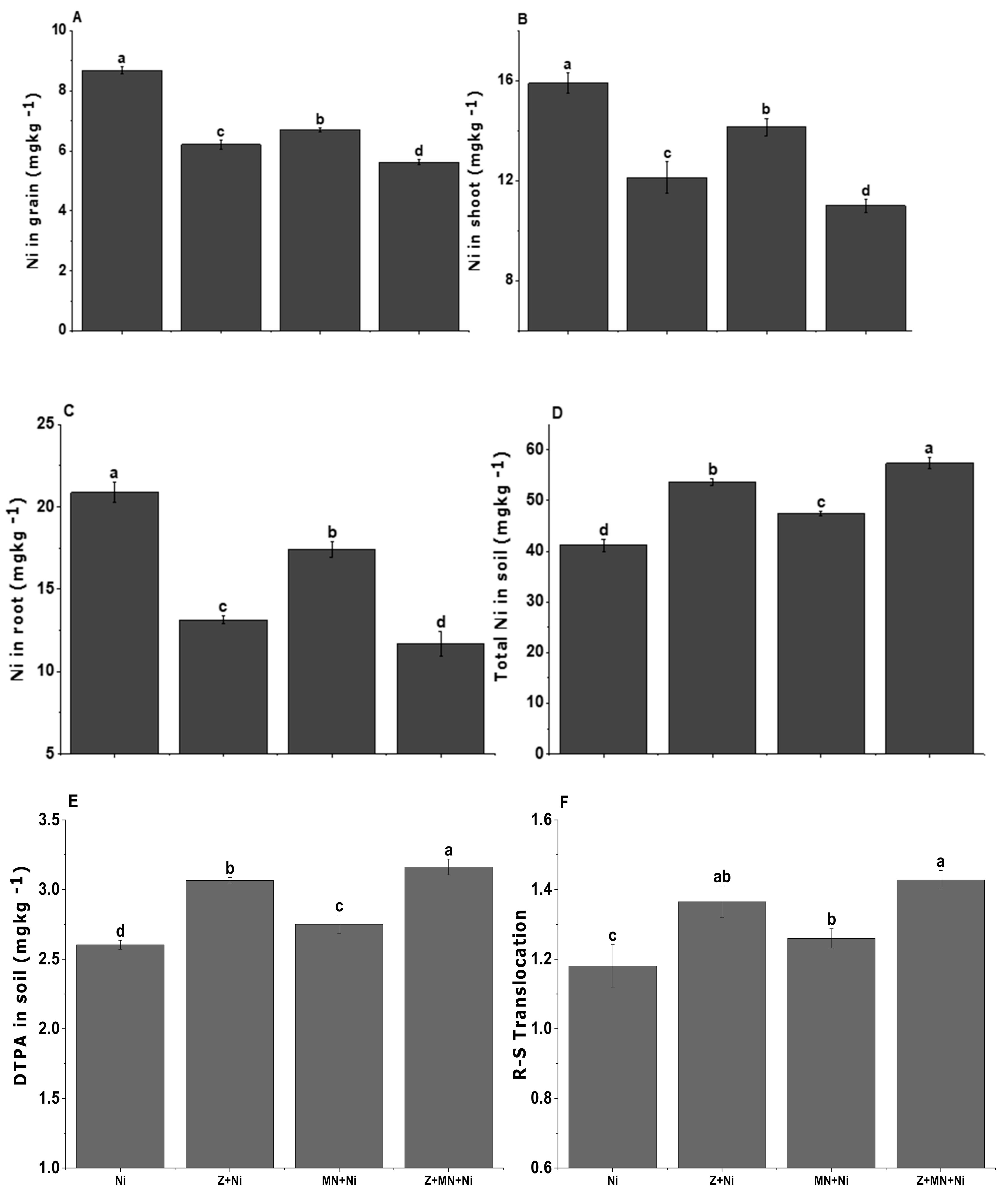
3.8. Ni Concentration in Soil, Root, Shoot, and Grain of Sesame
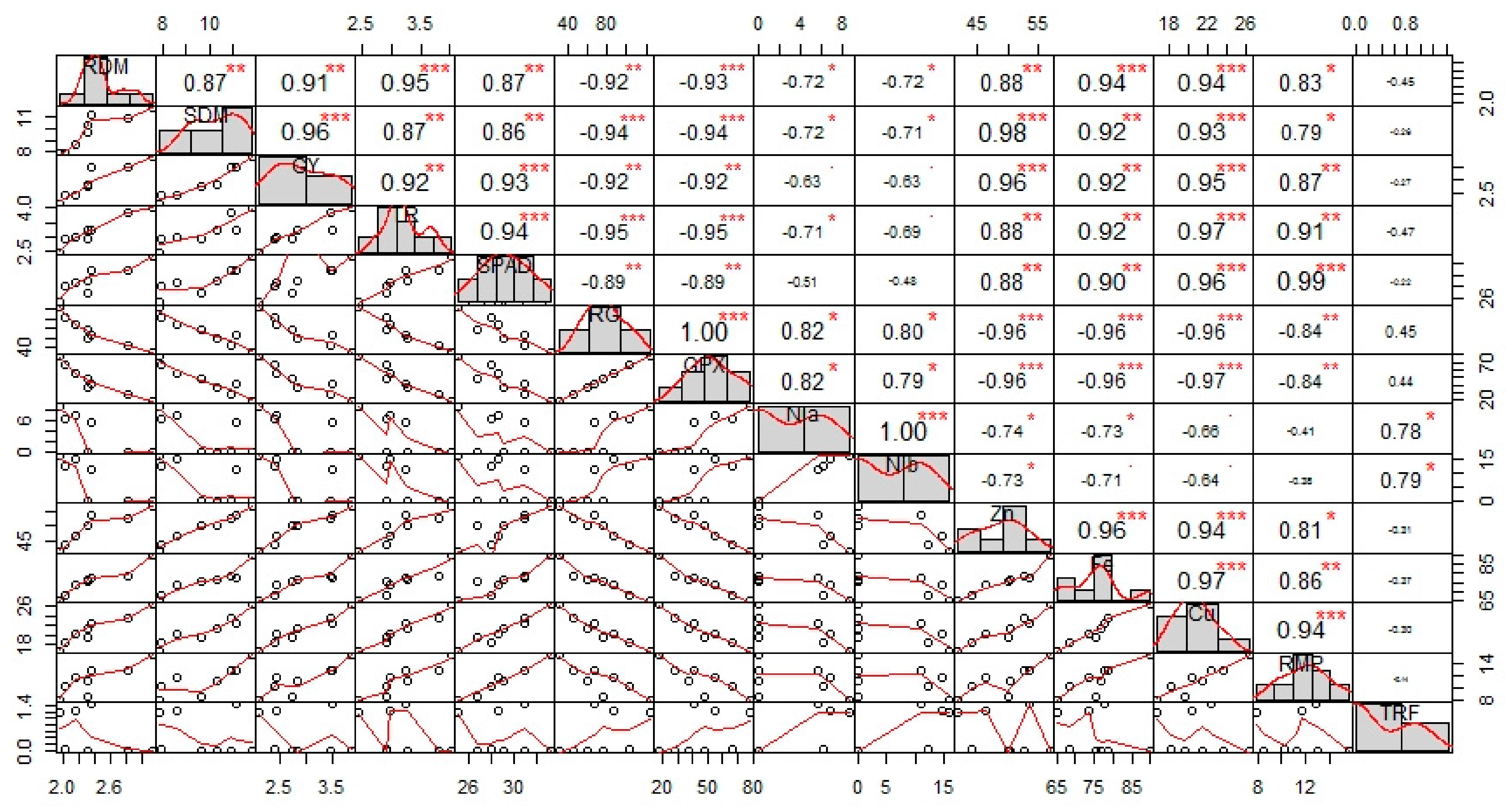
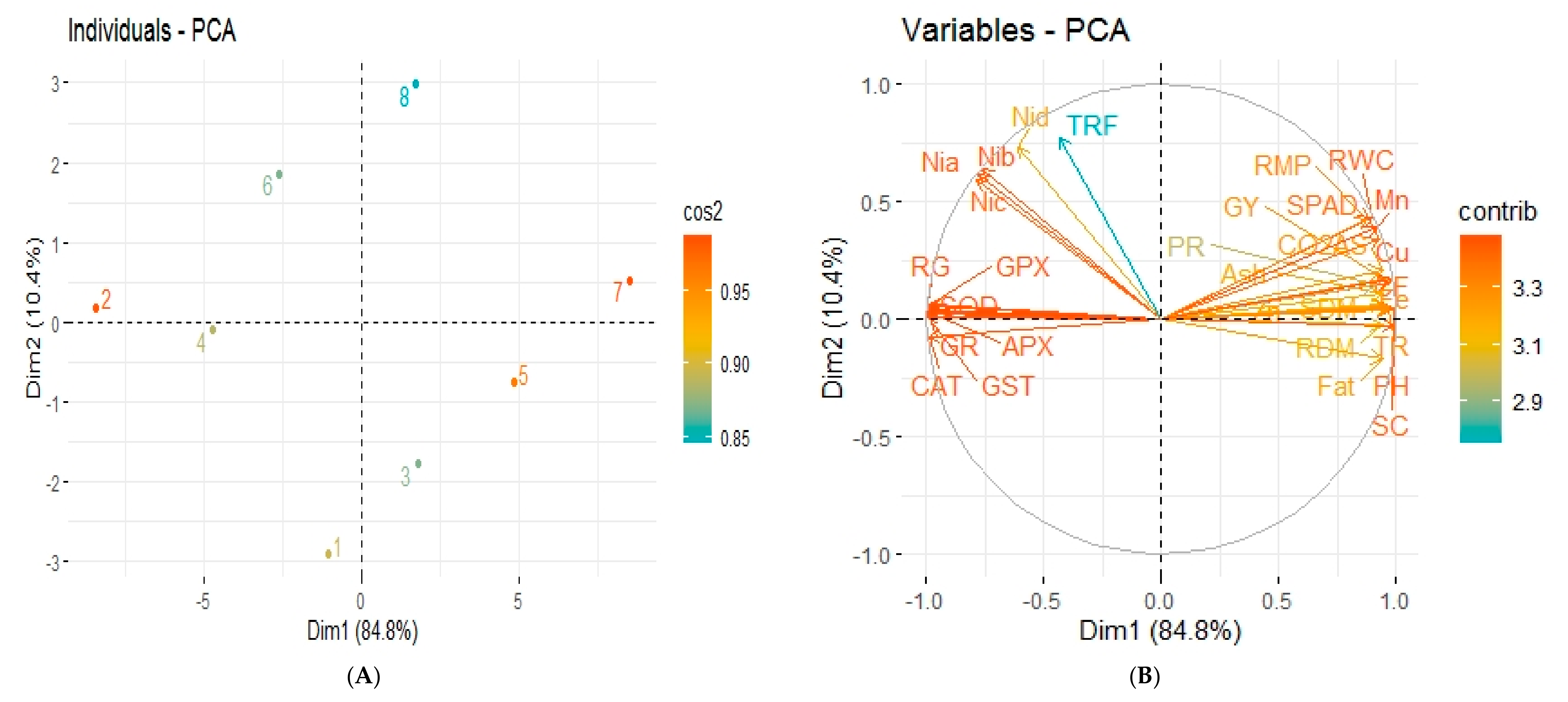
3.9. Correlation among Different Attributes of Sesame Plants and Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rizwan, M.; Imtiaz, M.; Dai, Z.; Mehmood, S.; Adeel, M.; Liu, J.; Tu, S. Nickel stressed responses of rice in Ni subcellular distribution, antioxidant production, and osmolyte accumulation. Environ. Sci. Pollut. Res. 2017, 24, 20587–20598. [Google Scholar] [CrossRef]
- Çelik, Ö.; Akdaş, E.Y. Tissue-specific transcriptional regulation of seven heavy metal stress responsive miRNAs and their putative targets in nickel indicator castor bean (R. communis L.) plants. Ecotoxicol. Environ. Saf. 2019, 170, 682–690. [Google Scholar]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef]
- Kozlow, M.V. Pollution resistance of mountain birch, Betula pubescens subsp. czerepanovii, near the copper-nickel smelter: Natural selection or phenotypic acclimation? Chemosphere 2005, 59, 189–197. [Google Scholar] [CrossRef]
- Mousavi, H.Z.; Seyedi, S.R. Nettle ash as a low cost adsorbent for the removal of nickel and cadmium from wastewater. Int. J. Environ. Sci. Technol. 2011, 8, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Harasim, P.; Filipek, T. Nickel in the environment. J. Elem. 2015, 20, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Chakankar, M.; Su, C.; Hocheng, H. Recovery of metals from solar cells by bioleaching. In Proceedings of the International Conference on Advanced Manufacturing Engineering and Technologies, Belgrade, Serbia, 5–9 June 2017; Springer: Cham, Switzerland, 2017; pp. 277–284. [Google Scholar]
- Shahbaz, A.K.; Lewińska, K.; Iqbal, J.; Ali, Q.; Iqbal, M.; Abbas, F.; Ramzani, P.M.A. Improvement in productivity, nutritional quality, and antioxidative defense mechanisms of sunflower (Helianthus annuus L.) and maize (Zea mays L.) in nickel contaminated soil amended with different biochar and zeolite ratios. J. Environ. Manag. 2018, 218, 256–270. [Google Scholar] [CrossRef]
- Ramzani, P.M.A.; Shan, L.; Anjum, S.; Ronggui, H.; Iqbal, M.; Virk, Z.A.; Kausar, S. Improved quinoa growth, physiological response, and seed nutritional quality in three soils having different stresses by the application of acidified biochar and compost. Plant Physiol. Biochem. 2017, 116, 127–138. [Google Scholar] [CrossRef]
- Parlak, K.U. Effect of nickel on growth and biochemical characteristics of wheat (Triticum aestivum L.) seedlings. NJAS Wageningen J. Life Sci. 2016, 76, 1–5. [Google Scholar] [CrossRef]
- Amjad, M.; Raza, H.; Murtaza, B.; Abbas, G.; Imran, M.; Shahid, M.; Naeem, M.A.; Zakir, A.; Iqbal, M.M. Nickel toxicity induced changes in nutrient dynamics and antioxidant profiling in two maize (Zea mays L.) Hybrids. Plants 2020, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Basalah, M.O. Interactive effect of calcium and gibberellin on nickel tolerance in relation to antioxidant systems in Triticum aestivum L. Protoplasma 2010, 248, 503–511. [Google Scholar] [CrossRef]
- Najafi, F.; Khavari-Nejad, R.A.; Hasanjanzadeh, F. The physiological responses of sunflower (Helianthus annuus L.) to NiSO4. Afr. J. Plant Sci. 2011, 5, 201–206. [Google Scholar]
- Wani, A.B.; Singh, J.; Upadhyay, N. Synthesis and characterization of transition metal complexes of para-aminosalicylic acid with evaluation of their antioxidant activities. Orient. J. Chem. 2017, 33, 1120–1126. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Kang, H.; Zhang, X.; Shao, H.; Chu, L.; Ruan, C. A critical review on the bio-removal of hazardous heavy metals from contaminated soils: Issues, progress, eco-environmental concerns and opportunities. J. Hazard. Mater. 2010, 174, 1–8. [Google Scholar] [CrossRef]
- Guo, H.; Luo, S.; Chen, L.; Xiao, X.; Xi, Q.; Wei, W.; Zeng, G.; Liu, C.; Wan, Y.; Chen, J.; et al. Bioremediation of heavy metals by growing hyperaccumulaor endophytic bacterium Bacillus sp. L14. Bioresour. Technol. 2010, 101, 8599–8605. [Google Scholar] [CrossRef]
- Saeed, Z.; Naveed, M.; Imran, M.; Bashir, M.A.; Sattar, A.; Mustafa, A.; Hussain, A.; Xu, M. Combined use of Enterobacter sp. MN17 and zeolite reverts the adverse effects of cadmium on growth, physiology and antioxidant activity of Brassica napus. PLoS ONE 2019, 14, e0213016. [Google Scholar] [CrossRef] [Green Version]
- Sabir, A.; Naveed, M.; Bashir, M.A.; Hussain, A.; Mustafa, A.; Zahir, Z.A.; Kamran, M.; Ditta, A.; Núñez-Delgado, A.; Saeed, Q.; et al. Cadmium mediated phytotoxic impacts in Brassica napus: Managing growth, physiological and oxidative disturbances through combined use of biochar and Enterobacter sp. MN17. J. Environ. Manag. 2020, 265, 110522. [Google Scholar] [CrossRef]
- Sarfraz, R.; Hussain, A.; Sabir, A.; Fekih, I.B.; Ditta, A.; Xing, S. Role of biochar and plant growth-promoting rhizobacteria to enhance soil carbon sequestration—A review. Environ. Monit. Assess. 2019, 191, 251. [Google Scholar] [CrossRef]
- Naveed, M.; Mustafa, A.; Azhar, S.Q.T.A.; Kamran, M.; Zahir, Z.A.; Núñez-Delgado, A. Burkholderia phytofirmans PsJN and tree twigs derived biochar together retrieved Pb-induced growth, physiological and biochemical disturbances by minimizing its uptake and translocation in mung bean (Vigna radiata L.). J. Environ. Manag. 2020, 257, 109974. [Google Scholar] [CrossRef]
- Sun, L.N.; Zhang, Y.F.; He, L.Y.; Chen, Z.J.; Wang, Q.Y.; Qian, M.; Sheng, X.F. Genetic diversity and characterization of heavy metal- resistant-endophytic bacteria from two copper-tolerant plant species on copper mine wasteland. Bioresour. Technol. 2010, 101, 501–509. [Google Scholar] [CrossRef]
- Ma, Y.; Oliveira, R.S.; Nai, F.; Rajkumar, M.; Luo, Y.; Rocha, I.; Freitas, H. The hyperaccumulator Sedum plumbizincicola harbors metal-resistant endophytic bacteria that improve its phytoextraction capacity in multi-metal contaminated soil. J. Environ. Manag. 2015, 156, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ge, C.; Xu, S.; Wu, Y.; Sahito, Z.A.; Ma, L.; Pan, F.; Zhou, Q.; Huang, L.; Feng, Y.; et al. The endophytic bacterium Sphingomonas SaMR12 alleviates Cd stress in oilseed rape through regulation of the GSH-AsA cycle and antioxidative enzymes. BMC Plant Biol. 2020, 20, 63. [Google Scholar] [CrossRef]
- Khan, M.A.; Khan, S.; Khan, A.; Alam, M. Soil contamination with cadmium, consequences and remediation using organic amendments. Sci. Total Environ. 2017, 601–602, 1591–1605. [Google Scholar] [CrossRef]
- Turan, V.; Khan, S.A.; Mahmood-ur-Rahman; Iqbal, M.; Ramzani, P.M.A.; Fatima, M. Promoting the productivity and quality of brinjal aligned with heavy metals immobilization in a wastewater irrigated heavy metal polluted soil with biochar and chitosan. Ecotoxicol. Environ. Saf. 2018, 161, 409–419. [Google Scholar] [CrossRef]
- Mench, M.; Renella, G.; Gelsomino, A.; Landi, L.; Nannipieri, P. Biochemical parameters and bacterial species richness in soils contaminated by sludge-borne metals and remediated with inorganic soil amendments. Environ. Pollut. 2006, 144, 24–31. [Google Scholar] [CrossRef]
- Mehmood, S.; Rizwan, M.; Bashir, S.; Ditta, A.; Aziz, O.; Yong, L.Z.; Dai, Z.; Akmal, M.; Ahmed, W.; Adeel, M.; et al. Comparative effects of biochar, slag and Ferrous–Mn ore on lead and cadmium immobilization in soil. Bull. Environ. Contam. Toxicol. 2018, 100, 286–292. [Google Scholar] [CrossRef]
- Mehmood, S.; Saeed, D.A.; Rizwan, M.; Khan, M.N.; Aziz, O.; Bashir, S.; Ibrahim, M.; Ditta, A.; Akmal, M.; Mumtaz, M.A.; et al. Impact of different amendments on biochemical responses of sesame (Sesamum indicum L.) plants grown in lead-cadmium contaminated soil. Plant Physiol. Biochem. 2018, 132, 345–355. [Google Scholar] [CrossRef]
- Mehmood, S.; Imtiaz, M.; Bashir, S.; Rizwan, M.; Irshad, S.; Yuvaraja, G.; Ikram, M.; Aziz, O.; Ditta, A.; Rehman, S.U.; et al. Leaching behaviour of Pb and Cd and transformation of their speciation in co-contaminated soil receiving different passivators. Environ. Eng. Sci. 2019, 36, 749–759. [Google Scholar] [CrossRef]
- Mierzwa-Hersztek, M.; Gondek, K.; Bajda, T.; Kope, M. Use of biochar and a zeolite as adsorbents of mineral pollution. Chem. Ind. 2019, 98, 1969–1972. [Google Scholar]
- Ali, M.A.; Naveed, M.; Mustafa, A.; Abbas, A. The good, the bad, and the ugly of rhizosphere microbiome. In Probiotics and Plant Health; Springer: Singapore, 2017; pp. 253–290. [Google Scholar]
- Boostani, H.R.; Najafi-Ghiri, M.; Hardie, A.G. Nickel immobilization in a contaminated calcareous soil with application of organic amendments and their derived biochars. Commun. Soil Sci. Plant Anal. 2020. [Google Scholar] [CrossRef]
- Radziemska, M.; Mazur, Z. Content of selected heavy metals in Ni-contaminated soil following the application of halloysite and zeolite. J. Ecol. Eng. 2016, 17, 125–133. [Google Scholar] [CrossRef]
- Lahori, A.H.; Mierzwa-Hersztek, M.; Demiraj, E.; Sajjad, R.U.; Ali, I.; Shehnaz, H.; Aziz, A.; Zuberi, M.H.; Pirzada, A.M.; Hassan, K.; et al. Direct and residual impacts of zeolite on the remediation of harmful elements in multiple contaminated soils using cabbage in rotation with corn. Chemosphere 2020, 250, 126317. [Google Scholar] [CrossRef]
- Jiménez-Castañeda, M.E.; Medina, D.I. Use of surfactant-modified zeolites and clays for the removal of heavy metals from water. Water 2017, 9, 235. [Google Scholar] [CrossRef] [Green Version]
- Peres-Caballero, R.; Gil, J.; Gondalez, J.L. The effect of adding zeolite to soils in order to improve the N-K nutrition of olive trees. Am. J. Aric. Biol. Sci. 2008, 2, 321–324. [Google Scholar]
- Prischl, M.; Hackl, E.; Pastar, M.; Pfeiffer, S.; Sessitsch, A. Genetically modified Bt maize lines containing cry3Bb1, cry1A105 or cry1Ab2 do not affect the structure and functioning of root-associated endophyte communities. Appl. Soil Ecol. 2012, 54, 39–48. [Google Scholar] [CrossRef]
- Naveed, M.; Mitter, B.; Reichenauer, T.G.; Wieczorek, K.; Sessitsch, A. Increased drought stress resilience of maize through endophytic colonization by Burkholderia phytofirmans PsJN and Enterobacter sp. FD-17. Environ. Exp. Bot. 2014, 97, 30–39. [Google Scholar] [CrossRef]
- Sarwar, M.; Ashraf, M.; Martens, A.D.; Frankenberger, W.T., Jr. Tryptophan-dependent biosynthesis of auxin in soil. Plant Soil 1992, 147, 207–215. [Google Scholar] [CrossRef]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting Rhizobacteria. Physiol. Plant 2003, 118, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.; Nautiyal, C.S. An efficient method for qualitative screening of phosphate-solubilizing bacteria. Curr. Microbiol. 2001, 43, 51–56. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Ashraf, M.; Hasnain, S.; Berge, O.; Mahmood, T. Inoculating wheat seedlings with exopolysaccharide-producing bacteria restricts sodium uptake and stimulates plant growth under salt stress. Biol. Fertil. Soils 2004, 40, 157–162. [Google Scholar] [CrossRef]
- Cappuccino, J.G.; Sherman, N. Biochemical activities of microorganisms. In Microbiology, a Laboratory Manual; The Benjamin/Cummings Publishing: San Francisco, CA, USA, 1992; pp. 125–178. [Google Scholar]
- Lorck, H. Production of hydrocyanic acid by bacteria. Physiol. Plant. 1948, 1, 142–146. [Google Scholar] [CrossRef]
- Djordjevic, D.; Wiedmann, M.; McLandsborough, L.A. Microtiter plate assay for assessment of Listeria monocytogenes biofilm formation. Appl. Environ. Microbiol. 2002, 68, 2950–2958. [Google Scholar] [CrossRef] [Green Version]
- Madi, L.; Henis, Y. Aggregation in Azospirillum brasilense Cd: Conditions and factors involved in cell-to-cell adhesion. Plant Soil 1989, 115, 89–98. [Google Scholar] [CrossRef]
- Baig, K.S.; Arshad, M.; Khalid, A.; Hussain, S.; Abbas, M.N.; Imran, M. Improving growth and yield of maize through bioinoculants carrying auxin production and phosphate solubilizing activity. Soil Environ. 2014, 33, 159–168. [Google Scholar]
- Wellburn, A.R. The spectral determination of chlorophyll-a and chlorophhyll-b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Teulat, B.; Zoumarou-Wallis, N.; Rotter, B.; Salem, M.B.; Bahri, H.; This, D. QTL for relative water content in field-grown barley and their stability across Mediterranean environments. Theor. Appl. Genet. 2003, 108, 181–188. [Google Scholar] [CrossRef]
- Yang, G.; Rhodes, D.; Joly, R.J. Effect of high temperature on membrane stability and chlorophyll fluorescence in glycinebetaine-containing maize lines. Aust. J. Plant Physiol. 1996, 23, 431–443. [Google Scholar]
- AOAC. Official Methods of Analysis of the Association of Official’s Analytical Chemists, 17th ed.; Association of official analytical chemists: Arlington, VA, USA, 2003. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for quantification of microgram quantities of protein utilizing the principle of dye binding. Anal. Chem. 1976, 72, 248–254. [Google Scholar]
- James, G.S. Analytical Chemistry of Foods; Blackie Academic and Professional: London UK, 1995; pp. 117–120. [Google Scholar]
- Jones, J.R.J.; Case, V.W. Sampling, handling, and analyzing plant tissue samples. In Soil Testing and Plant Analysis; Westerman, R.L., Ed.; Soil Science Society of America: Madison, WC, USA, 1990; pp. 389–428. [Google Scholar]
- Cakmak, I.; Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [Green Version]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposedrole in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef]
- Aebi, H.E.; Bergmeyer, H.U. Methods of Enzymatic Analysis; Verlag Chemic: Weinheim, Germany; John Wiley and Sons: Deerfield Beach, FL, USA, 1983; Volume 3, pp. 273–286. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione-S-transferase, the first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [PubMed]
- Roth, E.F.; Gilbert, H.S. The pyrogallol assay for superoxide dismutase: Absence of a glutathione artifact. Anal. Biochem. 1984, 137, 50–53. [Google Scholar] [CrossRef]
- Lindsay, W.L.; Norvell, W.A. Development of a DTPA soil test for zinc, iron, manganese and copper. Soil Sci. Soc. Am. J. 1978, 42, 421–428. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D. Principles and Procedures of Statistics: A Biometrical Approach, 3rd. ed.; McGraw Hill Book Co. Inc.: New York, NY, USA, 1997. [Google Scholar]
- Compant, S.; Mitter, B.; Colli-Mull, J.G.; Gangl, H.; Sessitsch, A. Endophytes of grapevine flowers, berries, and seeds: Identification of cultivable bacteria, comparison with other plant parts, and visualization of niches of colonization. Microbial Ecol. 2011, 62, 188–197. [Google Scholar] [CrossRef]
- Wei, Y.; Hou, H.; Xian, Y.; Guan, S.; Li, J.N.; Li, F.S. Genetic diversity of endophytic bacteria of the manganese- hyperaccumulating plant Phytolacca americana growing at a manganese mine. Eur. J. Soil Biol. 2014, 62, 15–21. [Google Scholar] [CrossRef]
- Zhu, L.J.; Guan, D.X.; Luo, J.; Rathinasabapathi, B.; Ma, L.Q. Characterization of arsenic-resistant endophytic bacteria from hyperaccumulators Pteris vittata and Pteris multifidi. Chemosphere 2014, 113, 9–16. [Google Scholar] [CrossRef]
- Malik, A. Metal bioremediation through growing cells. Environ. Inter. 2004, 30, 261–278. [Google Scholar] [CrossRef]
- Kamran, M.A.; Eqani, S.A.M.A.S.; Bibi, S.; Xu, R.K.; Monis, M.F.H.; Katsoyiannis, A.; Bokhari, H.; Chaudhary, H.J. Bioaccumulation of nickel by E. sativa and role of plant growth promoting rhizobacteria (PGPRs) under nickel stress. Ecotoxicol. Environ. Saf. 2016, 126, 256–263. [Google Scholar] [CrossRef]
- Coppola, E.; Battaglia, G.; Bucci, M.; Ceglie, D.; Colella, A.; Langella, A.; Buondonno, A.; Colella, C. Remediation of Cd-and Pb-polluted soil by treatment with organozeolite conditioner. Clays Clay Miner. 2003, 51, 609–615. [Google Scholar] [CrossRef]
- Supapron, J.; Pitayakon, L.; Touchamon, P. Effect of zeolite and chemical fertilizer on the change of physical and chemical properties on Lat Ya soil series for sugar cane. In Proceedings of the 17th WCSS, Bangkok, Thailand, 14–21 August 2002. [Google Scholar]
- Gajewska, E.; Sklodowska, M.; Slaba, M.; Mazur, J. Effect of nickel on antioxidant enzyme activities, proline and chlorophyll contents in wheat shoots. Biologia Plantarum 2006, 50, 653–659. [Google Scholar] [CrossRef]
- Seregin, I.V.; Kozhevnikova, A.D. Physiological role of nickel and its toxic effects on higher plants. Russ. J. Plant Physiol. 2006, 53, 257–277. [Google Scholar] [CrossRef]
- Kamran, M.; Malik, Z.; Parveen, A.; Zong, Y.; Abbasi, G.H.; Rafiq, M.T.; Shaaban, M.; Mustafa, A.; Bashir, S.; Rafay, M.; et al. Biochar alleviates Cd phytotoxicity by minimizing bioavailability and oxidative stress in pak choi (Brassica chinensis L.) cultivated in Cd-polluted soil. J. Environ. Manag. 2019, 250, 109500. [Google Scholar] [CrossRef]
- Gupta, A.K.; Sinha, S. Antioxidant response in sesame plants grown on industrially contaminated soil: Effect on oil yield and tolerance to lipid peroxidation. Bioresour. Technol. 2009, 100, 179–185. [Google Scholar] [CrossRef]
- Singh, K.; Pandey, N.S. Effect of nickel-stresses on uptake, pigments and antioxidative responses of water lettuce. J. Environ. Biol. 2011, 32, 391. [Google Scholar]
- Siddiqui, M.H.; Alamri, S.; Alsubaie, Q.D.; Ali, H.M.; Khan, M.N.; Al-Ghamdi, A.; Ibrahim, A.A.; Alsadon, A. Exogenous nitric oxide alleviates sulfur deficiency-induced oxidative damage in tomato seedlings. Nitric Oxide 2020, 94, 95–107. [Google Scholar] [CrossRef]
- Yandigeri, M.S.; Meena, K.K.; Singh, D.; Malviya, N.; Singh, P.D.; Solanki, K.M.; Arora, K.D. Drought-tolerant endophytic action bacteria promote growth of wheat (Triticum aestivum) under water stress conditions. Plant Growth Regul. 2012, 68, 411–420. [Google Scholar] [CrossRef]
- Zeb, H.; Hussain, A.; Naveed, M.; Ditta, A.; Ahmad, S.; Jamshaid, M.U.; Ahmad, H.T.; Hussain, B.; Aziz, R.; Haider, M.S. Compost enriched with ZnO and Zn-solubilizing bacteria improves yield and Zn-fortification in flooded rice. Ital. J. Agron. 2018, 13, 310–316. [Google Scholar] [CrossRef]
- Berrios, L.; Ely, B. Plant growth enhancement is not a conserved feature in the Caulobacter genus. Plant Soil 2020, 44, 81–95. [Google Scholar] [CrossRef]
- Hussain, A.; Zahir, Z.A.; Ditta, A.; Tahir, M.U.; Ahmad, M.; Mumtaz, M.Z.; Hayat, K.; Hussain, S. Production and implication of bio-activated organic fertilizer enriched with zinc-solubilizing bacteria to boost up maize (Zea mays L.) production and biofortification under two cropping seasons. Agronomy 2020, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Bothwell, T.H. Overview and mechanisms of iron regulation. Nutr. Rev. 1995, 53, 237–245. [Google Scholar] [CrossRef]
- Can, Ö.; Balköse, D.; Ülkü, S. Batch and column studies on heavy metal removal using a local zeolitic tuff. Desalination 2010, 259, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Turp, S.M.; Eren, B.; Ates, A. Prediction of adsorption efficiency for the removal of nickel (II) ions by zeolite using artificial neural network (ANN) approach. Fresen. Environ. Bull. 2011, 12, 3158–3165. [Google Scholar]
- Quintelas, C.; Pereira, R.; Kaplan, E.; Tavares, T. Removal of Ni (II) from aqueous solutions by an Arthrobacter viscosus biofilm supported on zeolite: From laboratory to pilot scale. Bioresour. Technol. 2013, 142, 368–374. [Google Scholar] [CrossRef] [Green Version]
- Rucinska, R.; Gwozdz, E.A. Influence of lead on membrane permeability and lipoxygenase activity in lupine roots. Biol. Plant. 2005, 49, 617–619. [Google Scholar] [CrossRef]
- Weyens, N.; Truyens, S.; Saenen, E.; Boulet, J.; Dupae, J.; van der Lelie, D. Endophytes and their potential to deal with co-contamination of organic contaminants (toluene) and toxic metals (nickel) during phytoremediation. Int. J. Phytoremediat. 2010, 13, 244–255. [Google Scholar] [CrossRef]
- Shi, Q.; Song, X.; Liu, Z.; Wang, Y.; Wang, Y.; Fu, J.; Song, Y. Quinones derived from polychlorinated biphenyls induce ROS-dependent autophagy by evoking an autophagic flux and inhibition of mTOR/p70S6k. Chem. Res. Toxicol. 2016, 29, 1160–1171. [Google Scholar] [CrossRef]
- Saleem, M.H.; Kamran, M.; Zhou, Y.; Parveen, A.; Rehman, M.; Ahmar, S.; Malik, Z.; Mustafa, A.; Anjum, R.M.A.; Wang, B.; et al. Appraising growth, oxidative stress and copper phytoextraction potential of flax (Linum usitatissimum L.) grown in soil differentially spiked with copper. J. Environ. Manag. 2020, 257, 109994. [Google Scholar] [CrossRef]
- Castaldi, P.; Santona, L.; Cozza, C.; Giuliano, V.; Abbruzzese, C.; Nastro, V.; Melis, P. Thermal and spectroscopic studies of zeolites exchanged with metal cations. J. Mol. Struct. 2005, 734, 99–105. [Google Scholar] [CrossRef]
- Akhtar, M.J.; Ullah, S.; Ahmad, I.; Rauf, A.; Nadeem, S.M.; Khan, M.Y.; Hussain, S.; Bulgariu, L. Nickel phytoextraction through bacterial inoculation in Raphanus sativus. Chemosphere 2018, 190, 234–242. [Google Scholar] [CrossRef]
- Naveed, M.; Mustafa, A.; Majeed, S.; Naseem, Z.; Saeed, Q.; Khan, A.; Nawaz, A.; Baig, K.S.; Chen, J.T. Enhancing cadmium tolerance and pea plant health through Enterobacter sp. MN17 inoculation together with biochar and gravel sand. Plants 2020, 9, 530. [Google Scholar] [CrossRef] [Green Version]
- Rajkumar, M.; Ae, N.; Freitas, H. Endophytic bacteria and their potential to enhance heavy metal phytoextraction. Chemosphere 2009, 77, 153–160. [Google Scholar] [CrossRef]
- Antoniadis, V.; Robinson, J.S.; Alloway, B.J. Effects of short-term pH fluctuations on cadmium, nickel, lead, and zinc availability to ryegrass in a sewage sludge-amended field. Chemosphere 2008, 71, 759–764. [Google Scholar] [CrossRef]
- Ditta, A.; Imtiaz, M.; Mehmood, S.; Rizwan, M.S.; Mubeen, F.; Aziz, O.; Qian, Z.; Ijaz, R.; Tu, S. Rock phosphate enriched organic fertilizer with phosphate solubilizing microorganisms improves nodulation, growth and yield of legumes. Commun. Soil Sci. Plant Anal. 2018, 49, 2715–2725. [Google Scholar] [CrossRef]
- Ditta, A.; Muhammad, J.; Imtiaz, M.; Mehmood, S.; Qian, Z.; Tu, S. Application of rock phosphate enriched composts increases nodulation, growth and yield of chickpea. Int. J. Recycl. Org. Waste Agric. 2018, 7, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, A.; Naveed, M.; Saeed, Q.; Ashraf, M.N.; Hussain, A.; Abbas, T.; Kamran, M.; Minggang, X. Application potentials of plant growth promoting rhizobacteria and fungi as an alternative to conventional weed control methods. In Crop Production; IntechOpen: London, UK, 2019. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PGP Traits | Ni (0 µg mL−1) | Ni (90 µg mL−1) |
|---|---|---|
| IAA (without L-Tryp) | 7.85 ± 0.49 | 2.64 ± 0.36 |
| IAA (With L-Tryp) | 15.25 ± 1.08 | 9.14 ± 0.98 |
| ACC deaminase activity | + | + |
| P-solubilization | ||
| Ca3(PO4)2 | − | − |
| KH2PO4 | + | + |
| FePO4 | + | + |
| AlPO4 | + | + |
| Siderophore production | + | + |
| Exopolysaccharides production | + | + |
| HCN production | − | − |
| NH4 production | + | + |
| Catalase | + | + |
| Oxidase | − | − |
| Biofilm formation | ||
| OD (600 nm) | 0.94 ± 0.04 | 0.64 ± 0.03 |
| Biofilm (595 nm) | 0.29 ± 0.02 | 0.20 ± 0.02 |
| Aggregate stability | 36.90 ± 2.52 | 25.07 ± 0.82 |
| Treatments | Ni (mg kg−1) | Plant Height (cm) | Root Dry Biomass (g plant−1) | Shoot Dry Biomass (g plant−1) | Number of Pods plant−1 | 1000-Grains Weight (g) |
|---|---|---|---|---|---|---|
| Control | 0 | 43.01 ± 1.91 c | 2.31 ± 0.162 bc | 9.67 ± 0.09 bc | 15.66 ± 1.7 cd | 2.75 ± 0.28 ab |
| 90 | 32.80 ± 1.42 de | 1.97 ± 0.087c | 7.88 ± 0.25 d | 12.33 ± 1.69 d | 2.11 ± 0.19 c | |
| Zeolite | 0 | 48.03 ± 1.68 b | 2.32 ± 0.162 bc | 10.31 ± 0.12 ab | 17.33 ± 0.5 a–d | 2.85 ± 0.21 ab |
| 90 | 37.56 ± 1.98 d | 2.03 ± 0.154 c | 7.90 ± 0.5 d | 16.0 ± 2.5 bc | 2.41 ± 0.08 bc | |
| Caulobacter sp. MN13 | 0 | 52.03 ± 1.88 ab | 2.81 ± 0.175 ab | 10.87 ± 1.02 ab | 18.66 ± 1.9 a–c | 3.47 ± 0.15 b |
| 90 | 40.54 ± 1.72 cd | 2.17 ± 0.159 c | 8.57 ± 0.73 cd | 16.96 ± 2.3 a-d | 2.44 ± 0.05 bc | |
| Zeolite + Caulobacter sp. MN13 | 0 | 55.03 ± 1.94 a | 3.13 ± 0.202 a | 11.80 ± 1.18 a | 21.66 ± 2.1 a | 3.88 ± 0.23 a |
| 90 | 49.62 ± 1.70 b | 2.85 ± 0.166 bc | 11.11 ± 1.23 ab | 19.00 ± 2.04 ab | 3.49 ± 0.09 b |
| Treatments | Ni (mg kg−1) | CO2 assimilation Rate (µmol CO2 m−2 s−1) | Transpiration Rate (µmol CO2 m−2 s−1) | Stomatal Conductance (µmol CO2 m−2 s−1) | Substomatal Conductance (µmol CO2 m−2 s−1) | Vapor Pressure Deficit (KPa) | Chlorophyll Contents (SPAD value) |
|---|---|---|---|---|---|---|---|
| Control | 0 | 9.03 ± 0.32 bc | 2.93 ± 0.95 c | 224.6 ± 5.59 b–d | 276.43 ± 4.21 bc | 2.36 ± 0.21 b–d | 26.8 ± 1.11 cd |
| 90 | 8.16 ± 0.22 c | 2.45 ± 0.66 e | 180.6 ± 4.41 d | 240.6 ± 4.78 c | 1.81 ± 0.09 e | 22.1 ± 0.75 d | |
| Zeolite | 0 | 9.6 ± 0.39 a–c | 3.26 ± 0.29 bc | 239.0 ± 6.11 a–c | 288.33 ± 5.98 ab | 2.57 ± 0.61 a–c | 29.0 ± 1.27 bc |
| 90 | 9.2 ± 0.53 bc | 2.93 ± 0.91 c | 191.3 ± 4.33 cd | 272.3 ± 5.06 bc | 1.86 ± 0.15 de | 27.9 ± 0.93 b–d | |
| Caulobacter sp. MN13 | 0 | 9.93 ± 0.33 ab | 3.82 ± 0.48 ab | 255.6 ± 5.82 ab | 299.0 ± 4.73 a | 2.94 ± 0.34 ab | 30.9 ± 0.69 ab |
| 90 | 9.5 ± 0.37 a–c | 2.98 ± 0.99 c | 204.6 ± 4.99 b–d | 276.7 ± 5.65 bc | 2.13 ± 0.19 c–e | 28.6 ± 1.10 b–d | |
| Zeolite + Caulobacter sp. MN13 | 0 | 10.3 ± 0.48 a | 4.03 ± 1.11 a | 285.0 ± 6.08 a | 330.3 ± 6.33 a | 3.04 ± 0.43 a | 33.3 ± 1.22 a |
| 90 | 9.8 ± 0.61 a–c | 3.65 ± 0.44 bc | 240.0 ± 4.37 a–c | 318.7 ± 5.87 ab | 2.55 ± 0.19 a–c | 30.9 ± 0.97 ab |
| Treatments | Ni (mg kg−1) | RG (nmol g−1 FW) | GPX (nmol min−1 mg−1) | GR (nmol min−1 mg−1) | GST (µmol min−1 mg−1) | APX (nmol min−1 mg−1) | CAT (nmol min−1 mg−1) | SOD (nmol min−1 mg−1) |
|---|---|---|---|---|---|---|---|---|
| Control | 0 | 76.0 ± 1.16 cd | 48.1 ± 0.85 cd | 19.8 ± 0.6 cd | 94.0 ± 1.27 c | 39.0 ± 1.4 cd | 13.9 ± 0.313 c | 142.7 ± 3.98 cd |
| 90 | 125.0 ± 1.19 a | 78.5 ± 1.19 a | 34.0 ± 0.71 a | 146.0 ± 1.34 a | 63.7 ± 2.03 a | 22.0 ± 0.521 a | 221.3 ± 8.31 a | |
| Zeolite | 0 | 60.7 ± 0.48 e | 37.5 ± 0.66 ef | 15.3 ± 0.41 e | 72.0 ± 1.16 d | 31.3 ± 1.31 e | 10.4 ± 0.181 de | 115.7 ± 1.41 e |
| 90 | 103.7 ± 0.98 b | 66.2 ± 1.14 b | 27.5 ± 1.04 b | 124.2 ± 1.10 b | 56.2 ± 1.27 b | 17.7 ± 0.495 b | 190.7 ± 2.41 b | |
| Caulobacter sp. MN13 | 0 | 46.0 ± 0.41 f | 28.3 ± 0.57 f | 10.7 ± 0.09 f | 54.5 ± 0.48 e | 22.3 ± 0.94 f | 7.4 ± 0.211 e | 88.3 ± 1.12 f |
| 90 | 86.3 ± 1.19 c | 54.5 ± 0.82 c | 22.3 ± 1.01 c | 101.6 ± 0.87 c | 45.3 ± 1.52 c | 13.3 ± 0.322 cd | 157.7 ± 3.33 c | |
| Zeolite + Caulobacter sp. MN13 | 0 | 30.7 ± 0.15 g | 17.3 ± 0.16 g | 5.8 ± 0.09 g | 34.5 ± 0.21 f | 13.5 ± 0.06 g | 3.6 ± 0.021 f | 56.7 ± 1.63 g |
| 90 | 67.0 ± 0.38 de | 41.5 ± 0.71 de | 16.2 ± 0.4 de | 75.4 ± 1.17 d | 34.7 ± 0.5 de | 10.1 ± 0.175 e | 124.0 ± 3.74 de |
| Treatments | Ni (mg kg−1) | Ash (%) | Crude Fibre (%) | Protein (%) | Fats (%) |
|---|---|---|---|---|---|
| Control | 0 | 3.81 ± 0.28 bc | 10.29 ± 0.37 a–c | 27.28 ± 1.11 bc | 14.40 ± 0.31 bc |
| 90 | 2.91 ± 0.19 d | 9.23 ± 0.34 c | 25.11 ± 0.84 e | 11.91 ± 0.17 c | |
| Zeolite | 0 | 3.90 ± 0.27 bc | 10.77 ± 0.39 a–c | 29.88 ± 1.15 ab | 16.30 ± 0.44 ab |
| 90 | 3.71 ± 0.25 c | 9.85 ± 0.26 bc | 25.99 ± 0.85 de | 13.87 ± 0.36 bc | |
| Caulobacter sp. MN13 | 0 | 4.42 ± 0.41 ab | 11.89 ± 0.31 ab | 29.11 ± 1.14 ab | 16.81 ± 0.15 ab |
| 90 | 3.91 ± 0.27 bc | 10.17 ± 0.29 bc | 26.14 ± 1.03 cd | 13.94 ± 0.18 bc | |
| Zeolite + Caulobacter sp. MN13 | 0 | 4.63 ± 0.43 a | 12.23 ± 0.41 a | 31.99 ± 1.11 a | 18.91 ± 0.12 a |
| 90 | 4.11 ± 0.4 a–c | 11.49 ± 0.34 ab | 30.88 ± 0.94 ab | 14.11 ± 0.23 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naveed, M.; Bukhari, S.S.; Mustafa, A.; Ditta, A.; Alamri, S.; El-Esawi, M.A.; Rafique, M.; Ashraf, S.; Siddiqui, M.H. Mitigation of Nickel Toxicity and Growth Promotion in Sesame through the Application of a Bacterial Endophyte and Zeolite in Nickel Contaminated Soil. Int. J. Environ. Res. Public Health 2020, 17, 8859. https://doi.org/10.3390/ijerph17238859
Naveed M, Bukhari SS, Mustafa A, Ditta A, Alamri S, El-Esawi MA, Rafique M, Ashraf S, Siddiqui MH. Mitigation of Nickel Toxicity and Growth Promotion in Sesame through the Application of a Bacterial Endophyte and Zeolite in Nickel Contaminated Soil. International Journal of Environmental Research and Public Health. 2020; 17(23):8859. https://doi.org/10.3390/ijerph17238859
Chicago/Turabian StyleNaveed, Muhammad, Syeda Sosan Bukhari, Adnan Mustafa, Allah Ditta, Saud Alamri, Mohamed A. El-Esawi, Munazza Rafique, Sobia Ashraf, and Manzer H. Siddiqui. 2020. "Mitigation of Nickel Toxicity and Growth Promotion in Sesame through the Application of a Bacterial Endophyte and Zeolite in Nickel Contaminated Soil" International Journal of Environmental Research and Public Health 17, no. 23: 8859. https://doi.org/10.3390/ijerph17238859
APA StyleNaveed, M., Bukhari, S. S., Mustafa, A., Ditta, A., Alamri, S., El-Esawi, M. A., Rafique, M., Ashraf, S., & Siddiqui, M. H. (2020). Mitigation of Nickel Toxicity and Growth Promotion in Sesame through the Application of a Bacterial Endophyte and Zeolite in Nickel Contaminated Soil. International Journal of Environmental Research and Public Health, 17(23), 8859. https://doi.org/10.3390/ijerph17238859