Nickel: Human Health and Environmental Toxicology

 , ,
, , 
 and
and
Abstract
:1. Introduction
2. Chemical Form, Properties and Sources of Nickel Compounds
3. Nickel Toxicity and Carcinogenicity
4. Nickel Allergy
5. Epigenetic Effects of Nickel
6. Teratogenicity of Nickel Compounds
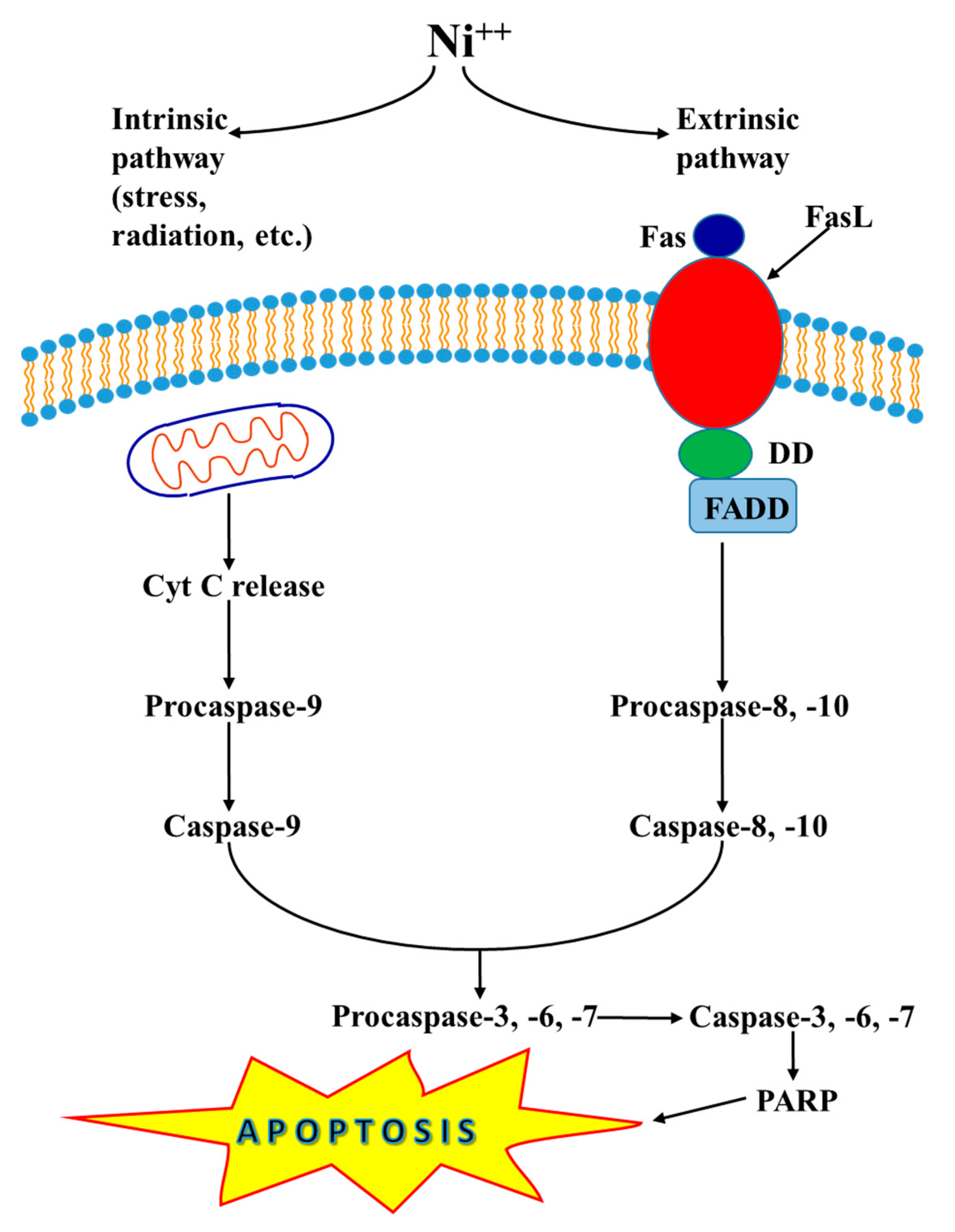
7. Nickel-Induced Apoptosis
8. Nickel Phytoremediation and Phytomining
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DNA | deoxyribonucleic acid; |
| RNA | ribonucleic acid; |
| AMP | adenosine 5′-monophosphate; |
| ATP | adenosine 5′-triphosphate; |
| ROS | reactive oxygen species; |
| SOD | superoxide dismutase; |
| CAT | catalase; |
| MDA | malondialdehyde; |
| NO | nitric oxide; |
| mRNA | messenger RNA; |
| Cyt C | Cytochrome C; |
| BAX | BCL2 Associated X; |
| BID | BH3 interacting domain death agonist; |
| Bcl-2 | B-cell lymphoma-2; |
| SAM | S-adenosyl methionine; |
| Fhit | fragile histidine triad protein; |
| Fas | first apoptosis signal; |
| FasL | Fas Ligant; |
| NAC | N-acetylcysteine; |
| TEMPO | 2,2,6,6-tetramethyl-1-piperidinyloxy. |
References
- Muñoz, A.; Costa, M. Elucidating the mechanisms of nickel compound uptake: A review of particulate and nano-nickel endocytosis and toxicity. Toxicol. Appl. Pharm. 2012, 260, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Kenston, S.S.F.; Kong, L.; Zhao, J. Molecular mechanisms of nickel induced neurotoxicity and chemoprevention. Toxicology 2017, 392, 47–54. [Google Scholar] [CrossRef]
- Ragsdale, S.W. Nickel-based enzyme systems. J. Biol. Chem. 2009, 284, 18571–18575. [Google Scholar] [CrossRef] [Green Version]
- Boer, J.L.; Mulrooney, S.B.; Hausinger, R.P. Nickel-dependent metalloenzymes. Arch. Biochem. Biophys. 2014, 544, 142–152. [Google Scholar] [CrossRef] [Green Version]
- Maroney, M.J.; Ciurli, S. Nonredox nickel enzymes. Chem. Rev. 2014, 114, 4206–4228. [Google Scholar] [CrossRef] [Green Version]
- Desguin, B.; Fellner, M.; Riant, O.; Hu, J.; Hausinger, R.P.; Hols, P.; Soumillion, P. Biosynthesis of the nickel-pincer nucleotide cofactor of lactate racemase requires a CTP-dependent cyclometallase. J. Biol. Chem. 2018, 293, 12303–12317. [Google Scholar] [CrossRef] [Green Version]
- Sawers, R.G. Nickel in Bacteria and Archaea. In Encyclopedia of Metalloproteins; Kretsinger, R.H., Uversky, V.N., Permyakov, E.A., Eds.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Sreekanth, T.V.M.; Nagajyothi, P.C.; Lee, K.D.; Prasad, T.N.V.K.V. Occurrence, physiological responses and toxicity of nickel in plants. Int. J. Environ. Sci. Technol. 2013, 10, 1129–1140. [Google Scholar] [CrossRef] [Green Version]
- Macomber, L.; Hausinger, R.P. Mechanisms of nickel toxicity in microorganisms. Metallomics 2011, 3, 1153–1162. [Google Scholar] [CrossRef] [Green Version]
- Can, M.; Armstrong, F.A.; Ragsdale, S.W. Structure, function, and mechanism of the nickel metalloenzymes, CO dehydrogenase, and acetyl-CoA synthase. Chem. Rev. 2014, 114, 4149–4174. [Google Scholar] [CrossRef]
- Guldan, H.; Sterner, R.; Babinger, P. Identification and characterization of a bacterial glycerol-1-phosphate dehydrogenase: Ni(2+)-dependent AraM from Bacillus subtilis. Biochemistry 2008, 47, 376–384. [Google Scholar] [CrossRef]
- Merkens, H.; Kappl, R.; Jakob, R.P.; Schmid, F.X.; Fetzner, S. Quercetinase QueD of Streptomyces sp. FLA, a monocupin dioxygenase with a preference for nickel and cobalt. Biochemistry 2008, 47, 12185–12196. [Google Scholar] [CrossRef]
- Ragsdale, S.W. Nickel and the carbon cycle. J. Inorg. Biochem. 2007, 101, 1657–1666. [Google Scholar] [CrossRef]
- Chen, Q.Y.; Brocato, J.; Laulicht, F.; Costa, M. Mechanisms of nickel carcinogenesis. In Essential and Non-Essential metals. Molecular and Integrative Toxicology; Mudipalli, A., Zelikoff, J.T., Eds.; Springer International Publishing AG: New York, NY, USA, 2017; pp. 181–197. [Google Scholar]
- Sinicropi, M.S.; Caruso, A.; Capasso, A.; Palladino, C.; Panno, A.; Saturnino, C. Heavy metals: Toxicity and carcinogenicity. Pharmacologyonline 2010, 2, 329–333. [Google Scholar]
- Sinicropi, M.S.; Amantea, D.; Caruso, A.; Saturnino, C. Chemical and biological properties of toxic metals and use of chelating agents for the pharmacological treatment of metal poisoning. Arch. Toxicol. 2010, 84, 501–520. [Google Scholar] [CrossRef]
- Sinicropi, M.S.; Rovito, N.; Carocci, A.; Genchi, G. Acetyl-L-carnitine in Parkinson’s disease. In Mechanisms in Parkinson’s Disease—Models and Treatments; Dushanova, J., Ed.; InTech: London, UK, 2012; Chapter 19; pp. 367–392. ISBN 978-953-307-876-2. [Google Scholar]
- Amado, A.; Jacob, S.E. Contact dermatitis to foods. Actas Dermo-Sifiliográficas 2007, 98, 452–458. [Google Scholar] [CrossRef]
- Carocci, A.; Sinicropi, M.S.; Catalano, A.; Lauria, G.; Genchi, G. Melatonin in Parkinson’s disease. In A Synopsis of Parkinson’s Disease; InTech: London, UK, 2014; Chapter 3; pp. 71–99. ISBN 978-953-51-1229-7. [Google Scholar]
- Xu, S.; He, M.; Zhong, M.; Li, L.; Lu, Y.; Zhang, Y.; Zhang, L.; Yu, Z.; Zhou, Z. The neuroprotective effects of taurine against nickel by reducing oxidative stress and maintaining mitochondrial function in cortical neurons. Neurosci. Lett. 2015, 590, 52–57. [Google Scholar] [CrossRef]
- Schaumlöffel, D. Nickel species: Analysis and toxic effects. J. Trace Elem. Med. Biol. 2012, 26, 1–6. [Google Scholar] [CrossRef]
- Denkhaus, E.; Salnikow, K. Nickel essentiality, toxicity, and carcinogenicity. Crit. Rev. Oncol. Hematol. 2002, 42, 35–56. [Google Scholar] [CrossRef]
- Cempel, M.; Nikel, G. Nickel: A review of its sources and environmental toxicology. Pol. J. Environ. Stud. 2006, 15, 375–382. [Google Scholar]
- Henderson, R.G.; Durando, J.; Oller, A.R.; Merkel, D.J.; Marone, P.A.; Bates, H.K. Acute oral toxicity of nickel compounds. Regul. Toxicol. Pharm. 2012, 62, 425–432. [Google Scholar] [CrossRef]
- Beattie, H.; Keen, C.; Coldwell, M.; Tan, E.; Morton, J.; McAlinden, J.; Smith, P. The use of bio-monitoring to assess exposure in the electroplating industry. J. Expo. Sci. Environ. Epidemiol. 2017, 27, 47–55. [Google Scholar] [CrossRef] [Green Version]
- International Agency for Research on Cancer (IARC). IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. In Nickel and Nickel Compounds Monograph; WHO Press: Geneva, Switzerland, 2017; pp. 169–218. [Google Scholar]
- Von Burg, R. Toxicology update. J. Appl. Toxicol. 1997, 17, 425–431. [Google Scholar] [CrossRef]
- Carocci, A.; Catalano, A.; Lauria, G.; Sinicropi, M.S.; Genchi, G. A review on mercury toxicity in food. In Food Toxicology; Debasis, B., Anand, S., Stohs, S.J., Eds.; CRC Press: Boca Raton, FL, USA, 2016; Chapter 16; pp. 315–326. [Google Scholar]
- Lavinia, B.; Florina, R.; Augustin, C. Is it possible a nickel-free diet? Acta Medica. Marisiensis. 2018, 64, 5. [Google Scholar]
- Vasiluk, L.; Sowa, J.; Sanborn, P.; Ford, F.; Dutton, M.D.; Hale, B. Bioaccessibility estimates by gastric SBRC method to determine relationships to bioavailability of nickel in ultramafic soils. Sci. Total Environ. 2019, 673, 685–693. [Google Scholar] [CrossRef]
- Kong, L.; Gao, X.; Zhu, J.; Cheng, K.; Tang, M. Mechanisms involved in reproductive toxicity caused by nickel nanoparticle in female rats. Environ. Toxicol. 2016, 31, 1674–1683. [Google Scholar] [CrossRef]
- Sharma, A.; Hickman, J.; Gazit, N.; Rabkin, E.; Mishin, Y. Nickel nanoparticles set a new record of strength. Nat. Commun. 2018, 9, 4102. [Google Scholar] [CrossRef]
- Future Markets, Inc. The Global Market for Metal and Metal Oxide Nanoparticles, 2010–2027, 4th ed.; Future Markets, Inc.: Edinburgh, UK, 2017. [Google Scholar]
- Zhao, J.; Shi, X.; Castranova, V.; Ding, M. Occupational toxicology of nickel and nickel compounds. J. Environ. Pathol. Toxicol. Oncol. 2009, 28, 177–208. [Google Scholar] [CrossRef]
- Zambelli, B.; Ciurli, S. Nickel and human health. Met. Ions Life Sci. 2013, 13, 321–357. [Google Scholar]
- McGregor, D.B.; Baan, R.A.; Partensky, C.; Rice, J.M.; Wilbourn, J.D. Evaluation of the carcinogenic risks to humans associated with surgical implants and other foreign bodies—A report of an IARC Monographs Programme Meeting. International Agency for Research on Cancer. Eur. J. Cancer 2000, 36, 307–313. [Google Scholar] [CrossRef]
- Seilkop, S.K.; Oller, A.R. Respiratory cancer risks associated with low-level nickel exposure: An integrated assessment based on animal, epidemiological, and mechanistic data. Regul. Toxicol. Pharm. 2003, 37, 173–190. [Google Scholar] [CrossRef]
- Sivulka, D.J. Assessment of respiratory carcinogenicity associated with exposure to metallic nickel: A review. Regul. Toxicol. Pharm. 2005, 43, 117–133. [Google Scholar] [CrossRef]
- Chiou, Y.H.; Wong, R.H.; Chao, M.R.; Chen, C.Y.; Liou, S.H.; Lee, H. Nickel accumulation in lung tissues is associated with increased risk of p53 mutation in lung cancer patients. Environ. Mol. Mutagen. 2014, 55, 624–632. [Google Scholar] [CrossRef]
- Zambelli, B.; Uversky, V.N.; Ciurli, S. Nickel impact on human health: An intrinsic disorder perspective. BBA Proteins Proteom. 2016, 1864, 1714–1731. [Google Scholar] [CrossRef]
- Sivulka, D.J.; Seilkop, S.K.; Lascelles, K.; Conard, B.R.; Jones, S.F.; Collinson, E.C. Reconstruction of historical exposures at a Welsh nickel refinery (1953–2000). Ann. Occup. Hyg. 2014, 58, 739–760. [Google Scholar] [CrossRef]
- Jose, C.C.; Jagannathan, L.; Tanwar, V.S.; Zhang, X.; Zang, C.; Cuddapah, S. Nickel exposure induces persistent mesenchymal phenotype in human lung epithelial cells through epigenetic activation of ZEB1. Mol. Carcinog. 2018, 57, 794–806. [Google Scholar] [CrossRef]
- Sutunkova, M.P.; Privalova, L.I.; Minigalieva, I.A.; Gurvich, V.B.; Panov, V.G.; Katsnelson, B.A. The most important inferences from the Ekaterinburg nanotoxicology team’s animal experiments assessing adverse health effects of metallic and metal oxide nanoparticles. Toxicol. Rep. 2018, 5, 363–376. [Google Scholar] [CrossRef]
- Horie, M.; Stowe, M.; Tabei, M.; Kuroda, E. Metal ion release of manufactured metal oxide nanoparticles is involved in the allergic response to inhaled ovalbumin in mice. Occup. Dis. Environ. 2016, 4, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Latvala, S.; Hedberg, J.; Di Bucchianico, S.; Möller, L.; Odnevall Wallinder, I.; Elihn, K.; Karlsson, H.L. Nickel release, ROS generation and toxicity of Ni and NiO micro- and nanoparticles. PLoS ONE 2016, 11, e0159684. [Google Scholar] [CrossRef]
- IARC (The International Agency for Research on Cancer). Nickel and nickel compounds. IARC Monogr. Eval. Carcinog. Risk Hum. 2012, 100C, 169–218. [Google Scholar]
- Kumar, S.; Trivedi, A.V. A review on role of nickel in the biological system. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 719–727. [Google Scholar] [CrossRef]
- Sutherland, J.E.; Costa, M. Nickel. In Heavy Metals in the Environment; Sarkar, B., Ed.; Marcel Decker Inc.: New York, NY, USA, 2002. [Google Scholar]
- Goodman, J.E.; Prueitt, R.L.; Thakali, S.; Oller, A.R. The nickel ion bioavailability model of the carcinogenic potential of nickel-containing substances in the lung. Crit. Rev. Toxicol. 2011, 41, 142–174. [Google Scholar] [CrossRef]
- Buxton, S.; Garman, E.; Heim, K.E.; Lyons-Darden, T.; Schlekat, C.E.; Taylor, M.D.; Oller, A.R. Concise review of nickel human health toxicology and ecotoxicology. Inorganics 2019, 7, 89. [Google Scholar] [CrossRef] [Green Version]
- Kawanishi, S.; Inoue, S.; Oikawa, S.; Yamashita, N.; Toyokuni, S.; Kawanishi, M.; Nishino, K. Oxidative DNA damage in cultured cells and rat lungs by carcinogenic nickel compounds. Free Radic. Biol. Med. 2001, 31, 108–116. [Google Scholar] [CrossRef]
- Chen, H.; Costa, M. Iron- and 2-oxoglutarate-dependent dioxygenases: An emerging group of molecular targets for nickel toxicity and carcinogenicity. Biometals 2009, 22, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Giri, N.C.; Zhang, R.; Yamane, K.; Zhang, Y.; Maroney, M.; Costa, M. Nickel ions inhibit histone demethylase JMJD1A and DNA repair enzyme ABH2 by replacing the ferrous iron in the catalytic centers. J. Biol. Chem. 2010, 285, 7374–7383. [Google Scholar] [CrossRef] [Green Version]
- Niemirowicz, G.; Parussini, F.; Aguero, F.; Cazzulo, J.J. Two metallocarboxypeptidases from the protozoan Trypanosoma cruzi belong to the M32 family, found so far only in prokaryotes. Biochem. J. 2007, 401, 399–410. [Google Scholar] [CrossRef]
- Buan, N.R.; Suh, S.J.; Escalante-Semerena, J.C. The eutT gene of Salmonella enterica encodes an oxygen-labile, metal-containing ATP:corrinoid adenosyltransferase enzyme. J. Bacteriol. 2004, 186, 5708–5714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, C.L.; Buszko, M.L.; Bobik, T.A. Purification and initial characterization of the Salmonella enterica PduO ATP:Cob(I)alamin adenosyltransferase. J. Bacteriol. 2004, 186, 7881–7887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buan, N.R.; Escalante-Semerena, J.C. Purification and initial biochemical characterization of ATP:Cob(I)alamin adenosyltransferase (EutT) enzyme of Salmonella Enterica. J. Biol. Chem. 2006, 281, 16971–16977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.C.; Hsu, W.H.; Chien, F.T.; Chen, C.Y. Crystal structure and site-directed mutagenesis studies of N-carbamoyl-D-amino-acid amidohydrolase from Agrobacterium radiobacter reveals a homotetramer and insight into a catalytic cleft. J. Mol. Biol. 2001, 306, 251–261. [Google Scholar] [CrossRef]
- Schaeffer, M.; Miranda, A.; Mottram, J.C.; Coombs, G.H. Differentiation of Leishmania major is impaired by over-expression of pyroglutamyl peptidase I. Mol. Biochem. Parasitol. 2006, 150, 318–329. [Google Scholar] [CrossRef]
- Carocci, A.; Rovito, N.; Sinicropi, M.S.; Genchi, G. Mercury toxicity and neurodegenerative effects. Rev. Environ. Contam. Toxicol. 2014, 229, 1–18. [Google Scholar]
- Carocci, A.; Catalano, A.; Lauria, G.; Sinicropi, M.S.; Genchi, G. Lead Toxicity, antioxidant defense and environment. Rev. Environ. Contam. Toxicol. 2016, 238, 45–67. [Google Scholar]
- Genchi, G.; Sinicropi, M.S.; Carocci, A.; Lauria, G.; Catalano, A. Mercury exposure and heart diseases. Int. J. Environ. Res. Public Health 2017, 14, 74. [Google Scholar] [CrossRef] [Green Version]
- Faccioni, F.; Franceschetti, P.; Cerpelloni, M.; Fracasso, M.E. In vivo study on metal release from fixed orthodontic appliances and DNA damage in oral mucosa cells. Am. J. Orthod. Dentofac. Orthop. 2003, 124, 687–693. [Google Scholar] [CrossRef]
- Lü, X.; Bao, X.; Huang, Y.; Qu, Y.; Lu, H.; Lu, Z. Mechanisms of cytotoxicity of nickel ions based on gene expression profiles. Biomaterials 2009, 30, 141–148. [Google Scholar] [CrossRef]
- Hafez, H.S.; Selim, E.M.; Kamel Eid, F.H.; Tawfik, W.A.; Al-Ashkar, E.A.; Mostafa, Y.A. Cytotoxicity, genotoxicity, and metal release in patients with fixed orthodontic appliances: A longitudinal in-vivo study. Am. J. Orthod. Dentofac. Orthop. 2011, 140, 298–308. [Google Scholar] [CrossRef]
- Saito, M.; Arakaki, R.; Yamada, A.; Tsunematsu, T.; Kudo, Y.; Ishimaru, N. Molecular mechanisms of nickel allergy. Int. J. Mol. Sci. 2016, 17, 202. [Google Scholar] [CrossRef] [Green Version]
- Gollhausen, R.; Ring, J. Allergy to coined money: Nickel contact dermatitis in cashiers. J. Am. Acad Derm. 1991, 25, 365–369. [Google Scholar] [CrossRef]
- Shum, K.W.; Meyer, J.D.; Chen, Y.; Cherry, N.; Gawkrodger, D.J. Occupational contact dermatitis to nickel: Experience of the British dermatologists (EPIDERM) and occupational physicians (OPRA) surveillance schemes. Occup. Environ. Med. 2003, 60, 954–957. [Google Scholar] [CrossRef] [Green Version]
- Nesle, F.O.; Spiedel, H.; Spiedel, M.O. High nickel release from 1- and 2-euro coins. Metallurgy 2002, 419, 132. [Google Scholar]
- Lachapelle, J.M.; Marot, L. High nickel release from 1- and 2-euro coins: Are there practical implications? Dermatology 2004, 209, 288–290. [Google Scholar] [CrossRef]
- Rothman, S. Überempfindlichkeit gegen Hartgelt. Derm. Wochenschr. 1930, 90, 98. [Google Scholar]
- Preininger, T. Überempfindlichkeit gegen Nickelgeld. Derm. Wochenschr. 1934, 99, 1082. [Google Scholar]
- McAlester, A.W., Jr.; McAlester, A.W. Nickel sensitization from white gold spectacle frames. A J. Ophthalmol. 1931, 14, 925–926. [Google Scholar] [CrossRef]
- Du Bois, C.H. Contribution a la dermite des parures. Am. Derm. Syph. 1932, 7, 209–215. [Google Scholar]
- Vahter, M.; Berglund, M.; Åkesson, A.; Lidén, C. Metals and women’s health. Environ. Res. 2002, 88, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Alinaghi, F.; Bennike, N.H.; Egeberg, A.; Thyssen, J.P.; Johansen, J.D. Prevalence of contact allergy in the general population: A systematic review and meta-analysis. Contact Dermat. 2019, 80, 77–85. [Google Scholar] [CrossRef]
- Noble, J.; Ahing, S.I.; Karaiskos, N.E.; Wiltshire, J. Nickel allergy and orthodontics, a review and report of two cases. Br. Dent. J. 2008, 204, 297–300. [Google Scholar] [CrossRef]
- Cioffi, M.; Gilliland, D.; Ceccone, G.; Chiesa, R.; Cigada, A. Electrochemical release testing of nickel-titanium orthodontic wires in artificial saliva using thin layer activation. Acta Biomater. 2005, 1, 717–724. [Google Scholar] [CrossRef]
- Vemula, P.K.; Anderson, R.R.; Karp, J.M. Nanoparticles reduce nickel allergy by capturing metal ions. Nat. Nanotechnol. 2011, 6, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Hill, H.; Goldenberger, A.; Sheehan, M.P.; Patel, A.; Jacob, S.E. Nickel-Free alternatives raise awareness. Dermatitis 2015, 26, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Tsang, A.C.O.; Nicholson, P.; Pereira, V.M. Nickel-related adverse reactions in the treatment of cerebral aneurysms: A literature review. World Neurosurg. 2018, 115, 147–153. [Google Scholar] [CrossRef]
- Honari, G.; Ellis, S.G.; Wilkoff, B.L.; Aronica, M.A.; Svensson, L.G.; Taylor, J.S. Hypersensitivity reactions associated with endovascular devices. Contact Dermat. 2008, 59, 7–22. [Google Scholar] [CrossRef]
- Da Mata Perez, L.; França, A.T.; Zimmerman, J.R. Systemic nickel allergy syndrome. World Allergy Organ. J. 2015, 8, A89. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, M.V.C.; Bourc’his, D. The diverse roles of DNA methylation in mammalian development and disease. Nat. Rev. Mol. Cell Biol. 2019, 20, 590–607. [Google Scholar] [CrossRef]
- Baccarelli, A.; Bollati, V. Epigenetics and environmental chemicals. Curr. Opin. Pediatr. 2009, 21, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Denhardt, D.T. Effect of stress on human biology: Epigenetics, adaptation, inheritance, and social significance. J. Cell Physiol. 2018, 233, 1975–1984. [Google Scholar] [CrossRef]
- Koturbash, I.; Beland, F.A.; Pogribny, I.P. Role of epigenetic events in chemical carcinogenesis—a justification for incorporating epigenetic evaluations in cancer risk assessment. Toxicol. Mech. Methods 2011, 21, 289–297. [Google Scholar] [CrossRef]
- Sun, H.; Shamy, M.; Costa, M. Nickel and epigenetic gene silencing. Genes 2013, 4, 583–595. [Google Scholar] [CrossRef]
- Grewal, S.I.; Moazed, D. Heterochromatin and epigenetic control of gene expression. Science 2003, 301, 798–802. [Google Scholar] [CrossRef] [Green Version]
- Sadakierska-Chudy, A.; Małgorzata, F.A. Comprehensive view of the epigenetic landscape. Part II: Histone post-translational modification, nucleosome level, and chromatin regulation by ncRNAs. Neurotox. Res. 2015, 27, 172–197. [Google Scholar] [CrossRef] [Green Version]
- Pillai, R.S.; Bhattacharyya, S.N.; Filipowicz, W. Repression of protein synthesis by miRNAs: How many mechanisms? Trends Cell Biol. 2007, 17, 118–126. [Google Scholar] [CrossRef]
- Wang, Z.; Yao, H.; Lin, S.; Zhu, X.; Shen, Z.; Lu, G.; Poon, W.S.; Xie, D.; Lin, M.C.; Kung, H.F. Transcriptional and epigenetic regulation of human microRNAs. Cancer Lett. 2013, 331, 1–10. [Google Scholar] [CrossRef]
- Costa, M.; Davidson, T.L.; Chen, H.; Ke, Q.; Zhang, P.; Yan, Y.; Huang, C.; Kluz, T. Nickel carcinogenesis: Epigenetics and hypoxia signaling. Mutat. Res. 2005, 592, 79–88. [Google Scholar] [CrossRef]
- Ke, Q.; Li, Q.; Ellen, T.P.; Sun, H.; Costa, M. Nickel compounds induce phosphorylation of histone H3 at serine 10 by activating JNK–MAPK pathway. Carcinogenesis 2008, 29, 1276–1281. [Google Scholar] [CrossRef] [Green Version]
- Cantone, L.; Nordio, F.; Hou, L.; Apostoli, P.; Bonzini, M.; Tarantini, L.; Angelici, L.; Bollati, V.; Zanobetti, A.; Schwartz, J.; et al. Inhalable metal-rich air particles and histone H3K4 dimethylation and H3K9 acetylation in a cross-sectional study of steel workers. Environ. Health Perspect. 2011, 119, 964–969. [Google Scholar] [CrossRef] [Green Version]
- Ke, Q.; Ellen, T.P.; Costa, M. Nickel compounds induce histone ubiquitination by inhibiting histone deubiquitinating enzyme activity. Toxicol. Appl. Pharm. 2008, 228, 190–199. [Google Scholar] [CrossRef] [Green Version]
- Arita, A.; Costa, M. Epigenetics in metal carcinogenesis: Nickel, arsenic, chromium and cadmium. Metallomics 2009, 1, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Ellen, T.P.; Kluz, T.; Harder, M.E.; Xiong, J.; Costa, M. Heterochromatinization as a potential mechanism of nickel-induced carcinogenesis. Biochemistry 2009, 48, 4626–4632. [Google Scholar] [CrossRef] [Green Version]
- Salnikow, K.; Zhitkovich, A. Genetic and epigenetic mechanisms in metal carcinogenesis and cocarcinogenesis: Nickel, arsenic, and chromium. Chem. Res. Toxicol. 2008, 21, 28–44. [Google Scholar] [CrossRef] [Green Version]
- Govindarajan, B.; Klafter, R.; Miller, M.S.; Mansur, C.; Mizesko, M.; Bai, X.; LaMontagne, K., Jr.; Arbiser, J.L. Reactive oxygen-induced carcinogenesis causes hypermethylation of p16(Ink4a) and activation of MAP kinase. Mol. Med. 2002, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kowara, R.; Salnikow, K.; Diwan, B.A.; Bare, R.M.; Waalkes, M.P.; Kasprzak, K.S. Reduced Fhit protein expression in nickel-transformed mouse cells and in nickel-induced murine sarcomas. Mol. Cell Biochem. 2004, 255, 195–202. [Google Scholar] [CrossRef]
- Chen, H.; Ke, Q.; Kluz, T.; Yan, Y.; Costa, M. Nickel ions increase histone H3 lysine 9 dimethylation and induce transgene silencing. Mol. Cell Biol. 2006, 26, 3728–3737. [Google Scholar] [CrossRef] [Green Version]
- Ke, Q.; Davidson, T.; Chen, H.; Kluz, T.; Costa, M. Alterations of histone modifications and transgene silencing by nickel chloride. Carcinogenesis 2006, 27, 1481–1488. [Google Scholar] [CrossRef] [Green Version]
- Cheng, T.F.; Choudhuri, S.; Muldoon-Jacobs, K. Epigenetic targets of some toxicologically relevant metals: A review of the literature. J. Appl. Toxicol. 2012, 32, 643–653. [Google Scholar] [CrossRef]
- Karaczyn, A.A.; Bal, W.; North, S.L.; Bare, R.M.; Hoang, V.M.; Fisher, R.J.; Kasprzak, K.S. The octapeptidic end of the C-terminal tail of histone H2A is cleaved off in cells exposed to carcinogenic nickel (II). Chem. Res. Toxicol. 2003, 16, 1555–1559. [Google Scholar] [CrossRef]
- Sunderman, F.W., Jr.; Shen, S.K.; Reid, M.C.; Allpass, P.R. Teratogenicity and embryotoxicity of nickel carbonyl in Syrian hamsters. Teratog. Carcinog. Mutagen. 1980, 1, 223–233. [Google Scholar] [CrossRef]
- Sunderman, F.W., Jr.; Reid, M.C.; Shen, S.K.; Kevorkian, C.B. Embryotoxicity and teratogenicity of nickel compounds. In Reproductive and Developmental Toxicity of Metals; Clarkson, T.W., Nordberg, G.F., Sager, P.R., Eds.; Springer: Boston, MA, USA, 1983; pp. 399–416. [Google Scholar]
- Léonard, A.; Gerber, G.B.; Jacquet, P. Carcinogenicity, mutagenicity and teratogenicity of nickel. Mutat. Res. Rev. Genet. Toxicol. 1981, 87, 1–15. [Google Scholar] [CrossRef]
- Leonard, A.; Jacquet, P. Embryotoxicity and Genotoxicity of Nickel; Iarc Scientific Publications: Lyon, France, 1984; pp. 277–291. [Google Scholar]
- Saini, S.; Nair, N.; Saini, M.R. Embryotoxic and teratogenic effects of nickel in Swiss albino mice during organogenetic period. BioMed Res. Int. 2013. [Google Scholar] [CrossRef] [Green Version]
- Hengartner, M.O. The biochemistry of apoptosis. Nature 2000, 407, 770–776. [Google Scholar] [CrossRef]
- D’Arcy, M.S. Cell death: A review of the major form of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, X.; Gueydan, C.; Han, J. Plasma membrane changes during programmed cell deaths. Cell Res 2018, 28, 9–21. [Google Scholar] [CrossRef]
- Guo, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Wang, X.; Zhao, L.; Wu, B.; Chen, K.; Deng, J. Nickel chloride-induced apoptosis via mitochondria- and Fas-mediated caspase-dependent pathways in broiler chickens. Oncotarget 2016, 7, 79747–79760. [Google Scholar] [CrossRef] [Green Version]
- Su, L.; Deng, Y.; Zhang, Y.; Li, C.; Zhang, R.; Sun, Y.; Zhang, K.; Li, J.; Yao, S. Protective effects of grape seed procyanidin extract against nickel sulfate-induced apoptosis and oxidative stress in rat testes. Toxicol. Mech. Methods 2011, 21, 487–494. [Google Scholar] [CrossRef]
- Zou, L.; Su, L.; Sun, Y.; Han, A.; Chang, X.; Zhu, A.; Liu, F.; Li, J.; Sun, Y. Nickel sulfate induced apoptosis via activating ROS-dependent mitochondria and endoplasmic reticulum stress pathways in rat Leydig cells. Environ. Toxicol. 2017, 32, 1918–1926. [Google Scholar] [CrossRef]
- Kang, J.; Zhang, D.; Chen, J.; Lin, C.; Liu, Q. Involvement of histone hypoacetylation in Ni2+-induced Bcl-2 down-regulation and human hepatoma cell apoptosis. J. Biol. Inorg. Chem. 2004, 9, 713–723. [Google Scholar] [CrossRef]
- Cavani, A. Breaking tolerance to nickel. Toxicology 2005, 209, 119–121. [Google Scholar] [CrossRef]
- Guan, F.; Zhang, D.; Wang, X.; Chen, J. Nitric oxide and Bcl-2 mediated the apoptosis induced by nickel(II) in human T hybridoma cells. Toxicol. Appl. Pharm. 2007, 221, 86–94. [Google Scholar] [CrossRef]
- Siddiqui, M.A.; Ahamed, M.; Ahmad, J.; Majeed Khan, M.A.; Musarrat, J.; Al-Khedhairy, A.A.; Alrokayan, S.A. Nickel oxide nanoparticles induce cytotoxicity, oxidative stress and apoptosis in cultured human cells that is abrogated by the dietary antioxidant curcumin. Food Chem. Toxicol. 2012, 50, 641–647. [Google Scholar] [CrossRef]
- Ahamed, M.; Ali, D.; Alhadlaq, H.A.; Akhtar, M.J. Nickel oxide nanoparticles exert cytotoxicity via oxidative stress and induce apoptotic response in human liver cells (HepG2). Chemosphere 2013, 93, 514–522. [Google Scholar] [CrossRef]
- Freitas, M.; Barcellos-de-Souza, P.; Barja-Fidalg, C.; Fernandes, E. Nickel induces apoptosis in human neutrophils. Biometals 2013, 26, 13–21. [Google Scholar] [CrossRef]
- Ma, C.; Song, M.; Zhang, Y.; Yan, M.; Zhang, M.; Bi, H. Nickel nanowires induce cell cycle arrest and apoptosis by generation of reactive oxygen species in HeLa cells. Toxicol. Rep. 2014, 1, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.L.; Xin, R.; Wang, S.Y.; Wang, Y.; Zhang, L.; Yu, C.P.; Wu, Y.H. Nickel-smelting fumes induce mitochondrial damage and apoptosis, accompanied by decreases in viability, in NIH/3T3 cells. Arch. Biochem. Biophys. 2018, 660, 20–28. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals-concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Rafati, M.; Khorasani, N.; Moattar, F.; Shirvany, A.; Moraghebi, F.; Hosseinzadeh, S. Phytoremediation potential of Populus alba and Morus alba for cadmium, chromium and nickel absorption from polluted soil. Int. J. Environ. Res. 2011, 5, 961–970. [Google Scholar]
- Mukhopadhyay, S.; Maiti, S.K. Phytoremediation of metal enriched mine waste: A review. Glob. J. Environ. Res. 2010, 4, 135–150. [Google Scholar]
- Singh, S. Phytoremediation: A sustainable alternative for environmental challenges. Int J. Gr. Herb. Che. 2012, 1, 133–139. [Google Scholar]
- Vishnoi, S.R.; Srivastava, P.N. Phytoremediation-green for environmental clean. In Proceedings of the 12th World Lake Conference, Jaipur, India, 28 October–2 November 2007. [Google Scholar]
- Sakai, Y.; Ma, Y.; Xu, C.; Wu, H.; Zhu, W.; Yang, J. Phytodesalination of a salt-affected soil with four halophytes in China. J. Arid Land Stud. 2012, 22, 17–20. [Google Scholar]
- Jiassi, A.; Zorrig, W.; El Khouni, A.; Lakhdar, A.; Smaoui, A.; Abdelly, C.; Rabhi, M. Phytodesalination of a moderately-salt-affected soil by Sulla carnosa. Int. J. Phytoremediat. 2013, 15, 398–404. [Google Scholar]
- Chaney, R.L.; Angle, J.S.; Baker, A.J.M.; Li, Y.M. Method for Phytomining of Nickel, Cobalt, and other Metals from Soil. US patent No 6786948, 7 September 2004. [Google Scholar]
- Minguzzi, C.; Vergnano, O. Il contenuto di nichel nelle ceneri di Alyssum Bertolonii. Atti. Soc. Tosc. Sci. Nat. 1948, 55, 49–74. [Google Scholar]
- Jaffré, T.; Brooks, R.R.; Lee, J.; Reeves, R.D. Sebertia acuminata: A hyperaccumulator of nickel from New Caledonia. Science 1976, 193, 579–580. [Google Scholar] [CrossRef] [Green Version]
- Jaffré, T.; Pillon, Y.; Thomine, S.; Merlot, S. The metal hyperaccumulators from New Caledonia can broaden our understanding of nickel accumulation in plants. Front. Plant. Sci. 2013, 4, 279. [Google Scholar] [CrossRef] [Green Version]
- Jaffrè, T.; Reeves, R.D.; Baker, A.J.M.; Schat, H.; van der Ent, A. The discovery of nickel hyperaccumulation in the New Caledonian tree Pycnandra acuminata 40 years on: An introduction to a Virtual Issue. New Phytol. 2018, 218, 397–400. [Google Scholar] [CrossRef] [Green Version]
- Callahan, D.L.; Roessner, U.; Dumontet, V.; De Livera, A.M.; Doronila, A.; Baker, A.J.; Kolev, S.D. Elemental and metabolite profiling of nickel hyperaccumulators from New Caledonia. Phytochemistry 2012, 81, 80–89. [Google Scholar] [CrossRef]
- Giordani, C.; Cecchi, S.; Zanchi, C. Phytoremediation of soil polluted by nickel using agricultural crops. Environ. Manag. 2005, 36, 675–681. [Google Scholar] [CrossRef]
- Tappero, R.; Peltier, E.; Grafe, M.; Heidel, K.; Ginder-Vogel, M.; Livi, K.J.; Rivers, M.L.; Marcus, M.A.; Chaney, R.L.; Sparks, D.L. Hyperaccumulator Alyssum murale relies on a different metal storage mechanism for cobalt than for nickel. New Phytol. 2007, 175, 641–654. [Google Scholar] [CrossRef] [Green Version]
- Broadhurst, C.L.; Chaney, R.L. Growth and metal accumulation of an Alyssum murale nickel hyperaccumulator ecotype co-cropped with Alyssum montanum and perennial ryegrass in Serpentine Soil. Front. Plant. Sci. 2016, 7, 451. [Google Scholar] [CrossRef] [Green Version]
- Grison, C.; Escande, V.; Petit, E.; Garoux, L.; Boulanger, C.; Grison, C. Psychotria douarrei and Geissois pruinosa, novel resources for the plant-based catalytic chemistry. RSC. Adv. 2013, 44, 22340–22345. [Google Scholar] [CrossRef]
- Fernando, E.S.; Quimado, M.O.; Doronila, A.I. Rinorea niccolifera (Violaceae), a new, nickel-hyperaccumulating species from Luzon Island, Philippines. Phytokeys 2014, 37, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Fernando, E.S.; Quimado, M.O.; Trinidad, L.C.; Doronila, A.I. The potential use of indigenous nickel hyperaccumulators for small-scale mining. Philipp. J. Degrad. Min. Lands Manag. 2013, 1, 21–26. [Google Scholar]
- Roccotiello, E.; Serrano, H.C.; Mariotti, M.G.; Branquinho, C. Nickel phytoremediation potential of the Mediterranean Alyssoides utriculata (L.) Medik. Chemosphere 2015, 119, 1372–1378. [Google Scholar] [CrossRef]
- Roccotiello, E.; Serrano, H.C.; Mariotti, M.G.; Branquinho, C. The impact of Ni on the physiology of a Mediterranean Ni-hyperaccumulating plant. Environ. Sci. Pollut. Res. Int. 2016, 23, 12414–12422. [Google Scholar] [CrossRef]
- Van der Ent, A.; Callahan, D.L.; Noller, B.N.; Mesjasz-Przybylowicz, J.; Przybylowicz, W.J.; Barnabas, A.; Harris, H.H. Nickel biopathways in tropical nickel hyperaccumulating trees from Sabah (Malaysia). Sci. Rep. 2017, 7, 41861. [Google Scholar] [CrossRef]
- Malik, A. Metal bioremediation through growing cells. Environ. Int. 2004, 30, 261–278. [Google Scholar] [CrossRef]
- Akram Husain, R.S.; Thatheyus, A.J.; Ramya, D. Bioremoval of nickel using Pseudomonas Fluoresc. Am. J. Microbiol. Res. 2013, 1, 48–52. [Google Scholar]
- Zhu, X.; Li, W.; Zhan, L.; Huang, M.; Zhang, Q.; Achal, V. The large-scale process of microbial carbonate precipitation for nickel. Environmental Pollution remediation from an industrial soil. Environ. Pollut. 2016, 219, 149–155. [Google Scholar] [CrossRef]
- Massoud, R.; Hadiani, M.R.; Hamzehlou, P.; Khosravi-Darani, K. Bioremediation of heavy metals in food industry: Application of Saccharomyces cerevisiae. Electron. J. Biotechnol. 2019, 37, 56–60. [Google Scholar] [CrossRef]
- Siddiquee, S.; Rovina, K.; Al Azad, S.; Naher, L.; Suryani, S.; Chaikaew, P. Heavy metal contaminants removal from wastewater using the potential filamentous fungi biomass: A review. J. Microb. Biochem. Technol. 2015, 7, 384–393. [Google Scholar] [CrossRef]
- Abdel-Monem, M.O.; AL-Zubeiry, A.H.S.; AL-Gheethi, A.A.S. Biosorption of nickel by Pseudomonas cepacia 120S and Bacillus subtilis 117S. Water Sci. Technol. 2010, 61, 2994–3007. [Google Scholar] [CrossRef]


{kind=link}
| Nickel Containing Foods | Hazelnuts; cocoa and dark chocolate; fruits (almonds, dates, figs, pineapple, plums, raspberries); grains (bran, buckwheat, millet, whole grain bread, oats, brown rice, sesame seeds, sunflower seeds); seafood (shrimps, mussels, oysters, crab, salmon); vegetables (beans, savoy cabbage, leeks, lettuce, lentils, peas, spinach, cabbage), tea from drinks dispensers; soya and soya products; peanuts; licorice; baking powder. |
| Nickel Containing Items | Inexpensive jewelry; cosmetics; keys; cell phones; eyeglass frames; paper clips; orthodontic braces; stainless steel articles; nickel plated articles; clothing fasteners (zippers, snap buttons, belt buckles); electrical equipment; armaments; alloy; metallurgical and food processing industries; pigments; catalysts. |
| Nickel Toxic Effects | Contact dermatitis; headaches; gastrointestinal manifestations; respiratory manifestations; lung fibrosis; cardiovascular diseases; lung cancer; nasal cancer; epigenetic effects. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genchi, G.; Carocci, A.; Lauria, G.; Sinicropi, M.S.; Catalano, A. Nickel: Human Health and Environmental Toxicology. Int. J. Environ. Res. Public Health 2020, 17, 679. https://doi.org/10.3390/ijerph17030679
Genchi G, Carocci A, Lauria G, Sinicropi MS, Catalano A. Nickel: Human Health and Environmental Toxicology. International Journal of Environmental Research and Public Health. 2020; 17(3):679. https://doi.org/10.3390/ijerph17030679
Chicago/Turabian StyleGenchi, Giuseppe, Alessia Carocci, Graziantonio Lauria, Maria Stefania Sinicropi, and Alessia Catalano. 2020. "Nickel: Human Health and Environmental Toxicology" International Journal of Environmental Research and Public Health 17, no. 3: 679. https://doi.org/10.3390/ijerph17030679
APA StyleGenchi, G., Carocci, A., Lauria, G., Sinicropi, M. S., & Catalano, A. (2020). Nickel: Human Health and Environmental Toxicology. International Journal of Environmental Research and Public Health, 17(3), 679. https://doi.org/10.3390/ijerph17030679