Consumption of a Branched-Chain Amino Acid (BCAA) during Days 2–10 of Pregnancy Causes Abnormal Fetal and Placental Growth: Implications for BCAA Supplementation in Humans



Abstract
:1. Introduction
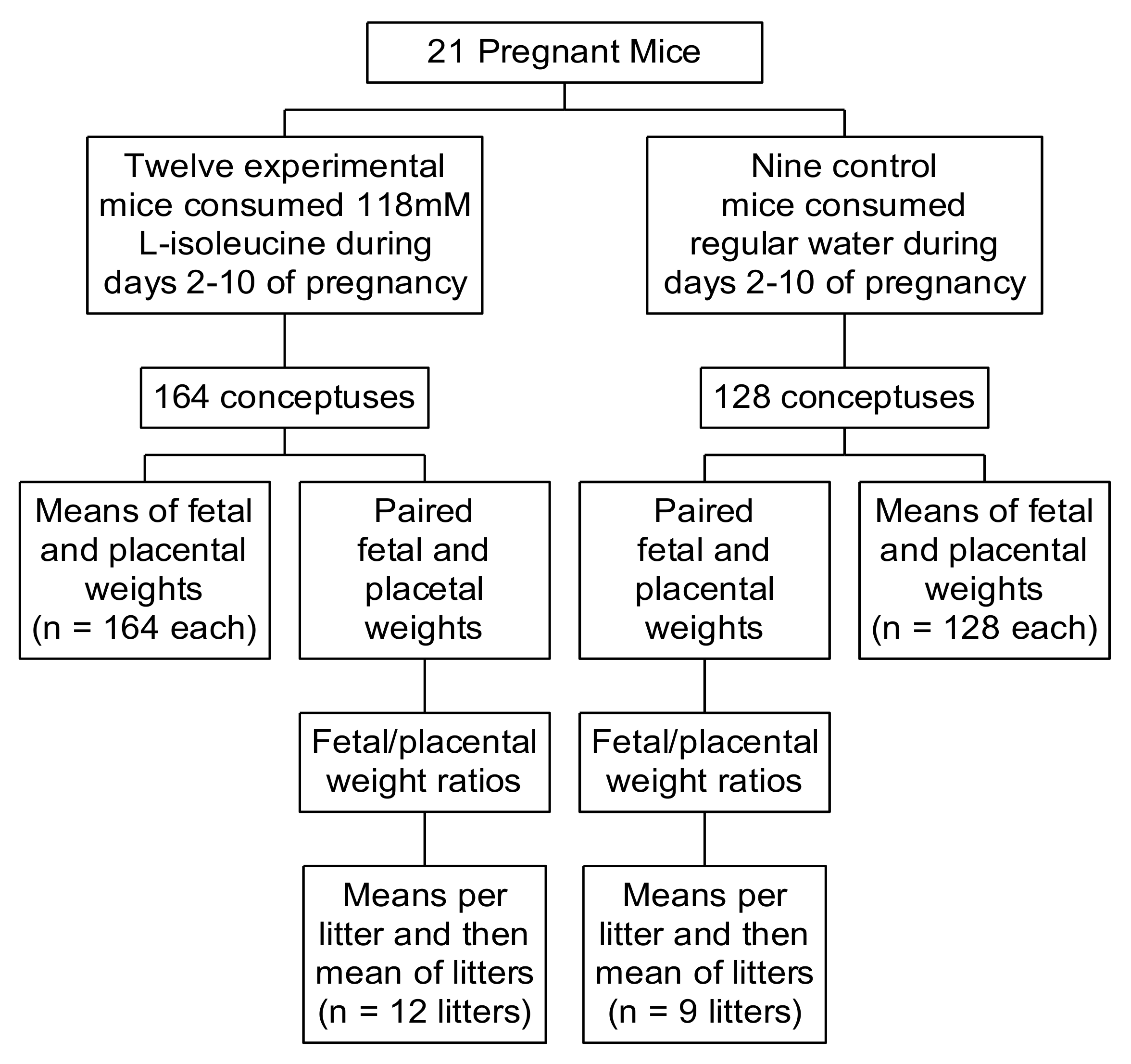
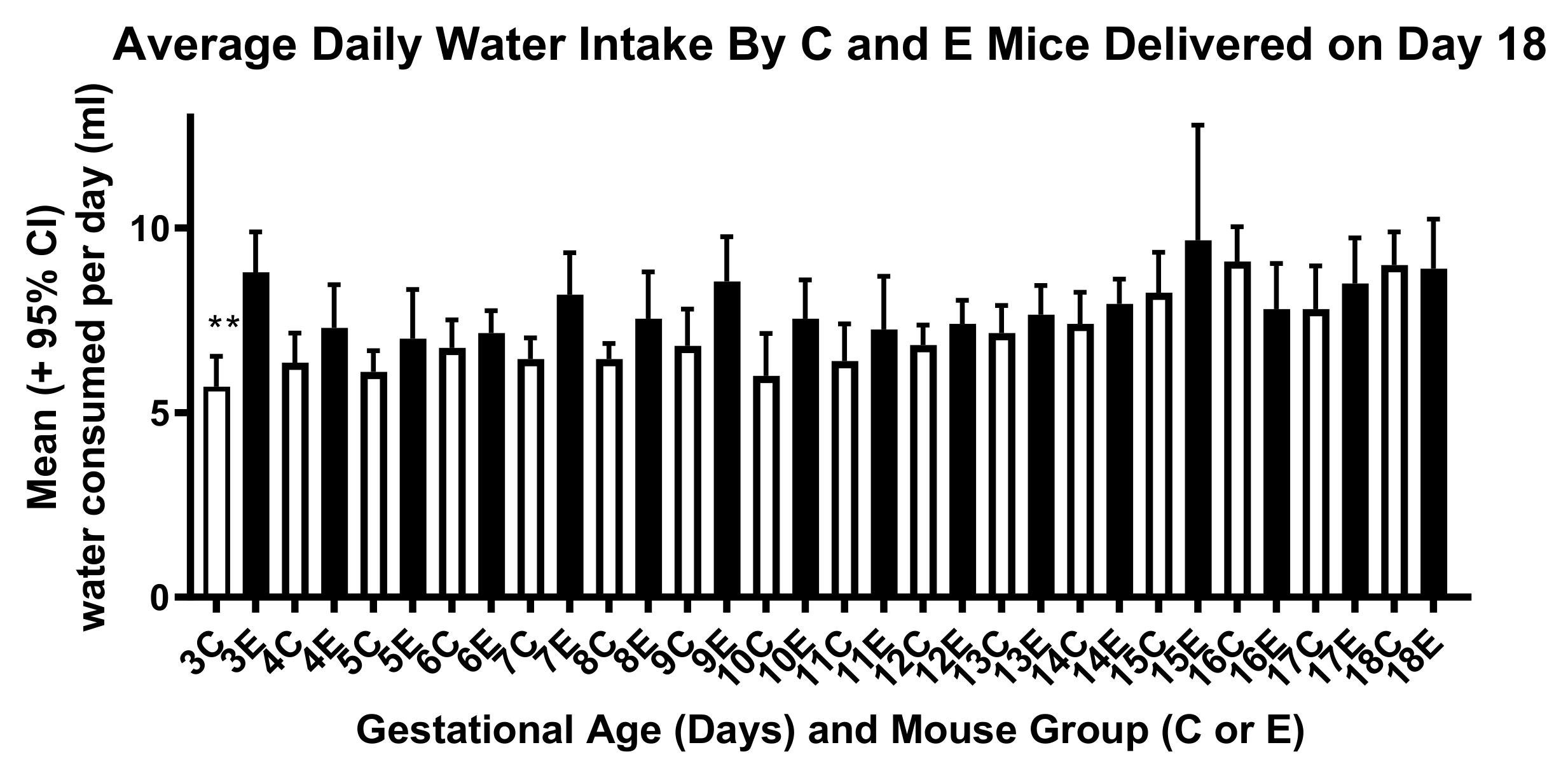
2. Methods
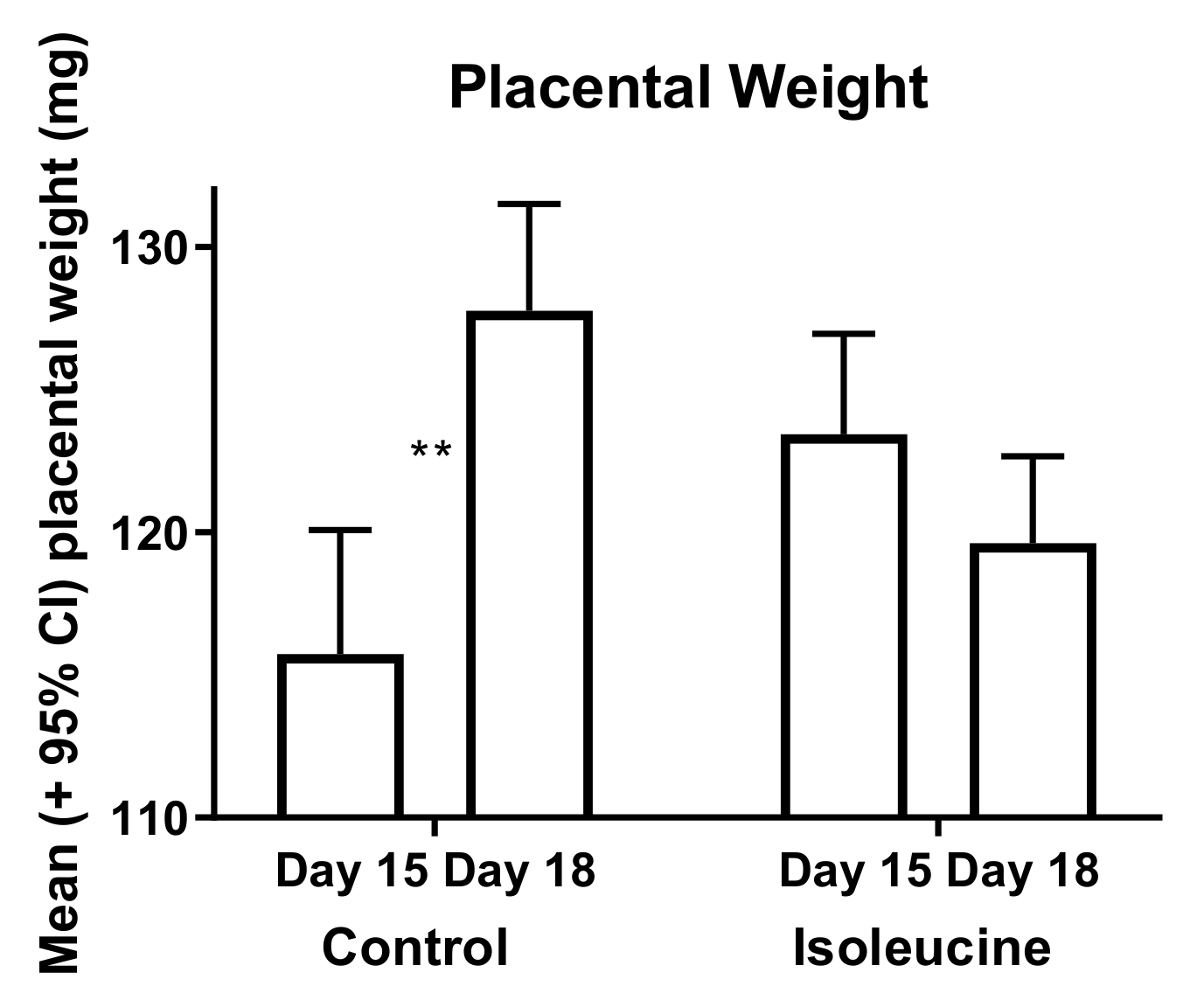
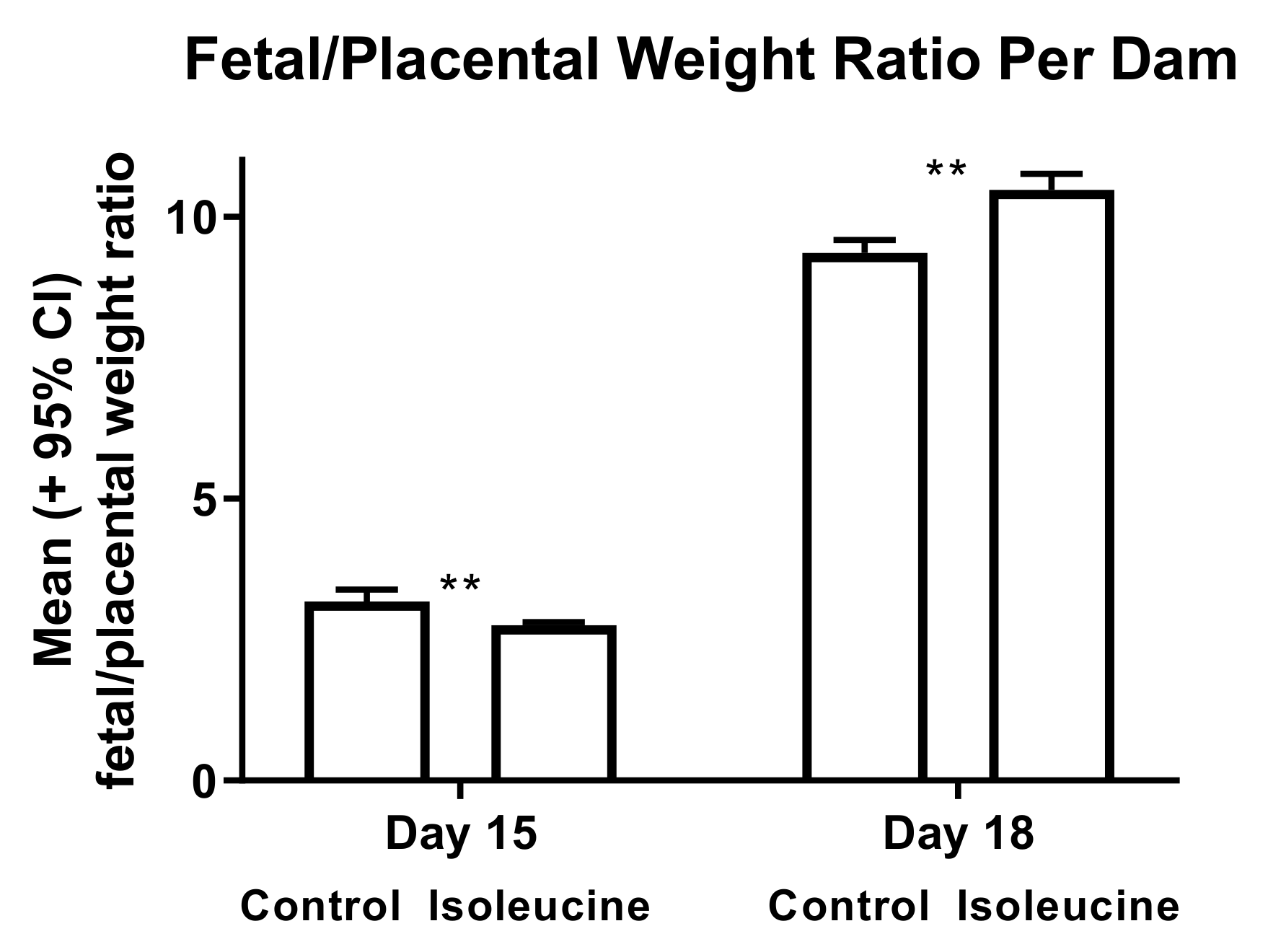
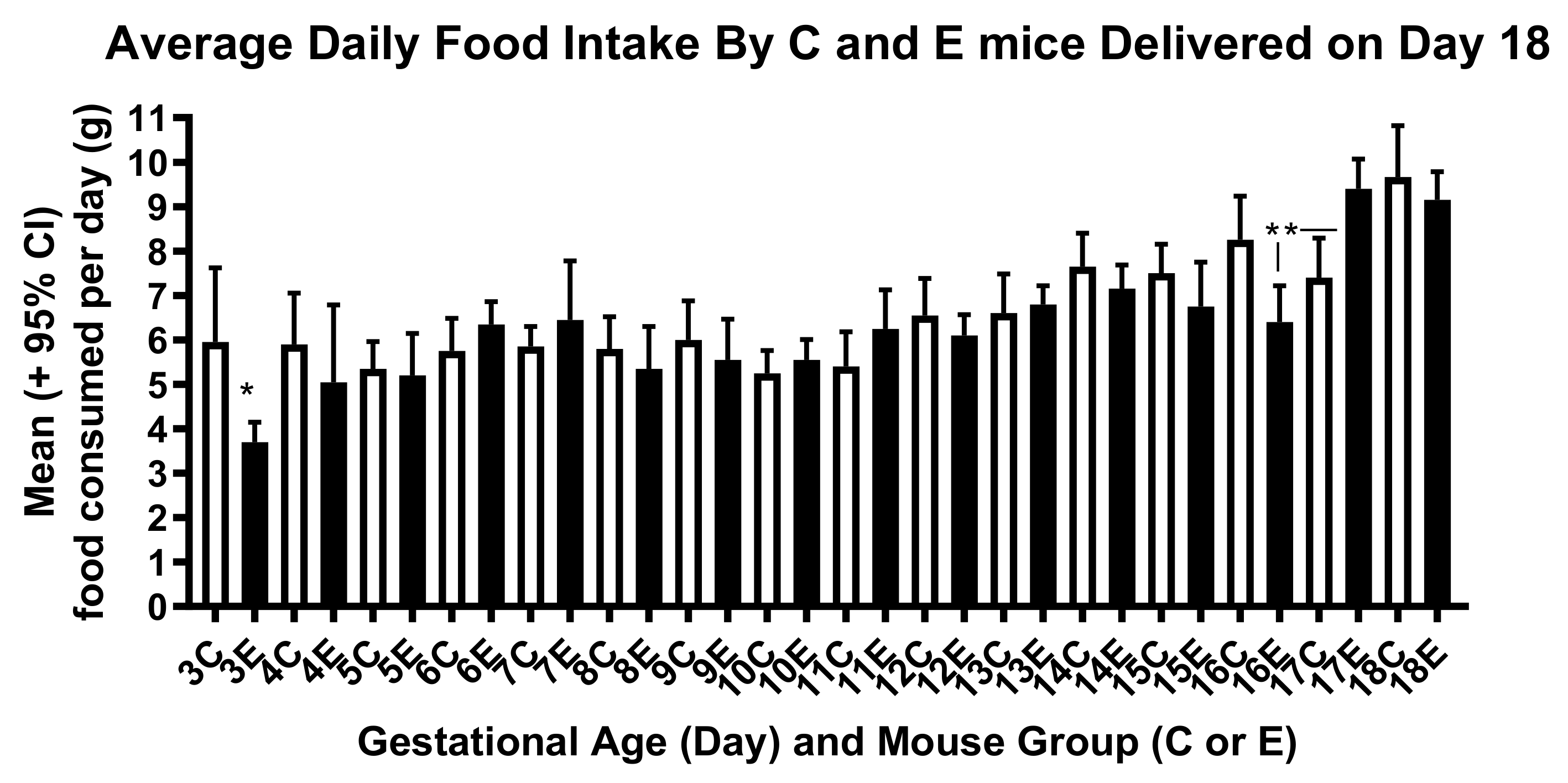
3. Results
4. Discussion
5. Limitations
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Matsumoto, K.; Koba, T.; Hamada, K.; Sakurai, M.; Higuchi, T.; Miyata, H. Branched-chain amino acid supplementation attenuates muscle soreness, muscle damage and inflammation during an intensive training program. J. Sports Med. Phys. Fit. 2009, 49, 424–431. [Google Scholar]
- Jackman, S.R.; Witard, O.C.; Jeukendrup, A.E.; Tipton, K.D. Branched-chain amino acid ingestion can ameliorate soreness from eccentric exercise. Med. Sci. Sport Exerc. 2010, 42, 962–970. [Google Scholar] [CrossRef] [PubMed]
- Fouré, A.; Bendahan, D. Is branched-chain amino acids supplementation an efficient nutritional strategy to alleviate skeletal muscle damage? A systematic review. Nutrients 2017, 9, 1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portier, H.; Chatard, J.C.; Filaire, E.; Jaunet-Devienne, M.F.; Robert, A.; Guezennec, C.Y. Effects of branched-chain amino acids supplementation on physiological and psychological performance during an offshore sailing race. Eur. J. Appl. Physiol. 2008, 104, 787–794. [Google Scholar] [CrossRef]
- Howatson, G.; Hoad, M.; Goodall, S.; Tallent, J.; Bell, P.G.; French, D.N. Exercise-induced muscle damage is reduced in resistance-trained males by branched chain amino acids: A randomized, double-blind, placebo controlled study. J. Int. Soc. Sports Nutr. 2012, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.Q.; Xun, P.; Bujnowski, D.; Daviglus, M.L.; Van Horn, L.; Stamler, J.; He, K. Higher branched-chain amino acid intake is associated with a lower prevalence of being overweight or obese in middle-aged East Asian and Western adults. J. Nutr. 2010, 141, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.C.; Li, Y.; Liu, L.Y.; Chen, Y.; Zi, T.Q.; Du, S.S.; Jiang, Y.S.; Feng, R.N.; Sun, C.H. The ratio of dietary branched-chain amino acids is associated with a lower prevalence of obesity in young northern Chinese adults: An internet-based cross-sectional study. Nutrients 2015, 7, 9573–9589. [Google Scholar] [CrossRef] [Green Version]
- Shu, X.; Kang, K.; Zhong, J.; Ji, S.; Zhang, Y.; Hu, H.; Zhang, D. Meta-analysis of branched chain amino acid-enriched nutrition to improve hepatic function in patients undergoing hepatic operation. Chin. J. Hepatol. 2014, 22, 43–47. [Google Scholar]
- Fleming, T.P.; Watkins, A.J.; Velazquez, M.A.; Mathers, J.C.; Prentice, A.M.; Stephenson, J.; Barker, M.; Saffery, R.; Yajnik, C.S.; Eckert, J.J.; et al. Origins of lifetime health around the time of conception: Causes and consequences. Lancet 2018, 391, 1842–1852. [Google Scholar] [CrossRef]
- Van Winkle, L.J.; Ryznar, R. Can uterine secretion of modified histones alter blastocyst implantation, embryo nutrition, and transgenerational phenotype? Biomol. Concepts 2018, 9, 176–183. [Google Scholar] [CrossRef]
- Van Winkle, L.J.; Ryznar, R. One-carbon metabolism regulates embryonic stem cell fate through epigenetic DNA and histone modifications: Implications for transgenerational metabolic disorders in adults. Front. Cell Dev. Biol. 2019, 7, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, W.D.; Hay, S.M.; Buchan, V.; Antipatis, C.; Palmer, R.M. The effects of maternal protein restriction on the growth of the rat fetus and its amino acid supply. Br. J. Nutr. 1999, 81, 243–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyppönen, E.; Power, C.; Smith, G.D. Prenatal growth, BMI, and risk of type 2 diabetes by early midlife. Diabets Care 2003, 26, 2512–2517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, K.; Mizuno, M.; Mizuno, T.; Dilling-Hansen, B.; Lahoz, A.; Bertelsen, V.; Münster, H.; Jordening, H.; Hamada, K.; Doi, T. Branched-chain amino acids and arginine supplementation attenuates skeletal muscle proteolysis induced by moderate exercise in young individuals. Int. J. Sports Med. 2007, 28, 531–538. [Google Scholar] [CrossRef] [Green Version]
- Van Winkle, L.J.; Tesch, J.K.; Shah, A.; Campione, A.L. System B0,+ amino acid transport regulates the penetration stage of blastocyst implantation with possible long-term developmental consequences through adulthood. Hum. Reprod. Update 2006, 12, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Van Winkle, L.J.; Mann, D.F.; Campione, A.L.; Farrington, B.H. Transport of benzenoid amino acids by system T and four broad scope systems in preimplantation mouse conceptuses. Biochim. Biophys. Acta 1990, 1027, 268–277. [Google Scholar] [CrossRef]
- Van Winkle, L.J.; Dickinson, H.R. Differences in amino acid content of preimplantation mouse embryos that develop in vitro versus in vivo: In vitro effects of five amino acids that are abundant in oviductal secretions. Biol. Reprod. 1995, 52, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Walters, E.; Edwards, R.G. On a fallacious invocation of the Barker hypothesis of anomalies in newborn rats due to mothers’ food restriction in preimplantation phases. Reprod. Biomed. Online 2003, 7, 580–582. [Google Scholar] [CrossRef]
- Kwong, W.Y.; Osmond, C.; Fleming, T.P. Support for Barker hypothesis upheld in rat model of maternal undernutrition during the preimplantation period: Application of integrated ‘random effects’ statistical model. Reprod. Biomed. Online 2004, 8, 574–576. [Google Scholar] [CrossRef]
- Walters, E.; Edwards, R.G. Further thoughts regarding evidence offered in support of the ‘Barker hypothesis’. Reprod. Biomed. Online 2004, 9, 129–131. [Google Scholar] [CrossRef]
- Hojat, M.; Xu, G. A visitor’s guide to effect sizes–statistical significance versus practical (clinical) importance of research findings. Adv. Health Sci. Educ. 2004, 9, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Watkins, A.J.; Dias, I.; Tsuro, H.; Allen, D.; Emes, R.D.; Moreton, J.; Wilson, R.; Ingram, R.J.; Sinclair, K.D. Paternal diet programs offspring health through sperm-and seminal plasma-specific pathways in mice. Proc. Natl. Acad. Sci. USA 2018, 115, 10064–10069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosario, F.J.; Jansson, N.; Kanai, Y.; Prasad, P.D.; Powell, T.L.; Jansson, T. Maternal protein restriction in the rat inhibits placental insulin, mTOR, and STAT3 signaling and down-regulates placental amino acid transporters. Endocrinology 2011, 152, 1119–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosario, F.J.; Kanai, Y.; Powell, T.L.; Jansson, T. Mammalian target of rapamycin signaling modulates amino acid uptake by regulating transporter cell surface abundance in primary human trophoblast cells. J. Physiol. 2013, 591, 609–625. [Google Scholar] [CrossRef] [PubMed]
- Rees, W.D.; Hay, S.M.; Brown, D.S.; Antipatis, C.; Palmer, R.M. Maternal protein deficiency causes hypermethylation of DNA in the livers of rat fetuses. J. Nutr. 2000, 130, 1821–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Per Offspring (g/pup) | Per Dam (g/pup/dam) | |
|---|---|---|
| C19 | 1.34 ± 0.03 a n = 60 | 1.35 ± 0.10 a n = 4 |
| C20 | 1.66 ± 0.03 b n =102 | 1.66 ± 0.06 b n = 8 |
| E19 | 1.47 ± 0.03 c n =106 | 1.47 ± 0.08 c n = 8 |
| E20 | 1.51 ± 0.05 c n = 58 | 1.52 ± 0.10 c n = 4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
To, C.Y.; Freeman, M.; Van Winkle, L.J. Consumption of a Branched-Chain Amino Acid (BCAA) during Days 2–10 of Pregnancy Causes Abnormal Fetal and Placental Growth: Implications for BCAA Supplementation in Humans. Int. J. Environ. Res. Public Health 2020, 17, 2445. https://doi.org/10.3390/ijerph17072445
To CY, Freeman M, Van Winkle LJ. Consumption of a Branched-Chain Amino Acid (BCAA) during Days 2–10 of Pregnancy Causes Abnormal Fetal and Placental Growth: Implications for BCAA Supplementation in Humans. International Journal of Environmental Research and Public Health. 2020; 17(7):2445. https://doi.org/10.3390/ijerph17072445
Chicago/Turabian StyleTo, Chiu Yuen, Muriel Freeman, and Lon J. Van Winkle. 2020. "Consumption of a Branched-Chain Amino Acid (BCAA) during Days 2–10 of Pregnancy Causes Abnormal Fetal and Placental Growth: Implications for BCAA Supplementation in Humans" International Journal of Environmental Research and Public Health 17, no. 7: 2445. https://doi.org/10.3390/ijerph17072445
APA StyleTo, C. Y., Freeman, M., & Van Winkle, L. J. (2020). Consumption of a Branched-Chain Amino Acid (BCAA) during Days 2–10 of Pregnancy Causes Abnormal Fetal and Placental Growth: Implications for BCAA Supplementation in Humans. International Journal of Environmental Research and Public Health, 17(7), 2445. https://doi.org/10.3390/ijerph17072445