Dynamics and Distribution of the Invasive Mosquito Aedes koreicus in a Temperate European City


Abstract
:1. Introduction
2. Materials and Methods
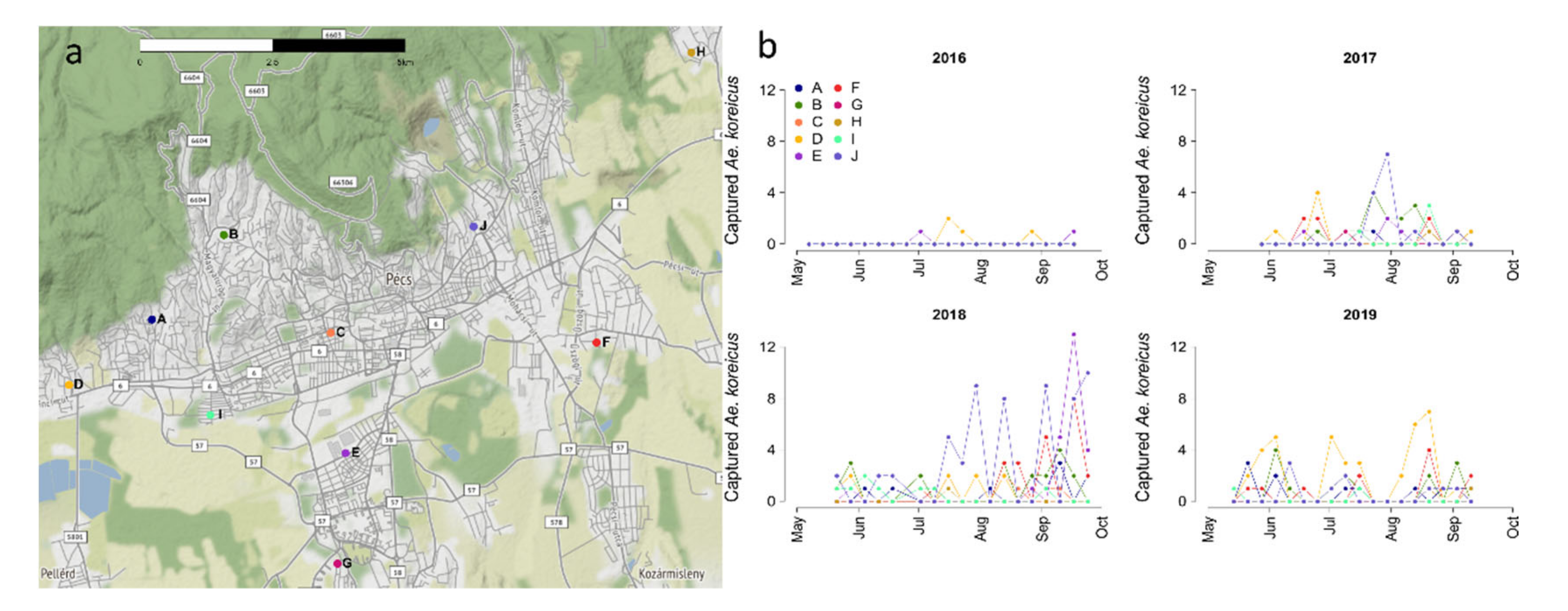
2.1. Study Area and Data Collection
2.2. Statistical and Mathematical Models
3. Results
3.1. Entomological Collections
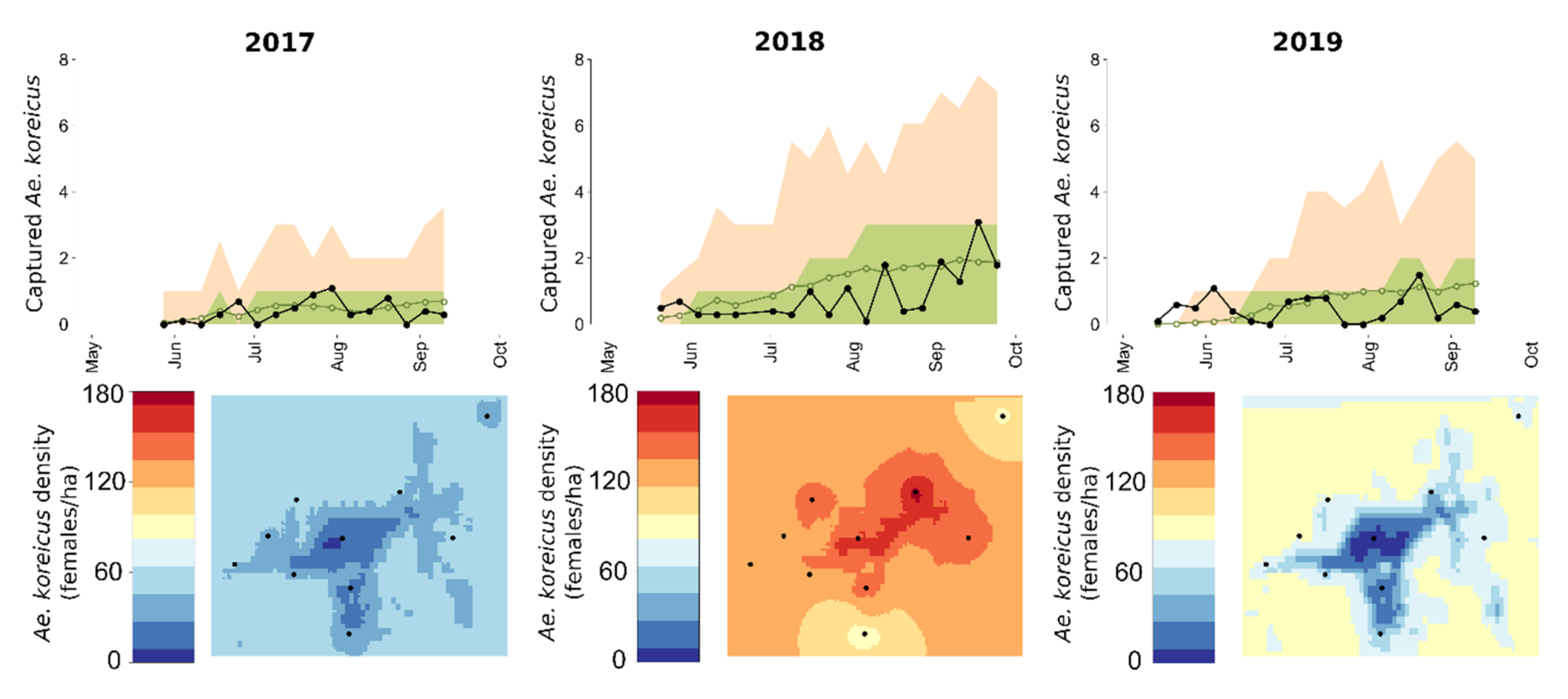
3.2. Modelling Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Medlock, J.M.; Hansford, K.M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Van Bortel, W. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector Borne Zoonotic Dis. 2012, 12, 435–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, N.; Fernández de Marco, M.; Giovannini, A.; Ippoliti, C.; Danzetta, M.; Svartz, G.; Erster, O.; Groschup, M.; Ziegler, U.; Mirazimi, A.; et al. Emerging Mosquito-Borne Threats and the Response from European and Eastern Mediterranean Countries. Int. J. Environ. Res. Public Health 2018, 15, 2775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbrink, A.; Zotzmann, S.; Cunze, S.; Klimpel, S. Aedes koreicusa new member of the genus Aedes establishing in Germany? Parasitol. Res. 2019, 118, 1073–1076. [Google Scholar] [CrossRef] [PubMed]
- Eritja, R.; Ruiz-Arrondo, I.; Delacour-Estrella, S.; Schaffner, F.; Alvarez-Chachero, J.; Bengoa, M.; Puig, M.-A.; Melero-Alcibar, R.; Oltra, A.; Bartumeus, F. First detection of Aedes japonicus in Spain: An unexpected finding triggered by citizen science. Parasit. Vectors 2019, 12, 53. [Google Scholar] [CrossRef]
- Pfitzner, W.P.; Lehner, A.; Hoffmann, D.; Czajka, C.; Becker, N. First record and morphological characterization of an established population of Aedes (Hulecoeteomyia) koreicus (Diptera: Culicidae) in Germany. Parasit. Vectors 2018, 11, 662. [Google Scholar] [CrossRef]
- Seidel, B.; Montarsi, F.; Huemer, H.P.; Indra, A.; Capelli, G.; Allerberger, F.; Nowotny, N. First record of the Asian bush mosquito, Aedes japonicus japonicus, in Italy: Invasion from an established Austrian population. Parasit. Vectors 2016, 9, 284. [Google Scholar] [CrossRef] [Green Version]
- Wilson, A.L.; Courtenay, O.; Kelly-Hope, L.A.; Scott, T.W.; Takken, W.; Torr, S.J.; Lindsay, S.W. The importance of vector control for the control and elimination of vector-borne diseases. PLoS Negl. Trop. Dis. 2020, 14, e0007831. [Google Scholar] [CrossRef] [Green Version]
- Papa, A. Emerging arboviruses of medical importance in the Mediterranean region. J. Clin. Virol. 2019, 115, 5–10. [Google Scholar] [CrossRef]
- Riccardo, F.; Monaco, F.; Bella, A.; Savini, G.; Russo, F.; Cagarelli, R.; Dottori, M.; Rizzo, C.; Venturi, G.; Luca, M.D.; et al. An early start of West Nile virus seasonal transmission: The added value of One Heath surveillance in detecting early circulation and triggering timely response in Italy, June to July 2018. Eurosurveillance 2018, 23, 1800427. [Google Scholar] [CrossRef] [Green Version]
- Mosquito Maps. Available online: https://www.ecdc.europa.eu/en/disease-vectors/surveillance-and-disease-data/mosquito-maps (accessed on 11 March 2020).
- Versteirt, V.; Pecor, J.E.; Fonseca, D.M.; Coosemans, M.; Bortel, W.V. Confirmation of Aedes koreicus (Diptera: Culicidae) in Belgium and description of morphological differences between Korean and Belgian specimens validated by molecular identification. Zootaxa 2012, 3191, 21–32. [Google Scholar] [CrossRef]
- Tanaka, K.; Mizusawa, K.; Saugstad, E. A revision of the adult and larval mosquitoes of Japan (Including the Ryukyu Archipelago and the Ogasawara Islands) and Korea (Diptera: Culicidae). Contr. Am. Entomol. Inst. 1979, 16, 1–989. [Google Scholar]
- Kurucz, K.; Kiss, V.; Zana, B.; Schmieder, V.; Kepner, A.; Jakab, F.; Kemenesi, G. Emergence of Aedes koreicus (Diptera: Culicidae) in an urban area, Hungary, 2016. Parasitol. Res. 2016, 115, 4687–4689. [Google Scholar] [CrossRef]
- Kurucz, K.; Kiss, V.; Zana, B.; Jakab, F.; Kemenesi, G. Filarial nematode (order: Spirurida) surveillance in urban habitats, in the city of Pécs (Hungary). Parasitol. Res. 2018, 117, 3355–3360. [Google Scholar] [CrossRef] [PubMed]
- Knight, K.L. Contributions to the mosquito fauna of Southeast Asia.—IV. Species of subgroup Chrysolineatus of group D, genus Aedes, subgenus Finlaya Theobald. Contr. Am. Entomol. Inst. 1968, 2, 1–45. [Google Scholar]
- Montarsi, F.; Ciocchetta, S.; Devine, G.; Ravagnan, S.; Mutinelli, F.; Frangipane di Regalbono, A.; Otranto, D.; Capelli, G. Development of Dirofilaria immitis within the mosquito Aedes (Finlaya) koreicus, a new invasive species for Europe. Parasit. Vectors 2015, 8, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciocchetta, S.; Prow, N.A.; Darbro, J.M.; Frentiu, F.D.; Savino, S.; Montarsi, F.; Capelli, G.; Aaskov, J.G.; Devine, G.J. The new European invader Aedes (Finlaya) koreicus: A potential vector of chikungunya virus. Pathog. Glob. Health 2018, 112, 107–114. [Google Scholar] [CrossRef]
- Autochthonous Transmission of Chikungunya Virus in EU/EEA, 2007–2017. Available online: https://www.ecdc.europa.eu/en/all-topics-z/chikungunya-virus-disease/surveillance-and-disease-data/autochthonous-transmission (accessed on 11 March 2020).
- Alfano, N.; Tagliapietra, V.; Rosso, F.; Manica, M.; Arnoldi, D.; Pindo, M.; Rizzoli, A. Changes in Microbiota Across Developmental Stages of Aedes koreicus, an Invasive Mosquito Vector in Europe: Indications for Microbiota-Based Control Strategies. Front. Microbiol. 2019, 10, 2832. [Google Scholar] [CrossRef]
- Kim, H.C.; Lee, K.W.; Richards, R.S.; Schleich, S.S.; Herman, W.E.; Klein, T.A. Seasonal Prevalence of Mosquitoes Collected from Light Traps in Korea (1999–2000). Entomol. Res. 2003, 33, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Marini, G.; Arnoldi, D.; Baldacchino, F.; Capelli, G.; Guzzetta, G.; Merler, S.; Montarsi, F.; Rizzoli, A.; Rosà, R. First report of the influence of temperature on the bionomics and population dynamics of Aedes koreicus, a new invasive alien species in Europe. Parasit. Vectors 2019, 12, 524. [Google Scholar] [CrossRef]
- Poletti, P.; Messeri, G.; Ajelli, M.; Vallorani, R.; Rizzo, C.; Merler, S. Transmission Potential of Chikungunya Virus and Control Measures: The Case of Italy. PLoS ONE 2011, 6, e18860. [Google Scholar] [CrossRef]
- Guzzetta, G.; Trentini, F.; Poletti, P.; Baldacchino, F.A.; Montarsi, F.; Capelli, G.; Rizzoli, A.; Rosà, R.; Merler, S.; Melegaro, A. Effectiveness and economic assessment of routine larviciding for prevention of chikungunya and dengue in temperate urban settings in Europe. PLoS Negl. Trop. Dis. 2017, 11, e0005918. [Google Scholar] [CrossRef] [PubMed]
- Marini, G.; Guzzetta, G.; Toledo, C.A.M.; Teixeira, M.; Rosà, R.; Merler, S. Effectiveness of Ultra-Low Volume insecticide spraying to prevent dengue in a non-endemic metropolitan area of Brazil. PLOS Comput. Biol. 2019, 15, e1006831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marini, G.; Calzolari, M.; Angelini, P.; Bellini, R.; Bellini, S.; Bolzoni, L.; Torri, D.; Defilippo, F.; Dorigatti, I.; Nikolay, B.; et al. A quantitative comparison of West Nile virus incidence from 2013 to 2018 in Emilia-Romagna, Italy. PLoS Negl. Trop. Dis. 2020, 14, e0007953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delatte, H.; Gimonneau, G.; Triboire, A.; Fontenille, D. Influence of temperature on immature development, survival, longevity, fecundity, and gonotrophic cycles of Aedes albopictus, vector of chikungunya and dengue in the Indian Ocean. J. Med. Entomol. 2009, 46, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Reuss, F.; Wieser, A.; Niamir, A.; Bálint, M.; Kuch, U.; Pfenninger, M.; Müller, R. Thermal experiments with the Asian bush mosquito (Aedes japonicus japonicus) (Diptera: Culicidae) and implications for its distribution in Germany. Parasit. Vectors 2018, 11, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loetti, V.; Schweigmann, N.; Burroni, N. Development rates, larval survivorship and wing length of Culex pipiens (Diptera: Culicidae) at constant temperatures. J. Nat. Hist. 2011, 45, 2203–2213. [Google Scholar] [CrossRef]
- Gazetteer of Hungary, 1 January 2019; Hungarian Central Statistical Office: Budapest, Hungary; ISSN 1217P–2952. 2019. (In Hungarian)
- Climate Characteristics of Pécs. Available online: https://www.met.hu/eghajlat/magyarorszag_eghajlata/varosok_jellemzoi/Pecs/ (accessed on 11 March 2020).
- OpenStreetMap. Available online: https://www.openstreetmap.org/copyright (accessed on 11 March 2020).
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes and Their Control; Springer: Berlin/Heidelberg, Germany, 2010; ISBN 978-3-540-92873-7. [Google Scholar]
- European Centre for Disease Prevention and Control and European Food Safety Authority (Ed.) Guidelines for the surveillance of invasive mosquitoes in Europe; ECDC technical report; ECDC: Stockholm, Sweden, 2012; ISBN 978-92-9193-378-5. [Google Scholar]
- Metz, M.; Rocchini, D.; Neteler, M. Surface Temperatures at the Continental Scale: Tracking Changes with Remote Sensing at Unprecedented Detail. Remote Sens. 2014, 6, 3822–3840. [Google Scholar] [CrossRef] [Green Version]
- CORINE Land Cover. Available online: https://land.copernicus.eu/pan-european/corine-land-cover (accessed on 11 March 2020).
- WorldPop. Available online: https://www.worldpop.org/ (accessed on 11 March 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: http://www.R-project.org/ (accessed on 14 April 2020).
- Robert, J.; Hijmans. Raster: Geographic Data Analysis and Modeling. R package version 2.9-22. Available online: https://CRAN.R-project.org/package=raster (accessed on 14 April 2020).
- Hiemstra, P.H.; Pebesma, E.J.; Twenhöfel, C.J.W.; Heuvelink, G.B.M. Real-time automatic interpolation of ambient gamma dose rates from the Dutch radioactivity monitoring network. Comput. & Geosci. 2009, 35, 1711–1721. [Google Scholar]
- Venables, W.N.; Ripley, B.D.; Venables, W.N. Modern applied statistics with S, Statistics and computing, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 978-0-387-95457-8. [Google Scholar]
- Lühken, R.; Pfitzner, W.P.; Börstler, J.; Garms, R.; Huber, K.; Schork, N.; Steinke, S.; Kiel, E.; Becker, N.; Tannich, E.; et al. Field evaluation of four widely used mosquito traps in Central Europe. Parasit. Vectors 2014, 7, 268. [Google Scholar] [CrossRef] [Green Version]
- Baldacchino, F.; Arnoldi, D.; Lapère, C.; Rosà, R.; Montarsi, F.; Capelli, G.; Rizzoli, A. Weak Larval Competition Between Two Invasive Mosquitoes Aedes koreicus and Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2017, 54, 1266–1272. [Google Scholar] [CrossRef]
- Montarsi, F.; Martini, S.; Dal Pont, M.; Delai, N.; Ferro Milone, N.; Mazzucato, M.; Soppelsa, F.; Cazzola, L.; Cazzin, S.; Ravagnan, S.; et al. Distribution and habitat characterization of the recently introduced invasive mosquito Aedes koreicus [Hulecoeteomyia koreica], a new potential vector and pest in north-eastern Italy. Parasit. Vectors 2013, 6, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurucz, K.; Kepner, A.; Krtinic, B.; Hederics, D.; Foldes, F.; Brigetta, Z.; Jakab, F.; Kemenesi, G. Blood-meal analysis and avian malaria screening of mosquitoes collected from human-inhabited areas in Hungary and Serbia. J. Eur. Mosq. Control Assoc. 2018, 36, 3–13. [Google Scholar]
- Jourdain, F.; Samy, A.M.; Hamidi, A.; Bouattour, A.; Alten, B.; Faraj, C.; Roiz, D.; Petrić, D.; Pérez-Ramírez, E.; Velo, E.; et al. Towards harmonisation of entomological surveillance in the Mediterranean area. PLoS Negl. Trop. Dis. 2019, 13, e0007314. [Google Scholar] [CrossRef] [PubMed]
- Takken, W.; van den Berg, H. Manual on Prevention of Establishment and Control of Mosquitoes of Public Health Importance in the WHO European Region (with Special Reference to Invasive Mosquitoes); World Health Organization: Copenhagen, Denmark, 2018; ISBN 978-92-890-5388-4. [Google Scholar]
- Roiz, D.; Wilson, A.L.; Scott, T.W.; Fonseca, D.M.; Jourdain, F.; Müller, P.; Velayudhan, R.; Corbel, V. Integrated Aedes management for the control of Aedes-borne diseases. PLoS Negl. Trop. Dis. 2018, 12, e0006845. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Explanatory Variable | Statistical Test/Model | Coefficients Estimate | p-Value |
|---|---|---|---|
| Average spring temperature | Negative Binomial GLM | Intercept = –1.47 | 0.565 0.174 |
| Coef = 0.21 | |||
| Average summer temperature | Negative Binomial GLM | Intercept = 16.57 | 0.005 0.016 |
| Coef = –0.57 | |||
| Human population | Kendall correlation test | τ = –0.29 | 0.03 |
| Vegetation cover | Kendall correlation test | τ = 0.09 | 0.49 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurucz, K.; Manica, M.; Delucchi, L.; Kemenesi, G.; Marini, G. Dynamics and Distribution of the Invasive Mosquito Aedes koreicus in a Temperate European City. Int. J. Environ. Res. Public Health 2020, 17, 2728. https://doi.org/10.3390/ijerph17082728
Kurucz K, Manica M, Delucchi L, Kemenesi G, Marini G. Dynamics and Distribution of the Invasive Mosquito Aedes koreicus in a Temperate European City. International Journal of Environmental Research and Public Health. 2020; 17(8):2728. https://doi.org/10.3390/ijerph17082728
Chicago/Turabian StyleKurucz, Kornélia, Mattia Manica, Luca Delucchi, Gábor Kemenesi, and Giovanni Marini. 2020. "Dynamics and Distribution of the Invasive Mosquito Aedes koreicus in a Temperate European City" International Journal of Environmental Research and Public Health 17, no. 8: 2728. https://doi.org/10.3390/ijerph17082728
APA StyleKurucz, K., Manica, M., Delucchi, L., Kemenesi, G., & Marini, G. (2020). Dynamics and Distribution of the Invasive Mosquito Aedes koreicus in a Temperate European City. International Journal of Environmental Research and Public Health, 17(8), 2728. https://doi.org/10.3390/ijerph17082728