
Oncogenic Role of miR-200c-3p in High-Grade Serous Ovarian Cancer Progression via Targeting the 3′-Untranslated Region of DLC1

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Human Tissues and Cell Lines
2.3. RNA isolation and quantitative Real Time-PCR (qRT-PCR)
2.4. Transient miR-200c-3p transfection
2.5. Protein Extraction and Western Blot Assay
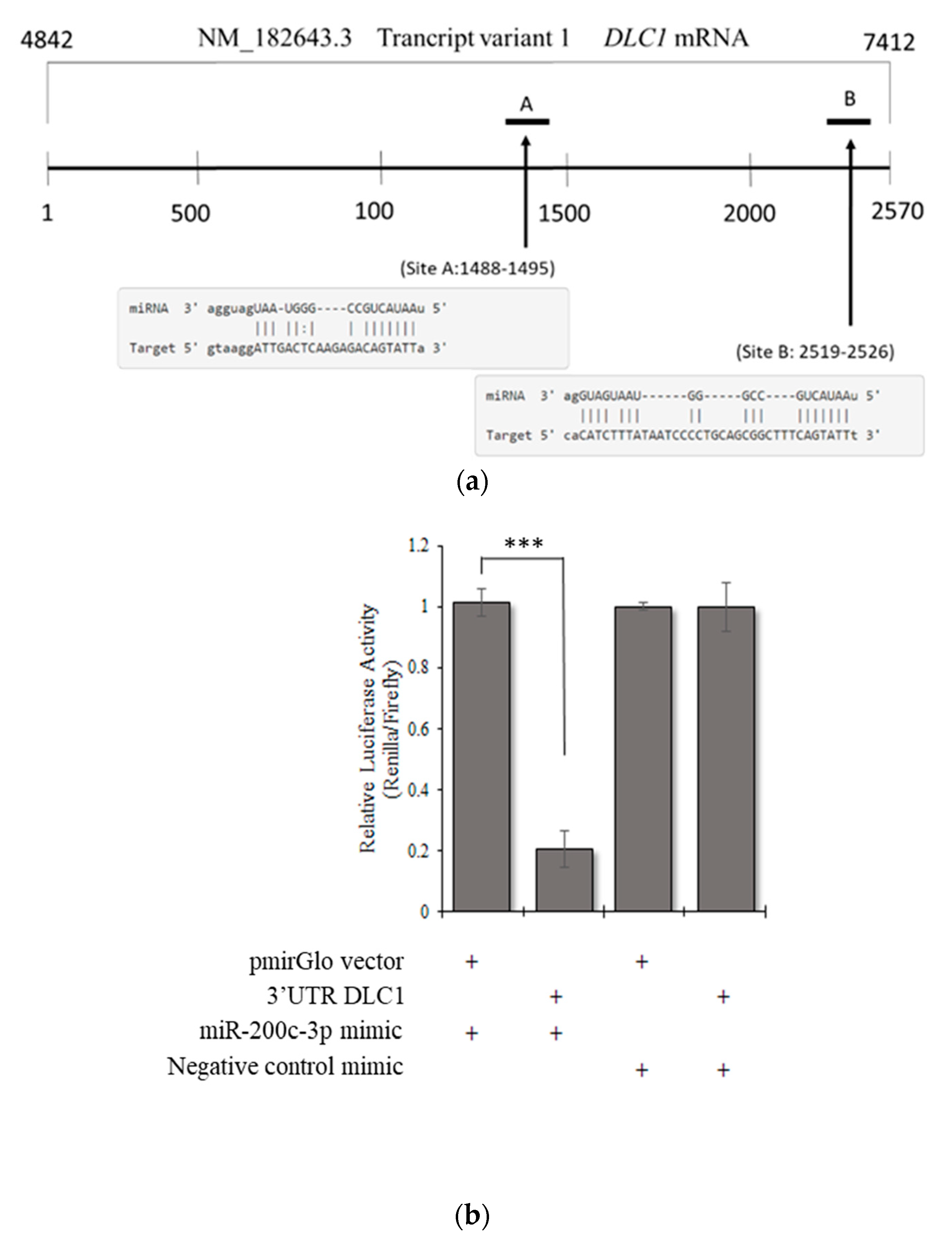
2.6. Prediction of miRNA target sites in the 3′UTR of DLC1
2.7. Cloning of the 3′UTR of DLC1
2.8. Luciferase Reporter Assay
2.9. Statistical Analysis
3. Results
3.1. Clinical and Pathological Characteristics of Samples
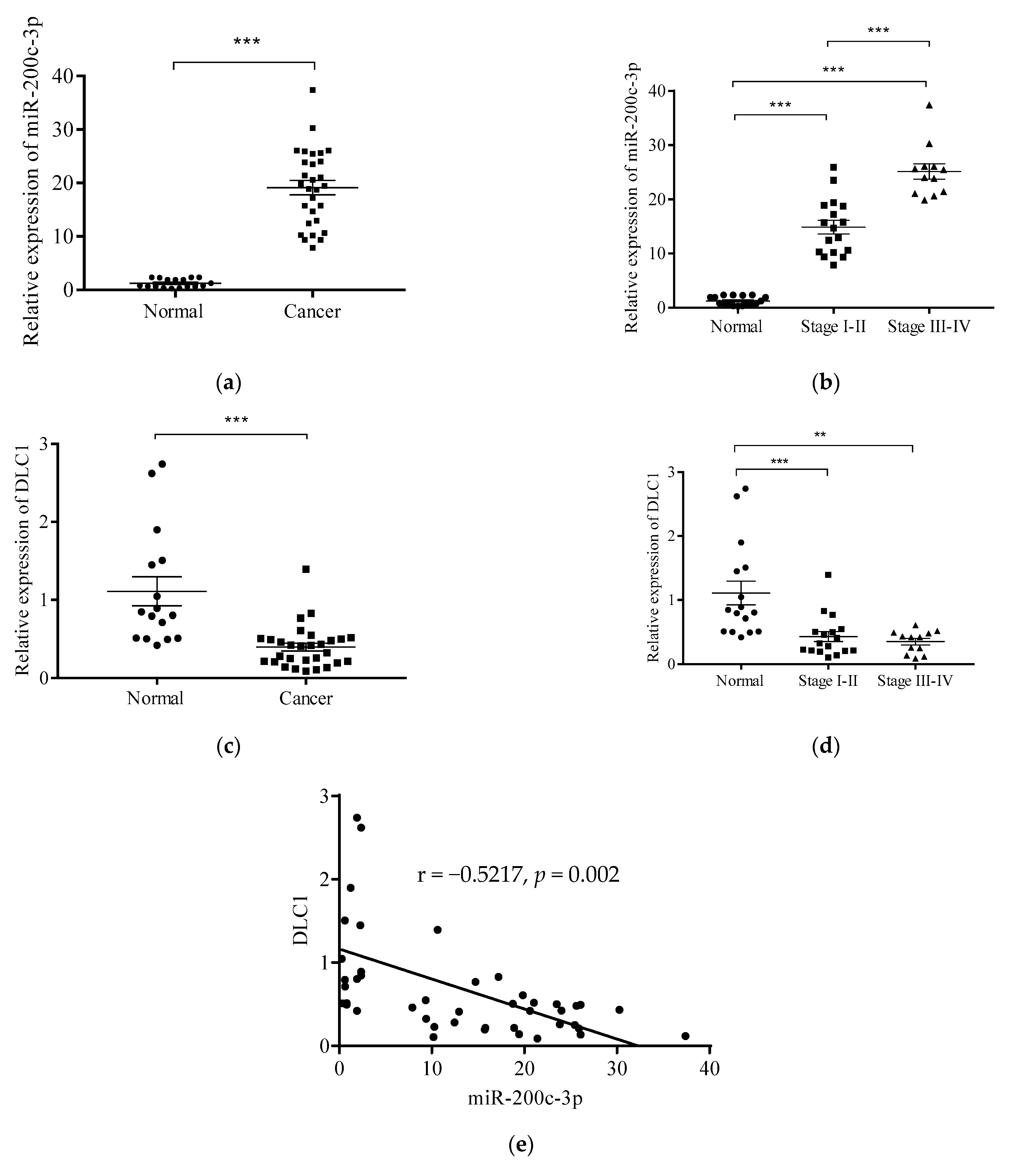
3.2. Expression Levels of miR-200c-3p and DLC1 in Clinical Specimens.
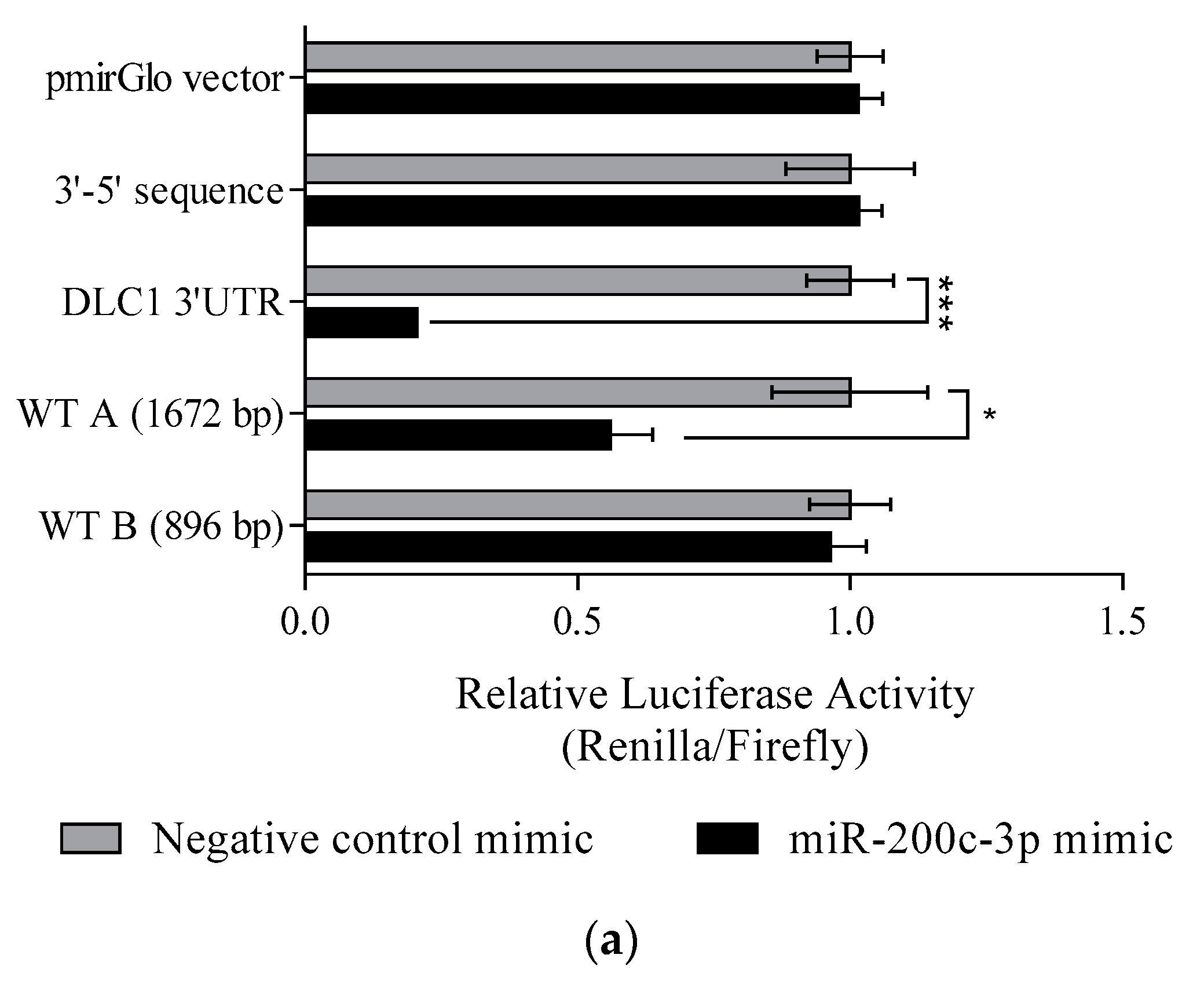
3.3. DLC1 Is a Direct Target of miR-200c-3p in an In Vitro Model of HGSC
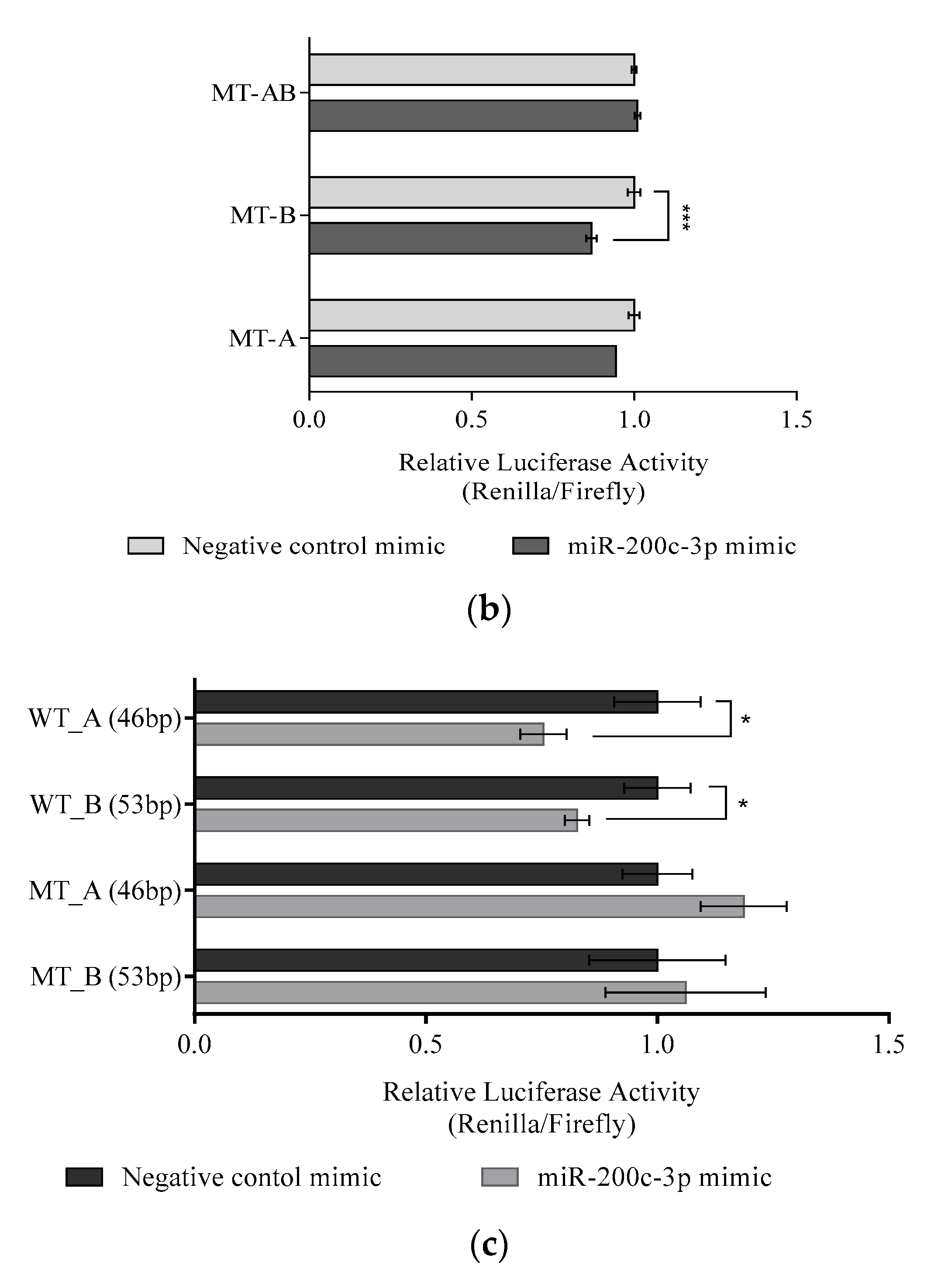
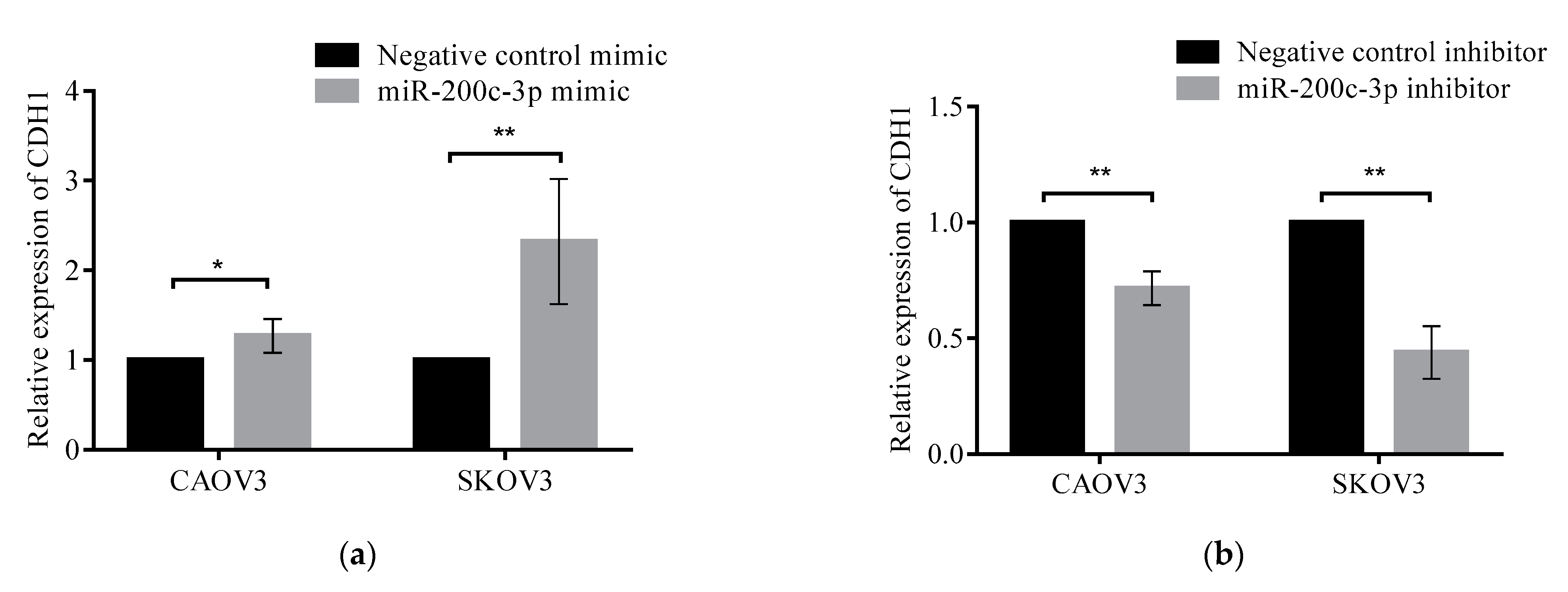
3.4. Regulation of DLC1 by miR-200c-3p in HGSC Cell Lines
3.5. Regulation of miR-200c-3p in Epithelial-Mesenchymal Transition through Modulation of DLC1 Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Torre, L.A.; Trabert, B.; DeSantis, C.E.; Miller, K.D.; Samimi, G.; Runowicz, C.D.; Gaudet, M.M.; Jemal, A.; Siegel, R.L. Ovarian cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 284–296. [Google Scholar] [CrossRef]
- Koshiyama, M.; Matsumura, N.; Konishi, I. Recent concepts of ovarian carcinogenesis: Type I and type II. BioMed Res. Int. 2014, 2014, 934261. [Google Scholar] [CrossRef] [Green Version]
- Jayson, G.C.; Kohn, E.C.; Kitchener, H.C.; Ledermann, J.A. Ovarian cancer. Lancet 2014, 384, 1376–1388. [Google Scholar] [CrossRef]
- Kurman, R.J.; Shih, I.-M. The dualistic model of ovarian carcinogenesis revisited, revised, and expanded. Am. J. Pathol. 2016, 186, 733–747. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.; Ralyea, C.; Lockwood, S. Ovarian Cancer: An Integrated Review. Semin. Oncol. Nurs. 2019, 35, 151–156. [Google Scholar] [CrossRef]
- Vang, R.; Levine, D.A.; Soslow, R.A.; Zaloudek, C.; Shih, I.-M.; Kurman, R.J. Molecular Alterations of TP53 are a Defining Feature of Ovarian High-Grade Serous Carcinoma: A Rereview of Cases Lacking TP53 Mutations in The Cancer Genome Atlas Ovarian Study Russell. Int. J. Gynecol. Pathol. 2016, 35, 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurman, R.J. Origin and molecular pathogenesis of ovarian high-grade serous carcinoma. Ann. Oncol. 2013, 24, x16–x21. [Google Scholar] [CrossRef]
- Motohara, T.; Masuda, K.; Morotti, M.; Zheng, Y.; El-Sahhar, S.; Chong, K.Y.; Wietek, N.; Alsaadi, A.; Karaminejadranjbar, M.; Hu, Z.; et al. An evolving story of the metastatic voyage of ovarian cancer cells: Cellular and molecular orchestration of the adipose-rich metastatic microenvironment. Oncogene 2019, 38, 2885–2898. [Google Scholar] [CrossRef] [Green Version]
- Motohara, T.; Fujimoto, K.; Tayama, S.; Narantuya, D.; Sakaguchi, I.; Tashiro, H.; Katabuchi, H. CD44 variant 6 as a predictive biomarker for distant metastasis in patients with epithelial ovarian cancer. Obstet. Gynecol. 2016, 127, 1003–1011. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Ankasha, S.J.; Shafiee, M.N.; Wahab, N.A.; Ali, R.A.R.; Mokhtar, N.M. Post-transcriptional regulation of microRNAs in cancer: From prediction to validation. Oncol. Rev. 2018, 12, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Dweep, H.; Sticht, C.; Gretz, N. In-Silico Algorithms for the Screening of Possible microRNA Binding Sites and Their Interactions. Curr. Genom. 2013, 14, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Lai, E.C. Predicting and validating microRNA targets. Genome Biol. 2004, 5, 115. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, F.F.; Jamal, R.; Effendi Syafruddin, S.; Syakima Ab Mutalib, N.; Saidin, S.; Rahayu MdZin, R.; Manir Hossain Mollah, M.; Mohd Mokhtar, N. MicroRNA-200c and microRNA-31 regulate proliferation, colony formation, migration and invasion in serous ovarian cancer. J. Ovarian Res. 2015, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Wahab, N.A.; Othman, Z.; Mohd Nasri, N.W.; Mokhtar, M.H.; Ibrahim, S.F.; Hamid, A.A.; Ali, R.A.R.; Mokhtar, N.M. Inhibition of mir-141 and mir-200a increase dlc-1 and zeb2 expression, enhance migration and invasion in metastatic serous ovarian cancer. Int. J. Environ. Res. Public Health 2020, 17, 2766. [Google Scholar] [CrossRef] [Green Version]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microRNA org resource: Targets and expression. Nucleic Acids Res. 2008, 36, D149–D153. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. ELife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Vilming Elgaaen, B.; Olstad, O.K.; Haug, K.B.F.; Brusletto, B.; Sandvik, L.; Staff, A.C.; Gautvik, K.M.; Davidson, B. Global miRNA expression analysis of serous and clear cell ovarian carcinomas identifies differentially expressed miRNAs including miR-200c-3p as a prognostic marker. BMC Cancer 2014, 14, 80. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Lu, Z.; Unruh, A.K.; Ivan, C.; Baggerly, K.A.; Calin, G.A.; Li, Z.; Bast, R.C.; Le, X.-F. Clinically Relevant microRNAs in Ovarian Cancer. Mol. Cancer Res. 2015, 13, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Müller, V.; Milde-Langosch, K.; Trillsch, F.; Pantel, K.; Schwarzenbach, H. Circulating cell-free miR-373, miR- 200a, miR-200b and miR-200c in patients with epithelial ovarian cancer. Adv. Exp. Med. Biol. 2016, 924, 3–8. [Google Scholar]
- Kim, T.Y.; Vigil, D.; Der, C.J.; Juliano, R.L. Role of DLC-1, a tumor suppressor protein with RhoGAP activity, in regulation of the cytoskeleton and cell motility. Cancer Metastasis Rev. 2009, 28, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, Y.; Yuan, S.Y.; Hao, S. Down-regulation of DLC1 in endothelial cells compromises the angiogenesis process. Cancer Lett. 2017, 398, 46–51. [Google Scholar] [CrossRef]
- Wang, D.; Qian, X.; Rajaram, M.; Durkin, M.E.; Lowy, D.R. DLC1 is the principal biologically-relevant down-regulated DLC family member in several cancers. Oncotarget 2016, 7, 45144–45157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.; Cho, S.; Kim, H.; Na, D.; Yun, J.; Chae, J.; Park, C. Genomic alterations in BCL2L1 and DLC1 contribute to drug sensitivity in gastric cancer. Proc. Natl. Acad. Sci. USA 2015, 112, 12492–12497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, J.S.W.; Tao, Q.; Ng, K.M.; Goh, H.K.; Shu, X.; Woo, W.L.; Ambinder, R.F.; Srivastava, G.; Shamay, M.; Chan, A.T.C.; et al. A novel isoform of the 8p22 tumor suppressor gene DLC1 suppresses tumor growth and is frequently silenced in multiple common tumors. Oncogene 2011, 30, 1923–1935. [Google Scholar] [CrossRef] [Green Version]
- Hassan, N.Z.A.; Mokhtar, N.M.; Sin, T.K.; Rose, I.M.; Sagap, I.; Harun, R.; Jamal, R. Integrated analysis of copy number variation and genome-wide expression profiling in colorectal cancer tissues. PLoS ONE 2014, 9, e92553. [Google Scholar] [CrossRef]
- Fan, D.; Shi, H. Pilot Study: Alteration of Deleted in Liver Cancer1 and Phosphorylated Focal Adhesion Kinase Y397 Cytoplasmic Expression and the Prognostic Value in Advanced Epithelial Ovarian Carcinoma. Int. J. Mol. Sci. 2011, 12, 8489–8501. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, J.; Luo, H. Clinicopathological Significance of DLC-1 Expression in Cancer: A Meta-Analysis. Asian Pac. J. Cancer Prev. 2015, 16, 7255–7260. [Google Scholar] [CrossRef]
- Grimson, A.; Farh, K.K.H.; Johnston, W.K.; Garrett-Engele, P.; Lim, L.P.; Bartel, D.P. MicroRNA Targeting Specificity in Mammals: Determinants beyond Seed Pairing. Mol. Cell 2007, 27, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Sætrom, P. MicroRNAs—Targeting and target prediction. New Biotechnol. 2010, 27, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Mak, M.P.; Tong, P.; Diao, L.; Cardnell, R.J.; Gibbons, D.L.; William, N.; Skoulidis, F.; Parra, E.R.; Rodriguez-canales, J.; Wistuba, I.I. A patient-derived, pan-cancer EMT signature identifies global molecular alterations and immune target enrichment following epithelial to mesenchymal transition. Clin. Cancer Res. 2015, 22, 609–620. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Liu, J.; Feng, X.; Chen, H.; Zeng, L.; Huang, G.; Liu, W.; Wang, L.; Jia, W.; Chen, J.; et al. DLC-1 induces mitochondrial apoptosis and epithelial mesenchymal transition arrest in nasopharyngeal carcinoma by targeting EGFR/Akt/NF-κB pathway. Med. Oncol. 2015, 32, 115. [Google Scholar] [CrossRef] [PubMed]
- Asnaghi, L.; Vass, W.C.; Quadri, R.; Day, P.M.; Qian, X.; Braverman, R.; Papageorge, A.G.; Lowy, D.R. E-cadherin negatively regulates neoplastic growth in non-small cell lung cancer: Role of Rho GTPases. Oncogene 2010, 29, 2760–2771. [Google Scholar] [CrossRef] [Green Version]
- Hur, K.; Toiyama, Y.; Takahashi, M.; Balaguer, F.; Nagasaka, T.; Koike, J.; Hemmi, H.; Koi, M.; Boland, C.R.; Goel, A. MicroRNA-200c modulates epithelial-tomesenchymal transition (EMT) in human colorectal cancer metastasis. Gut 2013, 62, 1315–1326. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Gaur, A.B.; Lengyel, E.; Peter, M.E. The miR-200 family determines the epithelial phenotype of cancer cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes Dev. 2008, 22, 894–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tudrej, P.; Olbryt, M.; Zembala-Nożyńska, E.; Kujawa, K.A.; Cortez, A.J.; Fiszer-Kierzkowska, A.; Pigłowski, W.; Nikiel, B.; Głowala-Kosińska, M.; Bartkowska-Chrobok, A.; et al. Establishment and characterization of the novel high-grade serous ovarian cancer cell line OVPA8. Int. J. Mol. Sci. 2018, 19, 2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaufort, C.M.; Helmijr, J.C.A.; Piskorz, A.M.; Hoogstraat, M.; Ruigrok-Ritstier, K.; Besselink, N.; Murtaza, M.; Van IJcken, W.F.J.; Heine, A.A.J.; Smid, M.; et al. Ovarian cancer cell line panel (OCCP): Clinical importance of in vitro morphological subtypes. PLoS ONE 2014, 9, e103988. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, J. MicroRNA-200c and microRNA-141 as potential diagnostic and prognostic biomarkers for ovarian cancer. Tumor Biol. 2015, 36, 4843–4850. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Su, X.; Zhang, X.; Zhang, Y.; Li, C.; Niu, W.; Liu, C.; Qu, K. miRNA-200a/c as potential biomarker in epithelial ovarian cancer (EOC): Evidence based on miRNA meta-signature and clinical investigations. Oncotarget 2016, 7, 81621–81633. [Google Scholar] [CrossRef] [Green Version]
- Bak, R.O.; Mikkelsen, J.G. miRNA sponges: Soaking up miRNAs for regulation of gene expression. Wiley Interdiscip. Rev. RNA 2014, 5, 317–333. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insert | Forward Primer with XbaI (5′ to 3′) | Reverse Primer with SbfI (5′ to 3′) | Size of Amplicon |
|---|---|---|---|
| DLC1 3′UTR | TAAGCATCTAGAGGCCACATGCCAGAATG | TGCTTACCTGCAGGCAGTATAGCAAATAATAAATTTATTAGGTG | 2582 bp |
| WT_A | TAAGCATCTAGAGGCCACATGCCAGAATG | TGCTTACCTGCAGGCACTGATATCCAAAATACTCAAATTTTAA | 1684 bp |
| WT_B | TAAGCATCTAGATTCCTCATGAAGATATACATGGA | TGCTTACCTGCAGGCAGTATAGCAAATAATAAATTTATTAGGTG | 900 bp |
| 3′–5′ | TAAGCATCTAGACAGTATAGCAAATAATAAATTTATTAGGTG | TGCTTACCTGCAGGGGCCACATGCCAGAATG | 2582 bp |
| Construct | Sequence | Size |
|---|---|---|
| WT_A (46bp) | 5′-AAGAGACAGTATTAGTAAA-3′ | 46 bp |
| MT_A (46bp) | 5′-ACTCAAGAGAGTCTATTAGTAA-3′ | 46 bp |
| WT_B (53bp) | 5′-GCGGCTTTCAGTATTTTGTACT-3′ | 53 bp |
| MT_B (53bp) | 5′-CGGCTTTGTCTATTTTGTAC-3′ | 53 bp |
| Characteristics | Cancer Samples (n = 29) | Normal Samples (n = 21) |
|---|---|---|
| Age (year) | ||
| Age (median) | 54 (24,80) | 55 (35,80) |
| ≤54 | 14 (48.23%) | 10 (47.62%) |
| >54 | 15 (51.72%) | 11 (52.38%) |
| Race | ||
| Malay | 18 (62.07%) | 11 (52.38%) |
| Chinese | 10 (34.48%) | 9 (42.86%) |
| Others | 1 (3.45%) | 1 (4.76%) |
| Stage | ||
| I | 11 (37.93%) | |
| II | 6 (20.69%) | |
| III | 8 (27.59%) | |
| IV | 4 (13.79%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ankasha, S.J.; Shafiee, M.N.; Abdul Wahab, N.; Raja Ali, R.A.; Mokhtar, N.M. Oncogenic Role of miR-200c-3p in High-Grade Serous Ovarian Cancer Progression via Targeting the 3′-Untranslated Region of DLC1. Int. J. Environ. Res. Public Health 2021, 18, 5741. https://doi.org/10.3390/ijerph18115741
Ankasha SJ, Shafiee MN, Abdul Wahab N, Raja Ali RA, Mokhtar NM. Oncogenic Role of miR-200c-3p in High-Grade Serous Ovarian Cancer Progression via Targeting the 3′-Untranslated Region of DLC1. International Journal of Environmental Research and Public Health. 2021; 18(11):5741. https://doi.org/10.3390/ijerph18115741
Chicago/Turabian StyleAnkasha, Sheril June, Mohamad Nasir Shafiee, Norhazlina Abdul Wahab, Raja Affendi Raja Ali, and Norfilza Mohd Mokhtar. 2021. "Oncogenic Role of miR-200c-3p in High-Grade Serous Ovarian Cancer Progression via Targeting the 3′-Untranslated Region of DLC1" International Journal of Environmental Research and Public Health 18, no. 11: 5741. https://doi.org/10.3390/ijerph18115741
APA StyleAnkasha, S. J., Shafiee, M. N., Abdul Wahab, N., Raja Ali, R. A., & Mokhtar, N. M. (2021). Oncogenic Role of miR-200c-3p in High-Grade Serous Ovarian Cancer Progression via Targeting the 3′-Untranslated Region of DLC1. International Journal of Environmental Research and Public Health, 18(11), 5741. https://doi.org/10.3390/ijerph18115741