
A Systematic Review of the Effects of Temperature on Anopheles Mosquito Development and Survival: Implications for Malaria Control in a Future Warmer Climate

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Methods
2.1. Eligibility Criteria
2.2. Search Strategy and Selection Criteria
2.3. Data Extraction
2.4. Risk of Bias Assessment
2.5. Data Analysis
3. Results
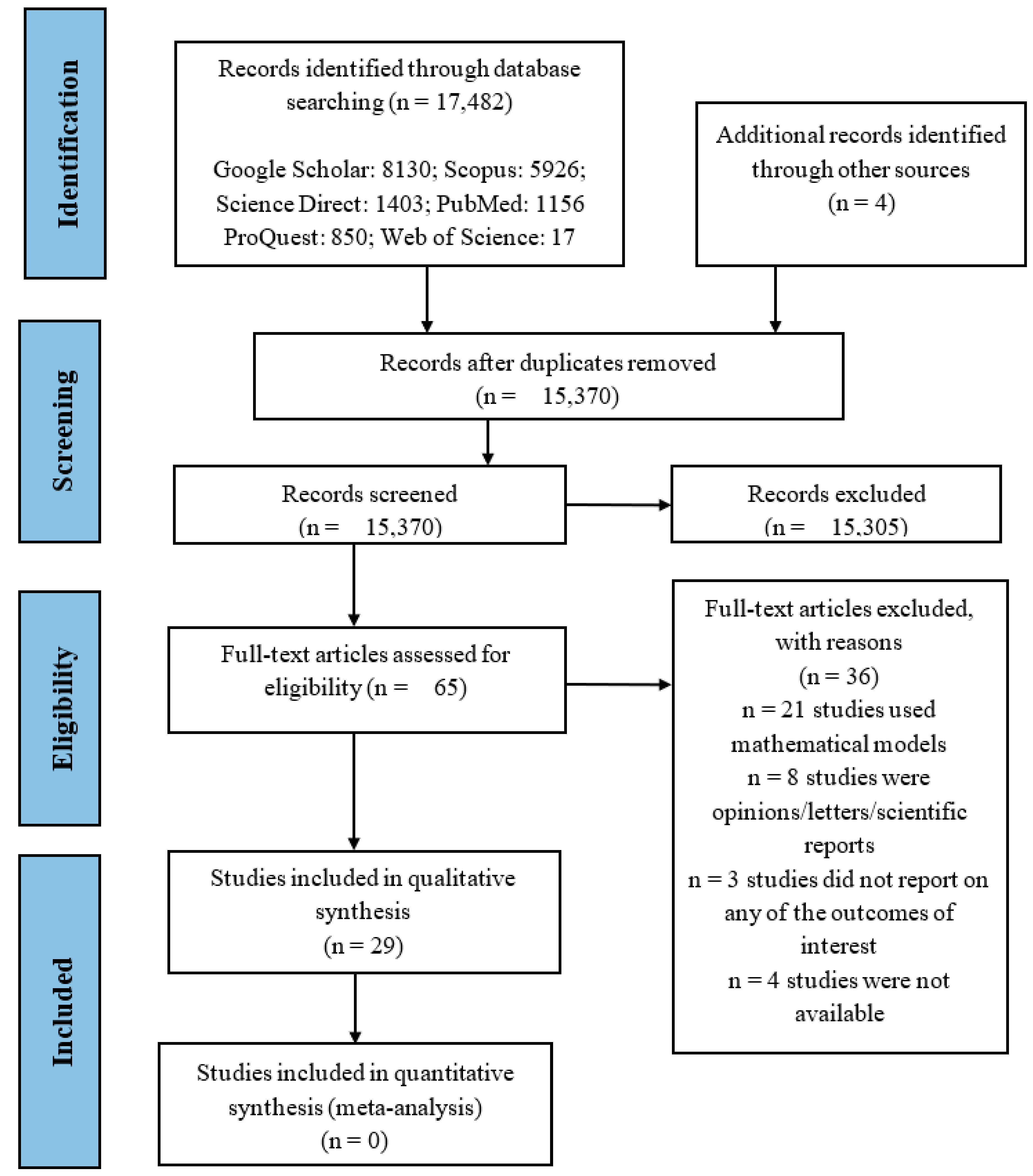
3.1. Search Results
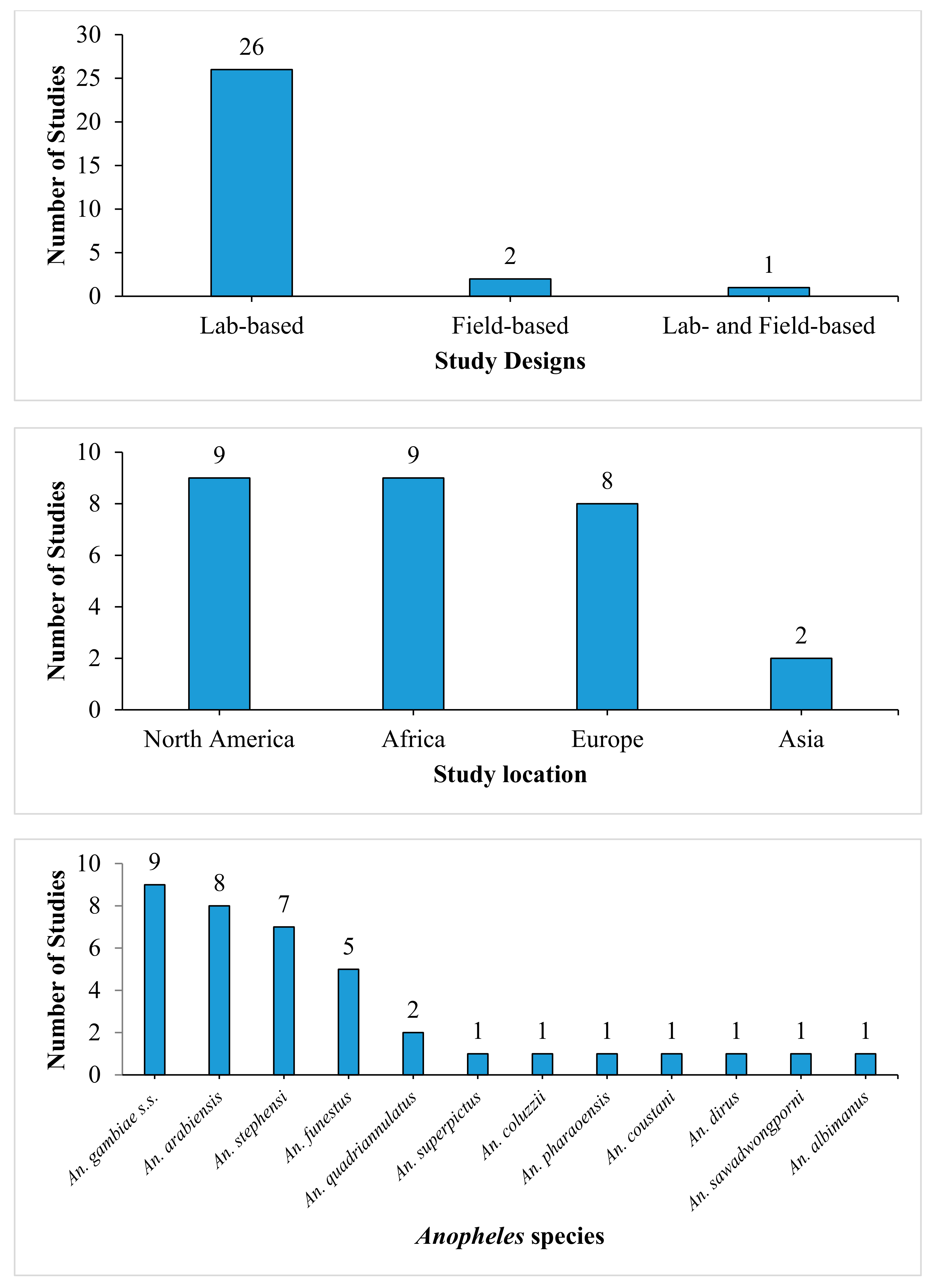
3.2. Study Characteristics
3.3. Risk of Bias Assessment
3.3.1. Selection Bias
3.3.2. Blinding (Performance and Detection Bias)
3.3.3. Randomization (Performance and Detection Bias)
3.3.4. Bias (Attrition and Reporting)
3.3.5. Other Sources of Bias (Funding Source and Rearing of Mosquitoes)
3.4. Effects of Temperature on the Immature stages of Anopheles Mosquitoes
3.5. Effects of Temperature on the Life History Traits of Adult Mosquitoes
3.5.1. Longevity
3.5.2. Body Size
3.5.3. Fecundity, Length of the Gonotrophic Cycle, and Biting Rate
3.6. Effects of Temperature on the Expression of Enzymes and Susceptibility to Insecticides
4. Discussion
4.1. Effects of Temperature on Immature Stages of Mosquitoes
4.2. Effects of Temperature on Adult Mosquitoes
4.2.1. Life-History Traits
4.2.2. Expression of Enzymes, Immune Responses, and Susceptibility to Insecticides
4.3. Implications of Findings for Malaria Control in a Future Warmer Climate
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DDT | Dichlorodiphenyltrichloroethane |
| EIP | Extrinsic incubation period |
| NOS | Nitric oxide synthase |
| PBO | Piperonyl butoxide |
| PRISMA | Preferred reporting items for systematic reviews and meta-analyses |
| ROS | Reactive oxygen species |
| SYRCLE | Systematic review center for laboratory animal experimentation |
| WHO | World health organization |
References
- Bulkeley, H. Cities and Climate Change; Routledge: London, UK, 2013. [Google Scholar]
- de Lima-Camara, T.N.; Honorio, N.A. Climate Change and Its Effect on Urban Mosquitoes in South America. Clim. Chang. Impacts Urban Pests 2016, 10, 127. [Google Scholar]
- Smith, E.K.; Mayer, A. A social trap for the climate? Collective action, trust and climate change risk perception in 35 countries. Glob. Environ. Chang. 2018, 49, 140–153. [Google Scholar] [CrossRef]
- Sánchez García, J.L.; Díez Sanz, J.M. Climate change, ethics and sustainability: An innovative approach. J. Innov. Knowl. 2018, 3, 70–75. [Google Scholar] [CrossRef]
- National Research Council. Advancing the Science of Climate Change; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Dantas-Torres, F. Climate change, biodiversity, ticks and tick-borne diseases: The butterfly effect. Int. J. Parasitol. Parasites Wildl. 2015, 4, 452–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ampadu, B.; Boateng, E.F.; Abassa, M.A. Assessing Adaptation Strategies to the Impacts of Climate Change: A Case Study of Pungu–Upper East Region, Ghana. Environ. Ecol. Res. 2018, 6, 33–44. [Google Scholar] [CrossRef]
- Antwi-Agyei, P.; Stringer, L.C.; Dougill, A.J. Livelihood adaptations to climate variability: Insights from farming households in Ghana. Reg. Environ. Chang. 2014, 14, 1615–1626. [Google Scholar] [CrossRef]
- McIntyre, K.M.; Setzkorn, C.; Hepworth, P.J.; Morand, S.; Morse, A.P.; Baylis, M. Systematic assessment of the climate sensitivity of important human and domestic animals pathogens in Europe. Sci. Rep. 2017, 7, 7134. [Google Scholar] [CrossRef] [Green Version]
- Elbers, A.; Koenraadt, C.; Meiswinkel, R. Mosquitoes and Culicoides biting midges: Vector range and the influence of climate change. Sci. Tech. Rev. Off. Int. Epizoot. 2015, 34, 123–137. [Google Scholar] [CrossRef] [Green Version]
- Ciota, A.T.; Matacchiero, A.C.; Kilpatrick, A.M.; Kramer, L.D. The effect of temperature on life history traits of Culex mosquitoes. J. Med. Entomol. 2014, 51, 55–62. [Google Scholar] [CrossRef]
- Guerra, C.; Howes, R.; Patil, A.; Gething, P.; Van Boeckel, T. The International Limits and Population at Risk of Plasmodium vivax transmission in 2009. PLoS Negl. Trop. Dis. 2010, 4, e774. [Google Scholar] [CrossRef]
- Hay, S.I.; Okiro, E.A.; Gething, P.W.; Patil, A.P.; Tatem, A.J.; Guerra, C.A.; Snow, R.W. Estimating the global clinical burden of Plasmodium falciparum malaria in 2007. PLoS Med. 2010, 7, e1000290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afrane, Y.A.; Githeko, A.K.; Yan, G. The ecology of Anopheles mosquitoes under climate change: Case studies from the effects of environmental changes in east Africa highlands. Ann. N. Y. Acad. Sci. 2012, 1249, 204. [Google Scholar] [CrossRef] [PubMed]
- Murdock, C.C.; Moller-Jacobs, L.L.; Thomas, M.B. Complex environmental drivers of immunity and resistance in malaria mosquitoes. Proc. R. Soc. B Biol. Sci. 2013, 280, 20132030. [Google Scholar] [CrossRef] [PubMed]
- Murdock, C.C.; Paaijmans, K.P.; Bell, A.S.; King, J.G.; Hillyer, J.F.; Read, A.F.; Thomas, M.B. Complex effects of temperature on mosquito immune function. Proc. R. Soc. B Biol. Sci. 2012, 279, 3357–3366. [Google Scholar] [CrossRef] [Green Version]
- Murdock, C.C.; Paaijmans, K.P.; Cox-Foster, D.; Read, A.F.; Thomas, M.B. Rethinking vector immunology: The role of environmental temperature in shaping resistance. Nat. Rev. Microbiol. 2012, 10, 869. [Google Scholar] [CrossRef] [Green Version]
- Glunt, K.D.; Oliver, S.V.; Hunt, R.H.; Paaijmans, K.P. The impact of temperature on insecticide toxicity against the malaria vectors Anopheles arabiensis and Anopheles funestus. Malar. J. 2018, 17, 131. [Google Scholar] [CrossRef] [Green Version]
- Glunt, K.D.; Paaijmans, K.P.; Read, A.F.; Thomas, M.B. Environmental temperatures significantly change the impact of insecticides measured using WHOPES protocols. Malar. J. 2014, 13, 350. [Google Scholar] [CrossRef] [Green Version]
- Oxborough, R.M.; N’Guessan, R.; Jones, R.; Kitau, J.; Ngufor, C.; Malone, D.; Rowland, M.W. The activity of the pyrrole insecticide chlorfenapyr in mosquito bioassay: Towards a more rational testing and screening of non-neurotoxic insecticides for malaria vector control. Malar. J. 2015, 14, 124. [Google Scholar] [CrossRef] [Green Version]
- Ezeakacha, N.F.; Yee, D.A. The role of temperature in affecting carry-over effects and larval competition in the globally invasive mosquito Aedes albopictus. Parasites Vectors 2019, 12, 123. [Google Scholar] [CrossRef] [Green Version]
- Carrington, L.B.; Armijos, M.V.; Lambrechts, L.; Barker, C.M.; Scott, T.W. Effects of fluctuating daily temperatures at critical thermal extremes on Aedes aegypti life-history traits. PLoS ONE 2013, 8, e58824. [Google Scholar] [CrossRef]
- Mohammed, A.; Chadee, D.D. Effects of different temperature regimens on the development of Aedes aegypti (L.) (Diptera: Culicidae) mosquitoes. Acta Trop. 2011, 119, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Mpho, M.; Callaghan, A.; Holloway, G.J. Temperature and genotypic effects on life history and fluctuating asymmetry in a field strain of Culex pipiens. Heredity 2002, 88, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Beck-Johnson, L.M.; Nelson, W.A.; Paaijmans, K.P.; Read, A.F.; Thomas, M.B.; Bjørnstad, O.N. The effect of temperature on Anopheles mosquito population dynamics and the potential for malaria transmission. PLoS ONE 2013, 8, e79276. [Google Scholar] [CrossRef] [PubMed]
- Christiansen-Jucht, C.D.; Parham, P.E.; Saddler, A.; Koella, J.C.; Basáñez, M.G. Temperature during larval development and adult maintenance influences the survival of Anopheles gambiae s.s. Parasites Vectors 2014, 7, 489. [Google Scholar] [CrossRef] [PubMed]
- Christiansen-Jucht, C.D.; Parham, P.E.; Saddler, A.; Koella, J.C.; Basáñez, M.G. Larval and adult environmental temperatures influence the adult reproductive traits of Anopheles gambiae s.s. Parasites Vectors 2015, 8, 456. [Google Scholar] [CrossRef] [Green Version]
- Lyons, C.L.; Coetzee, M.; Terblanche, J.S.; Chown, S.L. Thermal limits of wild and laboratory strains of two African malaria vector species, Anopheles arabiensis and Anopheles funestus. Malar. J. 2012, 11, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, S.V.; Brooke, B.D. The effect of elevated temperatures on the life history and insecticide resistance phenotype of the major malaria vector Anopheles arabiensis (Diptera: Culicidae). Malar. J. 2017, 16, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. J. Clin. Epidemiol. 2009, 62, e1–e34. [Google Scholar] [CrossRef] [Green Version]
- Hooijmans, C.R.; Rovers, M.M.; De Vries, R.B.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aytekin, S.; Murat Aytekin, A.; Alten, B. Effect of different larval rearing temperatures on the productivity (Ro) and morphology of the malaria vector Anopheles superpictus Grassi (Diptera: Culicidae) using geometric morphometrics. J. Vector Ecol. 2009, 34, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Barreaux, A.M.G.; Barreaux, P.; Thievent, K.; Koella, J.C. Larval environment influences vector competence of the malaria mosquito Anopheles gambiae. Malar. World J. 2016, 7, 1–6. [Google Scholar]
- Barreaux, A.M.G.; Stone, C.M.; Barreaux, P.; Koella, J.C. The relationship between size and longevity of the malaria vector Anopheles gambiae (s.s.) depends on the larval environment. Parasites Vectors 2018, 11, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayoh, M.N.; Lindsay, S.W. Effect of temperature on the development of the aquatic stages of Anopheles gambiae sensu stricto (Diptera: Culicidae). Bull. Entomol. Res. 2003, 93, 375–381. [Google Scholar] [CrossRef]
- Bayoh, M.N.; Lindsay, S.W. Temperature-related duration of aquatic stages of the Afrotropical malaria vector mosquito Anopheles gambiae in the laboratory. Med. Vet. Entomol. 2004, 18, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Charlwood, J.D.; Bragança, M. Some like it cool: The effect of ambient temperature on the size of Anopheles funestus from southern Mozambique. J. Med. Entomol. 2012, 49, 1154–1158. [Google Scholar] [CrossRef]
- Davies, C.; Coetzee, M.; Lyons, C.L. Effect of stable and fluctuating temperatures on the life history traits of Anopheles arabiensis and An. quadriannulatus under conditions of inter- and intra-specific competition. Parasites Vectors 2016, 9, 342. [Google Scholar] [CrossRef] [Green Version]
- Faiman, R.; Solon-Biet, S.; Sullivan, M.; Huestis, D.L.; Lehmann, T. The contribution of dietary restriction to extended longevity in the malaria vector Anopheles coluzzii. Parasites Vectors 2017, 10, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Impoinvil, D.E.; Cardenas, G.A.; Gihture, J.I.; Mbogo, C.M.; Beier, J.C. Constant temperature and time period effects on Anopheles gambiae egg hatching. J. Am. Mosq. Control Assoc. 2007, 23, 124. [Google Scholar] [CrossRef] [Green Version]
- Kirby, M.J.; Lindsay, S.W. Effect of temperature and inter-specific competition on the development and survival of Anopheles gambiae sensu stricto and An. arabiensis larvae. Acta Trop. 2009, 109, 118–123. [Google Scholar] [CrossRef]
- Lyons, C.L.; Coetzee, M.; Chown, S.L. Stable and fluctuating temperature effects on the development rate and survival of two malaria vectors, Anopheles arabiensis and Anopheles funestus. Parasites Vectors 2013, 6, 104. [Google Scholar] [CrossRef] [Green Version]
- Mala, A.O.; Irungu, L.W.; Mitaki, E.K.; Shililu, J.I.; Mbogo, C.M.; Njagi, J.K.; Githure, J.I. Gonotrophic cycle duration, fecundity and parity of Anopheles gambiae complex mosquitoes during an extended period of dry weather in a semi arid area in Baringo County, Kenya. Int. J. Mosq. Res. 2014, 1, 28–34. [Google Scholar]
- Mamai, W.; Lobb, L.N.; Bimbilé Somda, N.S.; Maiga, H.; Yamada, H.; Lees, R.S.; Bouyer, J.; Gilles, J.R.L. Optimization of mass-rearing methods for Anopheles arabiensis larval stages: Effects of rearing water temperature and larval density on mosquito life-history traits. J. Econ. Entomol. 2018, 111, 2383–2390. [Google Scholar] [CrossRef]
- Murdock, C.C.; Blanford, S.; Luckhart, S.; Thomas, M.B. Ambient temperature and dietary supplementation interact to shape mosquito vector competence for malaria. J. Insect Physiol. 2014, 67, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Olayemi, I.; Danlami, G.; Isah, B.; Odeyemi, O.; Ukubuiwe, A.; Mustapha, O.M. Indoor behaviour responses of the principal malaria vector, Anopheles gambiae (Diptera: Culicidae), in relation to micro-climatic conditions in Minna, North Central Nigeria. Res. J. Parasitol. 2011, 6, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Paaijmans, K.P.; Cator, L.J.; Thomas, M.B. Temperature-dependent pre-bloodmeal period and temperature-driven asynchrony between parasite development and mosquito biting rate reduce malaria transmission intensity. PLoS ONE 2013, 8, e55777. [Google Scholar] [CrossRef] [PubMed]
- Paaijmans, K.P.; Heinig, R.L.; Seliga, R.A.; Blanford, J.I.; Blanford, S.; Murdock, C.C.; Thomas, M.B. Temperature variation makes ectotherms more sensitive to climate change. Glob. Chang. Biol. 2013, 19, 2373–2380. [Google Scholar] [CrossRef] [Green Version]
- Phasomkusolsil, S.; Lerdthusnee, K.; Khuntirat, B.; Kongtak, W.; Pantuwatana, K.; Murphy, J.R. Effect of temperature on laboratory reared Anopheles dirus Peyton and Harrison and Anopheles sawadwongporni Rattanarithikul and Green. Southeast Asian J. Trop. Med. Public Health 2011, 42, 63–70. [Google Scholar]
- Shapiro, L.L.M.; Whitehead, S.A.; Thomas, M.B. Quantifying the effects of temperature on mosquito and parasite traits that determine the transmission potential of human malaria. PLoS Biol. 2017, 15, e2003489. [Google Scholar] [CrossRef] [Green Version]
- Rúa, G.L.; Quiñones, M.L.; Vélez, I.D.; Zuluaga, J.S.; Rojas, W.; Poveda, G.; Ruiz, D. Laboratory estimation of the effects of increasing temperatures on the duration of gonotrophic cycle of Anopheles albimanus (Diptera: Culicidae). Mem. Inst. Oswaldo Cruz 2005, 100, 515–520. [Google Scholar] [CrossRef]
- Wallace, J.R.; Merritt, R.W. Influence of microclimate, food, and predation on Anopheles quadrimaculatus (Diptera: Culicidae) growth and development rates, survivorship, and adult size in a Michigan pond. Environ. Entomol. 1999, 28, 233–239. [Google Scholar] [CrossRef]
- Green, B.S.; McCormick, M.I. Maternal and paternal effects determine size, growth and performance in larvae of a tropical reef fish. Mar. Ecol. Prog. Ser. 2005, 289, 263–272. [Google Scholar] [CrossRef]
- McCormick, M.; Gagliano, M. Carry-over effects-the importance of a good start. In Proceedings of the 11 th International Coral Reef Symposium, Ft. Lauderdale, FL, USA, 7–11 July 2008. [Google Scholar]
- Ukubuiwe, A.C.; Olayemi, I.K.; Arimoro, F.O.; Omalu, I.C.J.; Baba, B.M.; Ukubuiwe, C.C.; Odeyemi, M.O.; Adeniyi, K.A. Influence of rearing-water temperature on life stages’ vector attributes, distribution and utilisation of metabolic reserves in Culex quinquefasciatus (Diptera: Culicidae): Implications for disease transmission and vector control. J. Basic Appl. Zool. 2018, 79, 32. [Google Scholar] [CrossRef] [Green Version]
- Protopopoff, N.; Van Bortel, W.; Speybroeck, N.; Van Geertruyden, J.-P.; Baza, D.; D’Alessandro, U.; Coosemans, M. Ranking Malaria Risk Factors to Guide Malaria Control Efforts in African Highlands. PLoS ONE 2009, 4, e8022. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Huey, R.B. Size, temperature, and fitness: Three rules. Evol. Ecol. Res. 2008, 10, 251–268. [Google Scholar]
- Amer, K.; Saavedra-Rodriguez, K.; Black, W.C.; Gray, E.M. Effect of selection for pyrethroid resistance on abiotic stress tolerance in Aedes aegypti from Merida, Yucatan, Mexico. Insects 2021, 12, 124. [Google Scholar] [CrossRef]
- Shah, A.A.; Woods, H.A.; Havird, J.C.; Encalada, A.C.; Flecker, A.S.; Funk, W.C.; Guayasamin, J.M.; Kondratieff, B.C.; Poff, N.L.; Thomas, S.A.; et al. Temperature-dependence of metabolic rate in tropical and temperate aquatic insects: Support for the climate variability hypothesis in mayflies but not stoneflies. Glob. Chang. Biol. 2020, 27, 297–311. [Google Scholar] [CrossRef]
- Pörtner, H.-O.; Bock, C.; Mark, F.C. Oxygen- and capacity-limited thermal tolerance: Bridging ecology and physiology. J. Exp. Biol. 2017, 220, 2685–2696. [Google Scholar] [CrossRef] [Green Version]
- Dodson, B.L.; Kramer, L.D.; Rasgon, J.L. Effects of larval rearing temperature on immature development and West Nile virus vector competence of Culex tarsalis. Parasites Vectors 2012, 5, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Churcher, T.S.; Bousema, T.; Walker, M.; Drakeley, C.; Schneider, P.; Ouédraogo, A.L.; Basáñez, M.G. Predicting mosquito infection from Plasmodium falciparum gametocyte density and estimating the reservoir of infection. eLife 2013, 2, e00626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawes, E.J.; Churcher, T.S.; Zhuang, S.; Sinden, R.E.; Basáñez, M.-G. Anopheles mortality is both age- and Plasmodium-density dependent: Implications for malaria transmission. Malar. J. 2009, 8, 228. [Google Scholar] [CrossRef] [Green Version]
- Rajatileka, S.; Burhani, J.; Ranson, H. Mosquito age and susceptibility to insecticides. Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Bhuju, G.; Phaijoo, G.R.; Gurung, D.B. Mathematical study on impact of temperature in malaria disease transmission dynamics. Adv. Comput. Sci. 2018, 1, 107. [Google Scholar] [CrossRef]
- Marinho, R.A.; Beserra, E.B.; Bezerra-Gusmão, M.A.; Porto, V.d.S.; Olinda, R.A.; dos Santos, C.A. Effects of temperature on the life cycle, expansion, and dispersion of Aedes aegypti (Diptera: Culicidae) in three cities in Paraiba, Brazil. J. Vector Ecol. 2016, 41, 1–10. [Google Scholar] [CrossRef]
- Swain, V.; Seth, R.K.; Mohanty, S.S.; Raghavendra, K. Effect of temperature on development, eclosion, longevity and survivorship of malathion-resistant and malathion-susceptible strain of Culex quinquefasciatus. Parasitol. Res. 2008, 103, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Onyango, M.G.; Bialosuknia, S.M.; Payne, A.F.; Mathias, N.; Kuo, L.; Vigneron, A.; DeGennaro, M.; Ciota, A.T.; Kramer, L.D. Increased temperatures reduce the vectorial capacity of Aedes mosquitoes for Zika virus. Emerg. Microbes Infect. 2020, 9, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Keil, G.; Cummings, E.; De Magalhães, J.P. Being cool: How body temperature influences ageing and longevity. Biogerontology 2015, 16, 383–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, T.; Dalton, R.; Kim, E.H.; Dahl, E.; Diabate, A.; Dabire, R.; Dujardin, J.P. Genetic contribution to variation in larval development time, adult size, and longevity of starved adults of Anopheles gambiae. Infect. Genet. Evol. 2006, 6, 410–416. [Google Scholar] [CrossRef]
- Yeap, H.L.; Endersby, N.M.; Johnson, P.H.; Ritchie, S.A.; Hoffmann, A.A. Body size and wing shape measurements as quality indicators of Aedes aegypti mosquitoes destined for field release. Am. J. Trop. Med. Hyg. 2013, 89, 78–92. [Google Scholar] [CrossRef]
- Afrane, Y.A.; Zhou, G.; Lawson, B.W.; Githeko, A.K.; Yan, G. Effects of microclimatic changes caused by deforestation on the survivorship and reproductive fitness of Anopheles gambiae in Western Kenya highlands. Am. J. Trop. Med. Hyg. 2006, 74, 772–778. [Google Scholar] [CrossRef]
- Lardeux, F.J.; Tejerina, R.H.; Quispe, V.; Chavez, T.K. A physiological time analysis of the duration of the gonotrophic cycle of Anopheles pseudopunctipennis and its implications for malaria transmission in Bolivia. Malar. J. 2008, 7, 141. [Google Scholar] [CrossRef] [Green Version]
- Sy, V.E.; Agnew, P.; Sidobre, C.; Michalakis, Y. Reduced survival and reproductive success generates selection pressure for the dengue mosquito Aedes aegypti to evolve resistance against infection by the microsporidian parasite Vavraia culicis. Evol. Appl. 2014, 7, 468–479. [Google Scholar] [CrossRef]
- Nature Education. Dengue Transmission. 2014. Available online: https://www.nature.com/scitable/topicpage/dengue-transmission-22399758/ (accessed on 14 June 2021).
- Shaw, W.R.; Holmdahl, I.E.; Itoe, M.A.; Werling, K.; Marquette, M.; Paton, D.G. Multiple blood feeding in mosquitoes shortens the Plasmodium falciparum incubation period and increases malaria transmission potential. PLoS Pathog. 2021, 16, e1009131. [Google Scholar]
- Suwanchaichinda, C.; Paskewitz, S.M. Effects of Larval Nutrition, Adult Body Size, and Adult Temperature on the Ability of Anopheles gambiae (Diptera: Culicidae) to Melanize Sephadex Beads. J. Med. Entomol. 1998, 35, 157–161. [Google Scholar] [CrossRef]
- Kristan, M.; Abeku, T.A.; Lines, J. Effect of environmental variables and kdr resistance genotype on survival probability and infection rates in Anopheles gambiae (ss). Parasites Vectors 2018, 11, 560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbel, V.; N’Guessan, R. Distribution, mechanisms, impact and management of insecticide resistance in malaria vectors: A pragmatic review. In Anopheles Mosquitoes-New Insights into Malaria Vectors; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Liu, N. Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Matowo, J.; Kulkarni, M.A.; Mosha, F.W.; Oxborough, R.M.; Kitau, J.A.; Tenu, F.; Rowland, M. Biochemical basis of permethrin resistance in Anopheles arabiensis from Lower Moshi, north-eastern Tanzania. Malar. J. 2010, 9, 193. [Google Scholar] [CrossRef] [Green Version]
- Ranson, H.; N’Guessan, R.; Lines, J.; Moiroux, N.; Nkuni, Z.; Corbel, V. Pyrethroid resistance in African anopheline mosquitoes: What are the implications for malaria control? Trends Parasitol. 2011, 27, 91–98. [Google Scholar] [CrossRef]
- Ochomo, E.; Bayoh, M.N.; Brogdon, W.G.; Gimnig, J.E.; Ouma, C.; Vulule, J.M.; Walker, E.D. Pyrethroid resistance in Anopheles gambiae ss and Anopheles arabiensis in western Kenya: Phenotypic, metabolic and target site characterizations of three populations. Med. Vet. Entomol. 2013, 27, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Labbé, P.; Alout, H.; Djogbénou, L.; Pasteur, N.; Weill, M. Evolution of Resistance to Insecticide in Disease Vectors. In Genetics and Evolution of Infectious Disease; Tibayrenc, M., Ed.; Elsevier: London, UK, 2011; pp. 363–409. [Google Scholar]
- Matzrafi, M. Climate change exacerbates pest damage through reduced pesticide efficacy. Pest Manag. Sci. 2019, 75, 9–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casida, J.E.; Durkin, K.A. Neuroactive insecticides: Targets, selectivity, resistance, and secondary effects. Annu. Rev. Entomol. 2013, 58, 99–117. [Google Scholar] [CrossRef]
- Hodjati, M.; Curtis, C. Effects of permethrin at different temperatures on pyrethroid-resistant and susceptible strains of Anopheles. Med. Vet. Entomol. 1999, 13, 415–422. [Google Scholar] [CrossRef]
- Khan, H.A.A.; Akram, W. The effect of temperature on the toxicity of insecticides against Musca domestica L.: Implications for the effective management of Diarrhea. PLoS ONE 2014, 9, e95636. [Google Scholar]
- Amarasekare, K.G.; Edelson, J.V. Effect of temperature on efficacy of insecticides to differential grasshopper (Orthoptera: Acrididae). J. Econ. Entomol. 2004, 97, 1595–1602. [Google Scholar] [CrossRef]
- Ngarakana-Gwasira, E.; Bhunu, C.; Masocha, M.; Mashonjowa, E. Assessing the role of climate change in malaria transmission in Africa. Malar. Res. Treat. 2016, 2016, 7104291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshen, M.B.; Morse, A.P. A weather-driven model of malaria transmission. Malar. J. 2004, 3, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mordecai, E.A.; Paaijmans, K.P.; Johnson, L.R.; Balzer, C.; Ben-Horin, T.; de Moor, E.; McNally, A.; Pawar, S.; Ryan, S.J.; Smith, T.C.; et al. Optimal temperature for malaria transmission is dramatically lower than previously predicted. Ecol. Lett. 2013, 16, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Stresman, G.H. Beyond temperature and precipitation: Ecological risk factors that modify malaria transmission. Acta Trop. 2010, 116, 167–172. [Google Scholar] [CrossRef]
- Ikemoto, T. Tropical malaria does not mean hot environments. J. Med. Entomol. 2008, 45, 963–969. [Google Scholar] [CrossRef]
- Lefevre, T.; Ohm, J.; Dabiré, K.R.; Cohuet, A.; Choisy, M.; Thomas, M.B.; Cator, L. Transmission traits of malaria parasites within the mosquito: Genetic variation, phenotypic plasticity, and consequences for control. Evol. Appl. 2018, 11, 456–469. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Author/Year | Sequence Generation (Selection Bias) | Baseline Characteristics (Selection Bias) | Allocation Concealment (Selection Bias) | Incomplete Outcome Data (Attrition Bias) | Selective Reporting (Reporting Bias) | Other Bias (Rearing of Mosquito) | Other Bias (Funding Source) |
|---|---|---|---|---|---|---|---|
| Aytekin et al. [32] | High risk | Low risk | Unclear risk | Low risk | Low risk | Low risk | High risk |
| Barreaux et al. [33] | High risk | Low risk | Unclear risk | Low risk | Low risk | Low risk | High risk |
| Barreaux et al. [34] | High risk | Low risk | Unclear risk | Low risk | Low risk | Low risk | Low risk |
| Bayoh and Lindsay [35] | High risk | Low risk | Unclear risk | Low risk | Low risk | Low risk | Low risk |
| Bayoh and Lindsay [36] | High risk | Low risk | Unclear risk | Low risk | Low risk | Low risk | Low risk |
| Charlwood and Bragança [37] | High risk | Unclear risk | Unclear risk | Low risk | Low risk | High risk | High risk |
| Christiansen-Jucht et al. [26] | High risk | Low risk | Unclear risk | Low risk | Low risk | Low risk | Low risk |
| Christiansen-Jucht et al. [27] | High risk | Low risk | Unclear risk | Low risk | Low risk | Low risk | Low risk |
| Davies et al. [38] | High risk | Low risk | Unclear risk | Low risk | Low risk | Low risk | Low risk |
| Faiman et al. [39] | High risk | Low risk | Unclear risk | Low risk | Low risk | Unclear risk | Low risk |
| Glunt et al. [18] | High risk | Low risk | High risk | Low risk | Low risk | High risk | Low risk |
| Glunt et al. [19] | High risk | Low risk | High risk | Low risk | Low risk | High risk | Low risk |
| Impoinvil et al. [40] | High risk | Low risk | Unclear risk | Low risk | Low risk | Low risk | Low risk |
| Kirby and Lindsay [41] | High risk | Low risk | Unclear risk | Low risk | Low risk | Low risk | High risk |
| Lyons et al. [42] | High risk | Low risk | High risk | Low risk | Low risk | Low risk | Low risk |
| Lyons et al. [28] | High risk | Low risk | High risk | Low risk | Low risk | High risk | Low risk |
| Mala et al. [43] | Low risk | Low risk | High risk | Low risk | Low risk | High risk | Low risk |
| Mamai et al. [44] | High risk | Low risk | High risk | Low risk | Low risk | Low risk | Low risk |
| Murdock et al. [45] | High risk | Low risk | High risk | Low risk | Low risk | High risk | Low risk |
| Murdock et al. [15] | High risk | Low risk | High risk | Low risk | Low risk | High risk | Low risk |
| Murdock et al. [16] | High risk | Low risk | High risk | Low risk | Low risk | High risk | Low risk |
| Olayemi et al. [46] | High risk | Unclear risk | High risk | Low risk | Low risk | High risk | High risk |
| Oliver and Brooke [29] | High risk | Low risk | High risk | Low risk | Low risk | Low risk | Low risk |
| Paaijmans et al. [47] | High risk | Low risk | High risk | Low risk | Low risk | Low risk | High risk |
| Paaijmans et al. [48] | High risk | Low risk | High risk | Low risk | Low risk | Low risk | Low risk |
| Phasomkusolsil et al. [49] | High risk | Low risk | High risk | Low risk | Low risk | Low risk | High risk |
| Rúa et al. [51] | High risk | Low risk | High risk | Low risk | Low risk | Low risk | High risk |
| Shapiro et al. [50] | High risk | Low risk | High risk | Low risk | Low risk | Low risk | Unclear risk |
| Wallace and Merritt [52] | High risk | Low risk | High risk | Low risk | Low risk | Low risk | Low risk |
| Author, Year | Study Type | Study Location | Species Considered | Conditions | Outcome Considered |
|---|---|---|---|---|---|
| Christiansen-Jucht et al. [27] | Laboratory-based | United Kingdom | An. gambiae s.s. | 23, 27, 31, and 35 ± 1 °C 12:12 (L:D) photoperiod RH 75% ± 5% |
|
| Davies et al. [38] | Laboratory-based | South Africa | An. arabiensis An. quadriannulatus | 25, 20–30, and 18–35 °C 12:12 (L:D) photoperiod RH 80% |
|
| Impoinvil et al. [40] | Laboratory-based | Kenya | An. gambiae s.s. | Immature: 30–35 °C Adult: 22–27 °C RH 80–90% |
|
| Mamai et al. [44] | Laboratory-based | Austria | An. arabiensis | 22 ± 1 °C, 22–27 ± 1 °C, 27 ± 1 °C 12:12 (L:D) photoperiod RH 80% |
|
| Phasomkusolsil et al. [49] | Laboratory-based | Thailand | An. dirus An. sawadwongporni | 23 and 30 °C |
|
| Aytekin et al. [32] | Laboratory-based | Turkey | An. superpictus | 15, 20, 25, 27, 30, and 35 °C, 12:12 (L:D) photoperiod RH 65% ± 5% |
|
| Bayoh and Lindsay [35] | Laboratory-based | United Kingdom | An. gambiae s.s. | 10 to 40 °C (±1 °C), with 2 °C increments 12:12 (L:D) photoperiod RH 80% ± 10% |
|
| Kirby and Lindsay [41] | Laboratory-based | United Kingdom | An. gambiae s.s. An. arabiensis | 25, 30, or 35 ◦C |
|
| Lyons et al. [42] | Laboratory-based | South Africa | An. arabiensis An. funestus | 15, 18, 20, 22, 25, 28, 30, 32 35, 15 °C–35, and 20–30 °C 12:12 (L:D) photoperiod RH 80% |
|
| Oliver and Brooke [29] | Laboratory-based | South Africa | An. arabiensis | 25, 30, and 35 °C RH 80% ± 5% |
|
| Paaijmans et al. [48] | Laboratory-based | United States of America | An. stephensi | 16 to 36 °C, with 2 °C increments |
|
| Wallace and Merritt [52] | Field and Laboratory-based | United States of America | An. quadrimaculatus | 18, 23, and 28 °C |
|
| Lyons et al. [28] | Laboratory-based | South Africa | An. funestus An. arabiensis | 20, 25, and 30 °C 12:12 (L:D) photoperiod RH 80% |
|
| Bayoh and Lindsay [36] | Laboratory-based | United Kingdom | An. gambiae s.s. | 10 to 40 °C (±1 °C), with 2 °C increments 12:12 (L:D) photoperiod RH 80 ± 10% |
|
| Christiansen-Jucht et al. [26] | Laboratory-based | United Kingdom | An. gambiae s.s. | 23, 27, 31, and 35 ± 1 °C 12:12 (L:D) photoperiod RH 75% ± 5% |
|
| Barreaux et al. [34] | Laboratory-based | Switzerland | An. gambiae s.s. | 21, 25, and 29 °C 12:12 (L:D) photoperiod RH 70% ± 5% |
|
| Author, Year | Study Type | Study Location | Species Considered | Conditions | Outcome Considered |
|---|---|---|---|---|---|
| Aytekin et al. [32] | Laboratory-based | Turkey | An. superpictus | 15, 20, 25, 27, 30, and 35 °C, 12:12 (L:D) photoperiod RH 65% ± 5% |
|
| Barreaux et al. [34] | Laboratory-based | Switzerland | An. gambiae s.s. | 21, 25, and 29 °C 12:12 (L:D) photoperiod RH 70% ± 5% |
|
| Faiman et al. [39] | Laboratory-based | United States of America | An. coluzzii | 22, 23.5, and 27 °C, 2:12 or 11:13 L:D photoperiod RH 85% and 50% |
|
| Olayemi et al. [46] | Field and Laboratory-based | Nigeria | An. gambiae | Seasons Dry: 31.12 ± 1.09 °C, RH 44.01 ± 7.02% Rainy: 27.67 ± 1.27 °C, RH 69.51% ± 12.44% |
|
| Oliver and Brooke [29] | Laboratory-based | South Africa | An. arabiensis | 25, 30, and 35 °C RH 80% ± 5% |
|
| Author, Year | Study Type | Study Location | Species Considered | Conditions | Outcome Considered |
|---|---|---|---|---|---|
| Aytekin et al. [32] | Laboratory-based | Turkey | An. superpictus | 15, 20, 25, 27, 30, and 35 °C, 12:12 (L:D) photoperiod RH 65% ± 5% |
|
| Barreaux et al. [33] | Laboratory-based | Switzerland | An. gambiae s.s. | 21 °C, 25 °C, and 29 °C |
|
| Barreaux et al. [34] | Laboratory-based | Switzerland | An. gambiae s.s. | 21, 25, and 29 °C 12:12 (L:D) photoperiod RH 70% ± 5% |
|
| Charlwood and Bragança [37] | Field-based | Mozambique | An. funestus | 17 to 33 °C |
|
| Christiansen-Jucht et al. [27] | Laboratory-based | United Kingdom | An. gambiae s.s. | 23, 27, 31, and 35 ± 1°C 12:12 (L:D) photoperiod RH 75% ± 5% |
|
| Kirby and Lindsay [41] | Laboratory-based | United Kingdom | An. gambiae s.s. An. arabiensis | 25, 30 or 35 °C |
|
| Phasomkusolsil et al. [49] | Laboratory-based | Thailand | An. dirus An. sawadwongporni | 23 and 30 °C |
|
| Author, Year | Study Type | Study Location | Species Considered | Conditions | Outcome Considered |
|---|---|---|---|---|---|
| Aytekin et al. [32] | Laboratory-based | Turkey | An. superpictus | 15, 20, 25, 27, 30, and 35 °C, 12:12 (L:D) photoperiod RH 65% ± 5% |
|
| Christiansen-Jucht et al. [27] | Laboratory-based | United Kingdom | An. gambiae s.s. | 23, 27, 31, and 35 ± 1 °C 12:12 (L:D) photoperiod RH 75% ± 5% |
|
| Phasomkusolsil et al. [49] | Laboratory-based | Thailand | An. dirus An. sawadwongporni | 23 and 30 °C |
|
| Mala et al. [43] | Field-based | Kenya | An. arabiensis An. pharaoensis An. coustani An. funestus | Indoor Temperature Dry season (28.22 ± 1.1 °C) Rainy season (27.12 ± 1.2 °C) Outdoor Temperature Dry season (26.32 ± 0.33 °C) Rainy season (24.82 ± 0.33 °C) |
|
| Paaijmans et al. [47] | Laboratory-based | United States of America | An. stephensi | 22, 24, and 26 °C 12:12 (L:D) photoperiod RH 90% ± 5% |
|
| Rúa et al. [51] | Laboratory-based | An. albimanus | 24, 27, and 30 °C |
| |
| Shapiro et al. [50] | Laboratory-based | United States of America | An. stephensi | 21, 24, 27, 30, 32, and 34 °C |
|
| Author, Year | Study Type | Study Location | Species Considered | Conditions | Outcome Considered |
|---|---|---|---|---|---|
| Glunt et al. [18] | Laboratory-based | South Africa | An. funestus An. arabiensis | 18 °C, 25 °C, and 30 °C RH 70% for 18 °C and 30 °C RH 80% for 25 °C |
|
| Glunt et al. [19] | Laboratory-based | United States of America | An. stephensi | 12, 18, 22, and 26 °C |
|
| Oliver and Brooke [29] | Laboratory-based | South Africa | An. arabiensis | 25, 30, and 35 °C RH 80% ± 5% |
|
| Murdock et al. [45] | Laboratory-based | United States of America | An. stephensi | 20, 22, 24, 26, and 28 ± 0.5 °C 12:12 (L:D) photoperiod RH 80% ± 5% |
|
| Murdock et al. [15] | Laboratory-based | United States of America | An. stephensi | 16, 26, 32 ± 0.5 °C; 16, 26, 32 ± 6 °C 12:12 (L:D) photoperiod RH 80% ± 5% |
|
| Murdock et al. [16] | Laboratory-based | United States of America | An. stephensi | 12, 18, 24, 28, and 34 + 0.5 °C 12:12 (L:D) photoperiod RH 80% ± 5% |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agyekum, T.P.; Botwe, P.K.; Arko-Mensah, J.; Issah, I.; Acquah, A.A.; Hogarh, J.N.; Dwomoh, D.; Robins, T.G.; Fobil, J.N. A Systematic Review of the Effects of Temperature on Anopheles Mosquito Development and Survival: Implications for Malaria Control in a Future Warmer Climate. Int. J. Environ. Res. Public Health 2021, 18, 7255. https://doi.org/10.3390/ijerph18147255
Agyekum TP, Botwe PK, Arko-Mensah J, Issah I, Acquah AA, Hogarh JN, Dwomoh D, Robins TG, Fobil JN. A Systematic Review of the Effects of Temperature on Anopheles Mosquito Development and Survival: Implications for Malaria Control in a Future Warmer Climate. International Journal of Environmental Research and Public Health. 2021; 18(14):7255. https://doi.org/10.3390/ijerph18147255
Chicago/Turabian StyleAgyekum, Thomas P., Paul K. Botwe, John Arko-Mensah, Ibrahim Issah, Augustine A. Acquah, Jonathan N. Hogarh, Duah Dwomoh, Thomas G. Robins, and Julius N. Fobil. 2021. "A Systematic Review of the Effects of Temperature on Anopheles Mosquito Development and Survival: Implications for Malaria Control in a Future Warmer Climate" International Journal of Environmental Research and Public Health 18, no. 14: 7255. https://doi.org/10.3390/ijerph18147255
APA StyleAgyekum, T. P., Botwe, P. K., Arko-Mensah, J., Issah, I., Acquah, A. A., Hogarh, J. N., Dwomoh, D., Robins, T. G., & Fobil, J. N. (2021). A Systematic Review of the Effects of Temperature on Anopheles Mosquito Development and Survival: Implications for Malaria Control in a Future Warmer Climate. International Journal of Environmental Research and Public Health, 18(14), 7255. https://doi.org/10.3390/ijerph18147255