
SH-29 and SK-119 Attenuates Air-Pollution Induced Damage by Activating Nrf2 in HaCaT Cells

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
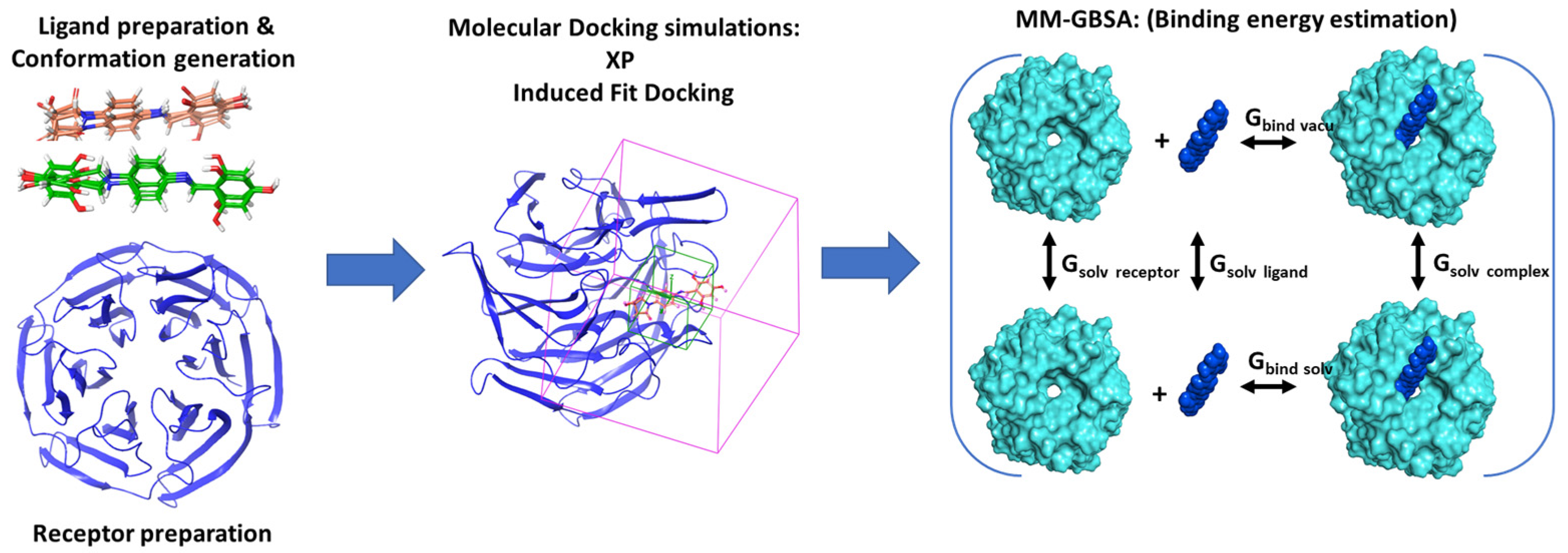
2.1. Computational Studies
2.1.1. Ligand Preparation
2.1.2. Conformational Search
2.1.3. Protein Preparations and Receptor Grid Generation
2.1.4. Molecular Docking
Glide
Induced Fit Docking
2.1.5. Binding Free Energy Analysis for Drug-Target Binding Energy Estimation
2.1.6. HOMO LUMO Stability Analysis of the Screened Compounds
2.2. Synthetic Chemistry
2.3. Experimental Biology
2.3.1. Cell Culture and Ex Vivo Skin Cultures
2.3.2. Cell and Tissue Viability Assay
2.3.3. Nrf2-Activation Assay
2.3.4. In Vitro Antioxidant Activity
2.3.5. Intracellular and Mitochondrial Reactive Oxygen Species (ROS) Determination
2.3.6. IL-8 and IL-1α Quantification
2.3.7. Histology
2.4. Statistical Analysis
3. Results
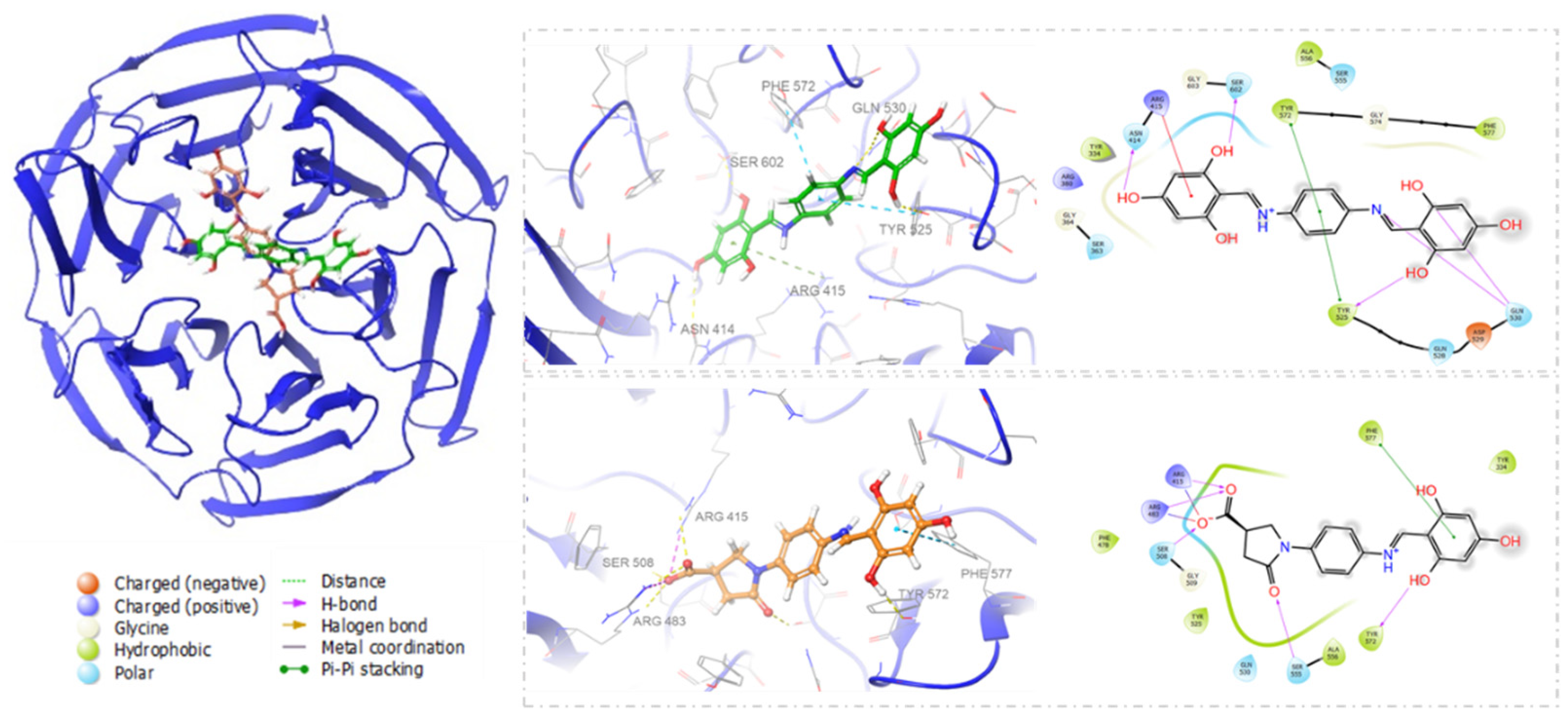
3.1. Molecular Docking
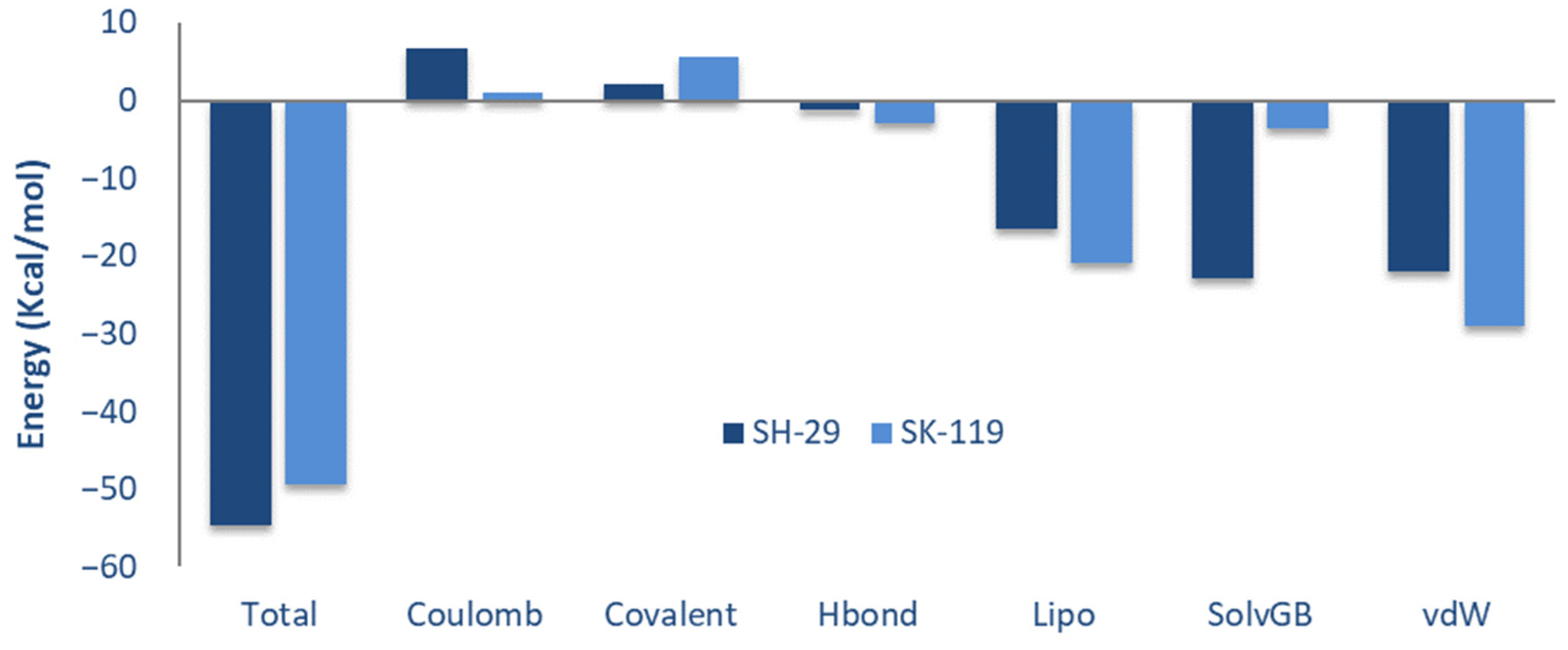
3.2. Calculation for Prime Molecular Mechanics/Generalized Born Surface Area (MM-GBSA)
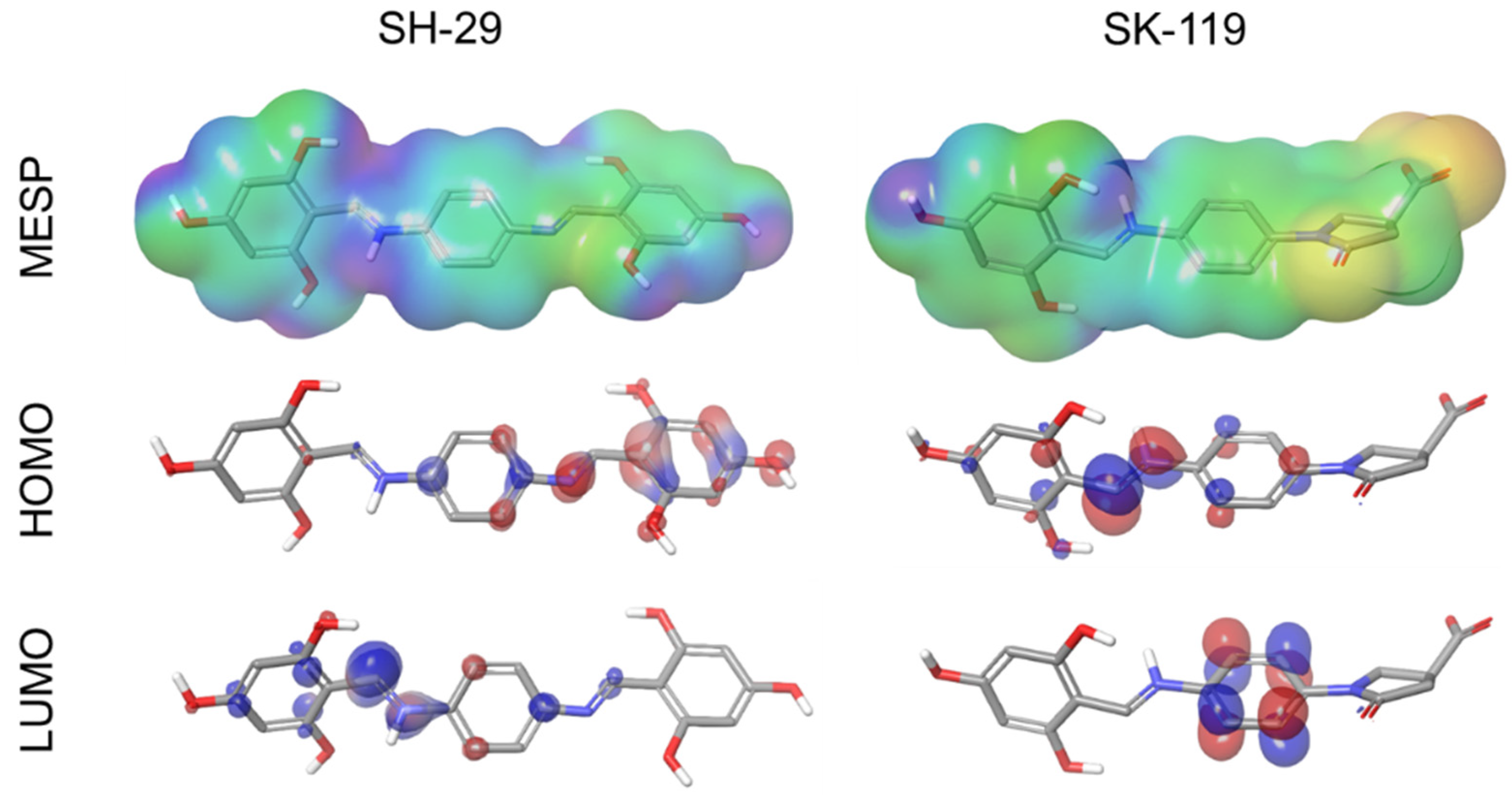
3.3. Molecular Electrostatic Potential (MESP), and HOMO LUMO Stability Analysis of SK-119 and SH-29
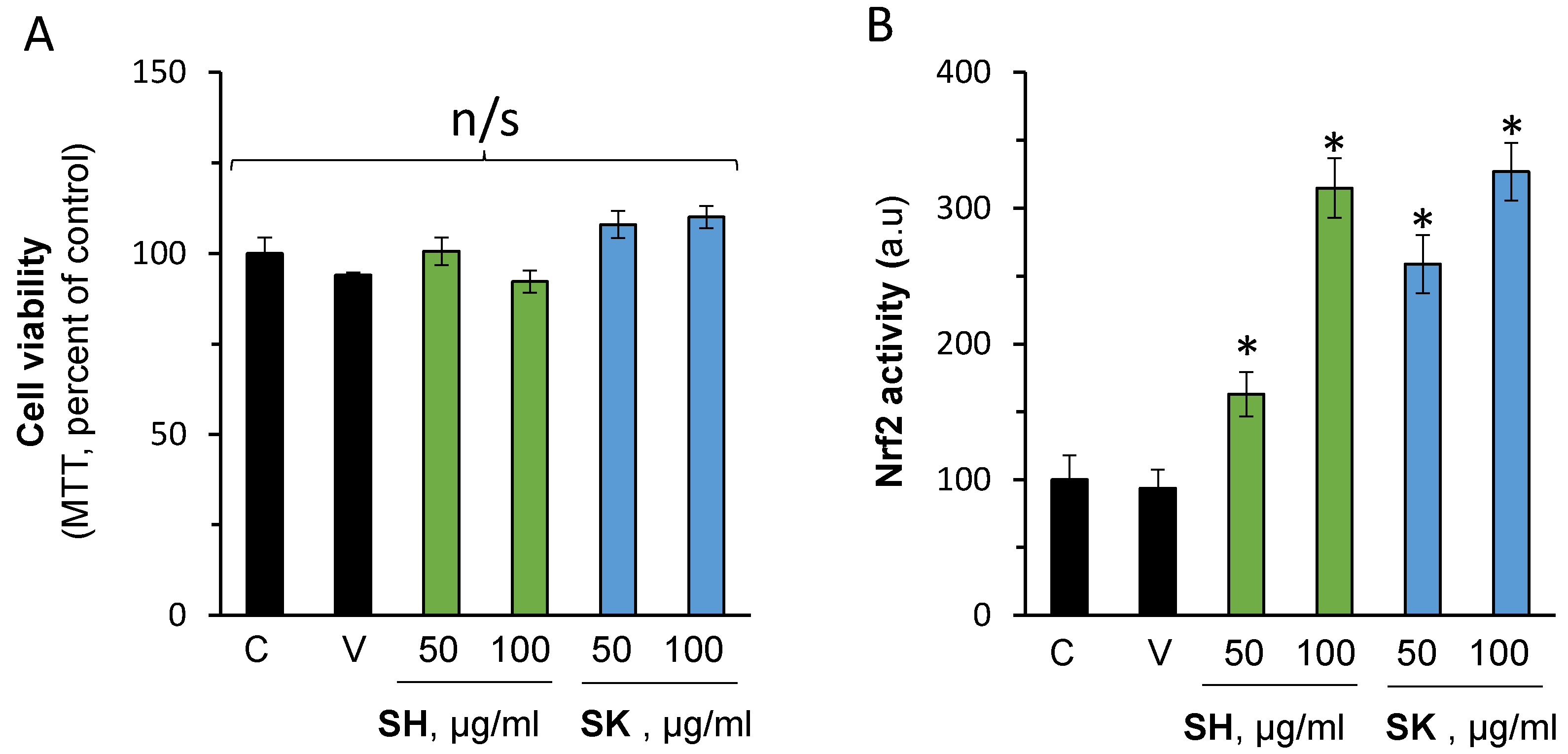
3.4. In Vitro Nrf2 Activation Validation
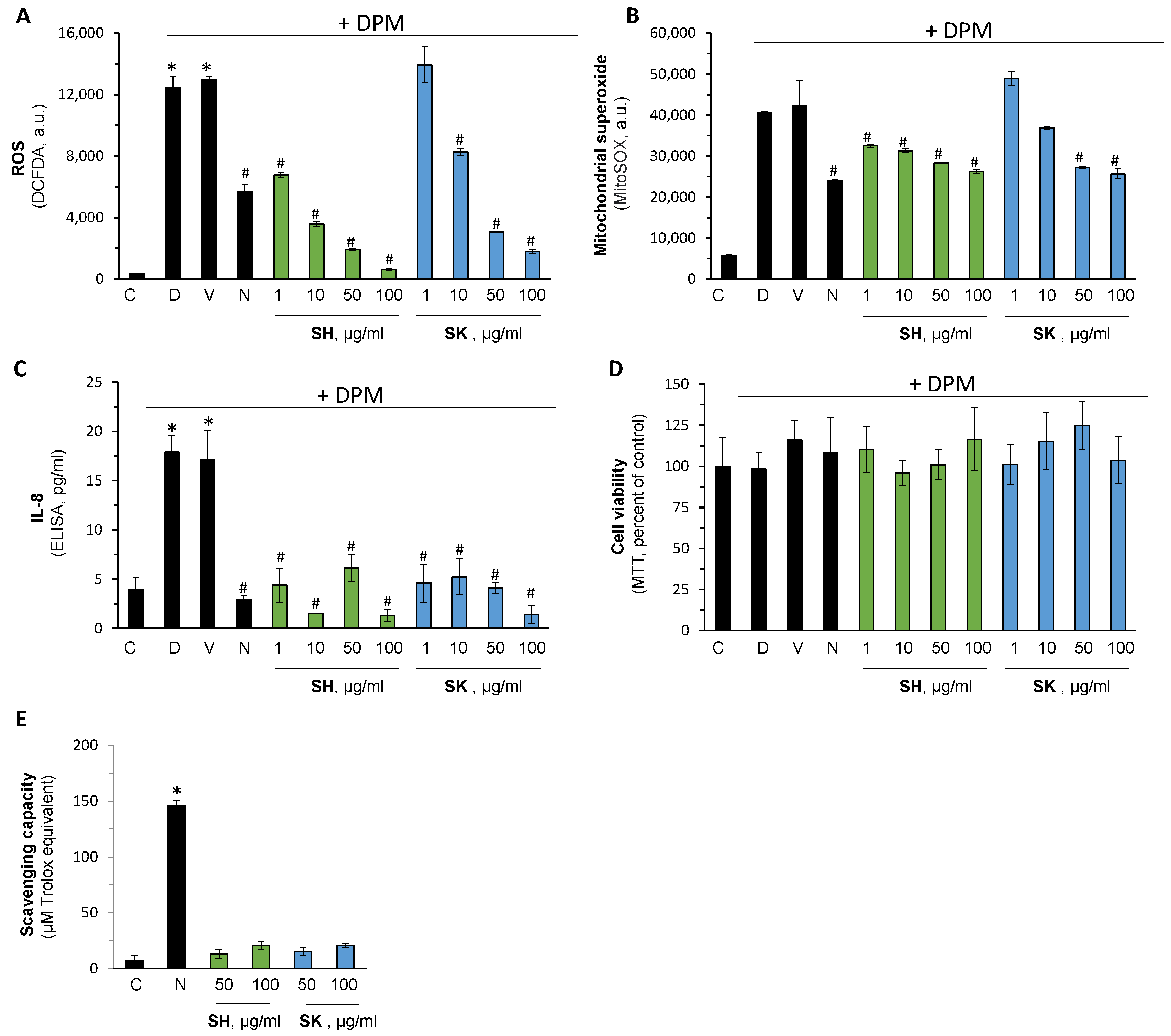
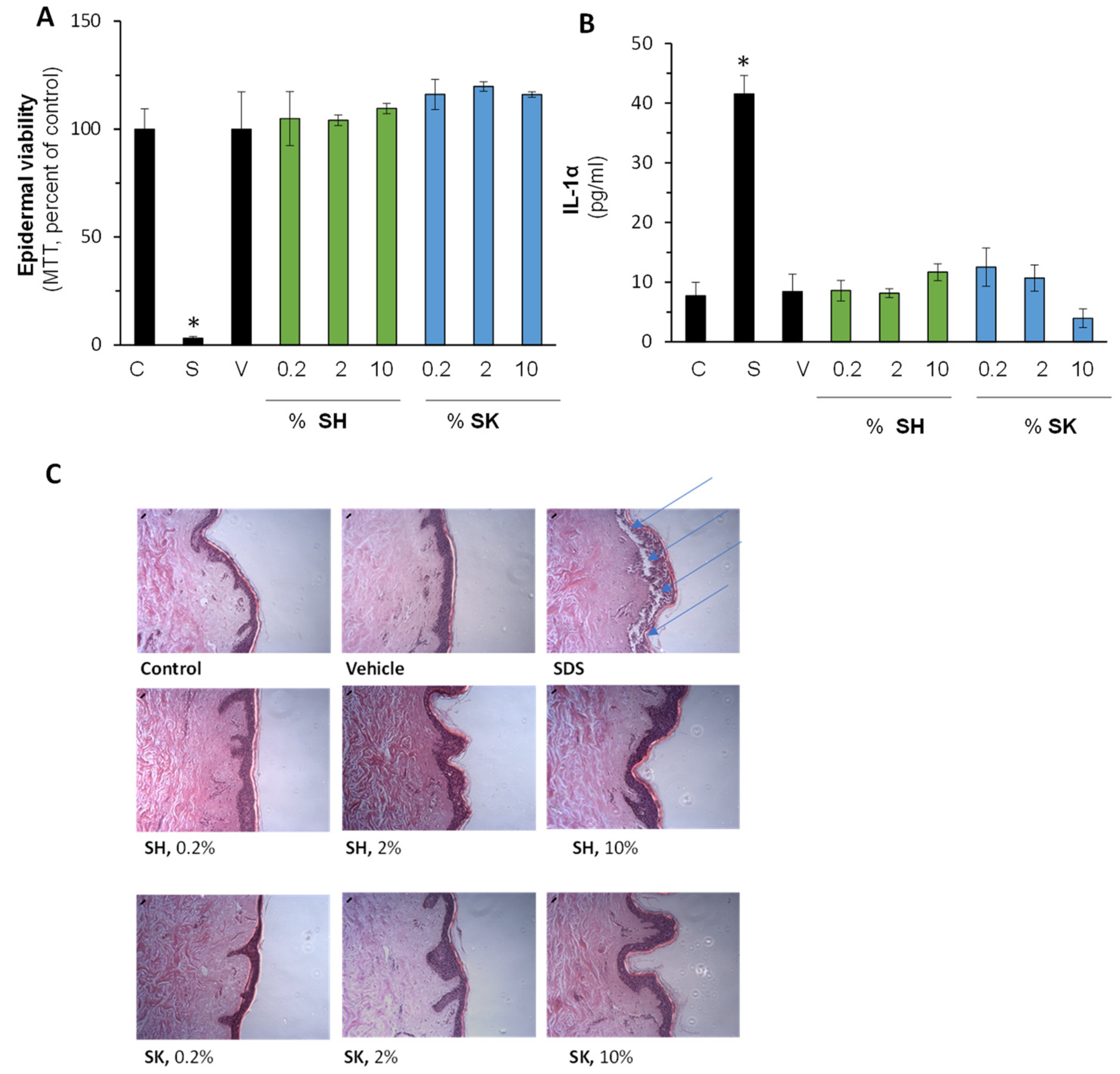
3.5. Pharmacological Activation of Nrf2 Reduce Air Pollution-Induced Damage
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Negev, M. Air Pollution Policy in Israel. Atmosphere 2020, 11, 1065. [Google Scholar] [CrossRef]
- Air Quality Trends Show Clean Air Progress. Available online: https://gispub.epa.gov/air/trendsreport/2021/#air_trends (accessed on 13 November 2021).
- Agency, E.E. Europe’s Air Quality Status 2021. Available online: https://www.eea.europa.eu/ds_resolveuid/XMEN5DFSU8 (accessed on 13 November 2021).
- Chen, H.; Goldberg, M.S. The effects of outdoor air pollution on chronic illnesses. McGill J. Med. 2009, 12, 58–64. [Google Scholar] [CrossRef]
- Drakaki, E.; Dessinioti, C.; Antoniou, C.V. Air pollution and the skin. Front. Environ. Sci. 2014, 2, 11. [Google Scholar] [CrossRef] [Green Version]
- Kahremany, S.; Hofmann, L.; Harari, M.; Gruzman, A.; Cohen, G. Pruritus in psoriasis and atopic dermatitis: Current treatments and new perspectives. Pharmacol. Rep. PR 2021, 73, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Kahremany, S.; Hofmann, L.; Gruzman, A.; Cohen, G. Advances in Understanding the Initial Steps of Pruritoceptive Itch: How the Itch Hits the Switch. Int. J. Mol. Sci. 2020, 21, 4883. [Google Scholar] [CrossRef]
- Kim, K.E.; Cho, D.; Park, H.J. Air pollution and skin diseases: Adverse effects of airborne particulate matter on various skin diseases. Life Sci. 2016, 152, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Hashimoto-Hachiya, A.; Tsuji, G. Aryl Hydrocarbon Receptor in Atopic Dermatitis and Psoriasis. Int. J. Mol. Sci. 2019, 20, 5424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouadi, P.W.; Idriss, S.A.; Naclerio, R.M.; Peden, D.B.; Ansotegui, I.J.; Canonica, G.W.; Gonzalez-Diaz, S.N.; Rosario Filho, N.A.; Ivancevich, J.C.; Hellings, P.W.; et al. Immunopathological features of air pollution and its impact on inflammatory airway diseases (IAD). World Allergy Organ. J. 2020, 13, 100467. [Google Scholar] [CrossRef]
- Zeka, A.; Sullivan, J.R.; Vokonas, P.S.; Sparrow, D.; Schwartz, J. Inflammatory markers and particulate air pollution: Characterizing the pathway to disease. Int. J. Epidemiol. 2006, 35, 1347–1354. [Google Scholar] [CrossRef] [Green Version]
- Puri, P.; Nandar, S.K.; Kathuria, S.; Ramesh, V. Effects of air pollution on the skin: A review. Indian J. Dermatol. Venereol. Leprol. 2017, 83, 415–423. [Google Scholar] [CrossRef]
- Araviiskaia, E.; Berardesca, E.; Bieber, T.; Gontijo, G.; Sanchez Viera, M.; Marrot, L.; Chuberre, B.; Dreno, B. The impact of airborne pollution on skin. J. Eur. Acad. Dermatol. Venereol. JEADV 2019, 33, 1496–1505. [Google Scholar] [CrossRef] [PubMed]
- Silva-Islas, C.A.; Maldonado, P.D. Canonical and non-canonical mechanisms of Nrf2 activation. Pharmacol. Res. 2018, 134, 92–99. [Google Scholar] [CrossRef]
- Rubio, V.; Valverde, M.; Rojas, E. Effects of atmospheric pollutants on the Nrf2 survival pathway. Environ. Sci. Pollut. Res. 2010, 17, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Yamamoto, M. The Molecular Mechanisms Regulating the KEAP1-NRF2 Pathway. Mol. Cell. Biol. 2020, 40, e00099-20. [Google Scholar] [CrossRef] [PubMed]
- Canning, P.; Sorrell, F.J.; Bullock, A.N. Structural basis of Keap1 interactions with Nrf2. Free. Radic. Biol. Med. 2015, 88, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Lo, S.-C.; Li, X.; Henzl, M.T.; Beamer, L.J.; Hannink, M. Structure of the Keap1:Nrf2 interface provides mechanistic insight into Nrf2 signaling. EMBO J. 2006, 25, 3605–3617. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.C.; Ji, J.A.; Jiang, Z.Y.; You, Q.D. The Keap1–Nrf2–ARE pathway as a potential preventive and therapeutic target: An update. Med. Res. Rev. 2016, 36, 924–963. [Google Scholar] [CrossRef]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]
- Shivakumar, D.; Williams, J.; Wu, Y.; Damm, W.; Shelley, J.; Sherman, W. Prediction of Absolute Solvation Free Energies using Molecular Dynamics Free Energy Perturbation and the OPLS Force Field. J. Chem. Theory Comput. 2010, 6, 1509–1519. [Google Scholar] [CrossRef]
- Sirin, S.; Kumar, R.; Martinez, C.; Karmilowicz, M.J.; Ghosh, P.; Abramov, Y.A.; Martin, V.; Sherman, W. A computational approach to enzyme design: Predicting ω-aminotransferase catalytic activity using docking and MM-GBSA scoring. J. Chem. Inf. Modeling 2014, 54, 2334–2346. [Google Scholar] [CrossRef]
- Banavath, H.N.; Sharma, O.P.; Kumar, M.S.; Baskaran, R. Identification of novel tyrosine kinase inhibitors for drug resistant T315I mutant BCR-ABL: A virtual screening and molecular dynamics simulations study. Sci. Rep. 2014, 4, 6948. [Google Scholar] [CrossRef] [Green Version]
- Lyne, P.D.; Lamb, M.L.; Saeh, J.C. Accurate prediction of the relative potencies of members of a series of kinase inhibitors using molecular docking and MM-GBSA scoring. J. Med. Chem. 2006, 49, 4805–4808. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Sun, H.; Li, Y.; Wang, J.; Hou, T. Assessing the performance of MM/PBSA and MM/GBSA methods. 3. The impact of force fields and ligand charge models. J. Phys. Chem. B 2013, 117, 8408–8421. [Google Scholar] [CrossRef]
- Bhattacharjee, A.K.; Gordon, J.A.; Marek, E.; Campbell, A.; Gordon, R.K. 3D-QSAR studies of 2,2-diphenylpropionates to aid discovery of novel potent muscarinic antagonists. Bioorganic Med. Chem. 2009, 17, 3999–4012. [Google Scholar] [CrossRef]
- Kahremany, S.; Babaev, I.; Gvirtz, R.; Ogen-Stern, N.; Azoulay-Ginsburg, S.; Senderowitz, H.; Cohen, G.; Gruzman, A. Nrf2 Activation by SK-119 Attenuates Oxidative Stress, UVB, and LPS-Induced Damage. Ski. Pharmacol. Physiol. 2019, 32, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Gvirtz, R.; Ogen-Shtern, N.; Cohen, G. Kinetic Cytokine Secretion Profile of LPS-Induced Inflammation in the Human Skin Organ Culture. Pharmaceutics 2020, 12, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogen-Shtern, N.; Chumin, K.; Cohen, G.; Borkow, G. Increased pro-collagen 1, elastin, and TGF-β1 expression by copper ions in an ex-vivo human skin model. J. Cosmet. Dermatol. 2020, 19, 1522–1527. [Google Scholar] [CrossRef]
- Roberts, W. Air pollution and skin disorders. Int. J. Womens Dermatol. 2021, 7, 91–97. [Google Scholar] [CrossRef]
- Shaddick, G.; Thomas, M.L.; Mudu, P.; Ruggeri, G.; Gumy, S. Half the world’s population are exposed to increasing air pollution. NPJ Clim. Atmos. Sci. 2020, 3, 23. [Google Scholar] [CrossRef]
- Manisalidis, I.; Stavropoulou, E.; Stavropoulos, A.; Bezirtzoglou, E. Environmental and Health Impacts of Air Pollution: A Review. Front. Public Health 2020, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Owusu, P.A.; Sarkodie, S.A. Global estimation of mortality, disability-adjusted life years and welfare cost from exposure to ambient air pollution. Sci. Total Environ. 2020, 742, 140636. [Google Scholar] [CrossRef] [PubMed]
- Adamkiewicz, Ł.; Badyda, A.J.; Gayer, A.; Mucha, D. Disability-adjusted life years in the assessment of health effects of traffic-related air pollution. Adv. Exp. Med. Biol. 2015, 834, 15–20. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. Summary of the Clean Air Act; U.S. Environmental Protection Agency: Washington, DC, USA, 1970. [Google Scholar]
- Huang, J.; Pan, X.; Guo, X.; Li, G. Health impact of China’s Air Pollution Prevention and Control Action Plan: An analysis of national air quality monitoring and mortality data. Lancet Planet. Health 2018, 2, e313–e323. [Google Scholar] [CrossRef] [Green Version]
- Tran, V.V.; Park, D.; Lee, Y.-C. Indoor Air Pollution, Related Human Diseases, and Recent Trends in the Control and Improvement of Indoor Air Quality. Int. J. Environ. Res. Public Health 2020, 17, 2927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laumbach, R.; Meng, Q.; Kipen, H. What can individuals do to reduce personal health risks from air pollution? J. Thorac. Dis. 2015, 7, 96–107. [Google Scholar] [CrossRef]
- Ou, Y.; West, J.J.; Smith, S.J.; Nolte, C.G.; Loughlin, D.H. Air pollution control strategies directly limiting national health damages in the US. Nat. Commun. 2020, 11, 957. [Google Scholar] [CrossRef] [Green Version]
- Emadi, A. Transportation 2.0. IEEE Power Energy Mag. 2011, 9, 18–29. [Google Scholar] [CrossRef]
- Hohmeyer, O.H.; Bohm, S. Trends toward 100% renewable electricity supply in Germany and Europe: A paradigm shift in energy policies. WIREs Energy Environ. 2015, 4, 74–97. [Google Scholar] [CrossRef]
- Manfren, M.; Caputo, P.; Costa, G. Paradigm shift in urban energy systems through distributed generation: Methods and models. Appl. Energy 2011, 88, 1032–1048. [Google Scholar] [CrossRef]
- Gugliandolo, A.; Bramanti, P.; Mazzon, E. Activation of Nrf2 by Natural Bioactive Compounds: A Promising Approach for Stroke? Int. J. Mol. Sci. 2020, 21, 4875. [Google Scholar] [CrossRef]
- Kahremany, S.; Babaev, I.; Hasin, P.; Tamir, T.Y.; Ben-Zur, T.; Cohen, G.; Jiang, Z.; Weintraub, S.; Offen, D.; Rahimipour, S.; et al. Computer-Aided Design and Synthesis of 1-{4-[(3,4-Dihydroxybenzylidene)amino]phenyl}-5-oxopyrrolidine-3-carboxylic Acid as an Nrf2 Enhancer. ChemPlusChem 2018, 83, 318. [Google Scholar] [CrossRef]
- Xian, D.; Xiong, X.; Xu, J.; Xian, L.; Lei, Q.; Song, J.; Zhong, J. Nrf2 Overexpression for the Protective Effect of Skin-Derived Precursors against UV-Induced Damage: Evidence from a Three-Dimensional Skin Model. Oxidative Med. Cell. Longev. 2019, 2019, 7021428. [Google Scholar] [CrossRef] [Green Version]
- Saw, C.L.L.; Yang, A.Y.; Huang, M.-T.; Liu, Y.; Lee, J.H.; Khor, T.O.; Su, Z.-Y.; Shu, L.; Lu, Y.; Conney, A.H.; et al. Nrf2 null enhances UVB-induced skin inflammation and extracellular matrix damages. Cell Biosci. 2014, 4, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helou, D.G.; Martin, S.F.; Pallardy, M.; Chollet-Martin, S.; Kerdine-Römer, S. Nrf2 Involvement in Chemical-Induced Skin Innate Immunity. Front. Immunol. 2019, 10, 1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saw, C.L.; Huang, M.T.; Liu, Y.; Khor, T.O.; Conney, A.H.; Kong, A.N. Impact of Nrf2 on UVB-induced skin inflammation/photoprotection and photoprotective effect of sulforaphane. Mol. Carcinog. 2011, 50, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Dijkhoff, I.M.; Drasler, B.; Karakocak, B.B.; Petri-Fink, A.; Valacchi, G.; Eeman, M.; Rothen-Rutishauser, B. Impact of airborne particulate matter on skin: A systematic review from epidemiology to in vitro studies. Part. Fibre Toxicol. 2020, 17, 35. [Google Scholar] [CrossRef]
- Rajagopalan, P.; Jain, A.P.; Nanjappa, V.; Patel, K.; Mangalaparthi, K.K.; Babu, N.; Cavusoglu, N.; Roy, N.; Soeur, J.; Breton, L.; et al. Proteome-wide changes in primary skin keratinocytes exposed to diesel particulate extract—A role for antioxidants in skin health. J. Dermatol. Sci. 2018, 91, 239–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, K.; Park, S.-H.; Han, B.S.; Oh, S.W.; Lee, S.E.; Yoo, J.A.; Park, S.J.; Kim, J.; Kim, J.W.; Cho, J.Y.; et al. Negative Cellular Effects of Urban Particulate Matter on Human Keratinocytes Are Mediated by P38 MAPK and NF-κB-dependent Expression of TRPV 1. Int. J. Mol. Sci. 2018, 19, 2660. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, W.J.; Hanson, P.S.; Critchley, A.; Griffiths, B.; Chavan, B.; Birch-Machin, M.A. Exposing human primary dermal fibroblasts to particulate matter induces changes associated with skin aging. FASEB J. 2020, 34, 14725–14735. [Google Scholar] [CrossRef]
- Piao, M.J.; Ahn, M.J.; Kang, K.A.; Ryu, Y.S.; Hyun, Y.J.; Shilnikova, K.; Zhen, A.X.; Jeong, J.W.; Choi, Y.H.; Kang, H.K.; et al. Particulate matter 2.5 damages skin cells by inducing oxidative stress, subcellular organelle dysfunction, and apoptosis. Arch. Toxicol. 2018, 92, 2077–2091. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.C.; Lee, C.W.; Tsai, M.H.; Ko, H.H.; Fang, J.Y.; Chiang, Y.C.; Liang, C.J.; Hsu, L.F.; Hu, S.C.; Yen, F.L. Eupafolin nanoparticles protect HaCaT keratinocytes from particulate matter-induced inflammation and oxidative stress. Int. J. Nanomed. 2016, 11, 3907–3926. [Google Scholar] [CrossRef] [Green Version]
- Kovac, S.; Angelova, P.R.; Holmström, K.M.; Zhang, Y.; Dinkova-Kostova, A.T.; Abramov, A.Y. Nrf2 regulates ROS production by mitochondria and NADPH oxidase. Biochim. Et Biophys. Acta (BBA)—Gen. Subj. 2015, 1850, 794–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardo, M.; Qiu, X.; Zimmermann, R.; Rudich, Y. Particulate Matter Toxicity Is Nrf2 and Mitochondria Dependent: The Roles of Metals and Polycyclic Aromatic Hydrocarbons. Chem. Res. Toxicol. 2020, 33, 1110–1120. [Google Scholar] [CrossRef]
- Pardo, M.; Xu, F.; Shemesh, M.; Qiu, X.; Barak, Y.; Zhu, T.; Rudich, Y. Nrf2 protects against diverse PM(2.5) components-induced mitochondrial oxidative damage in lung cells. Sci. Total Environ. 2019, 669, 303–313. [Google Scholar] [CrossRef]
- Li, Q.; Kang, Z.; Jiang, S.; Zhao, J.; Yan, S.; Xu, F.; Xu, J. Effects of Ambient Fine Particles PM(2.5) on Human HaCaT Cells. Int. J. Environ. Res. Public Health 2017, 14, 72. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zheng, L.; Tuo, J.; Liu, Q.; Zhang, X.; Xu, Z.; Liu, S.; Sui, G. Analysis of PM2.5-induced cytotoxicity in human HaCaT cells based on a microfluidic system. Toxicol. In Vitro 2017, 43, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Andrade, T.A.; Aguiar, A.F.; Guedes, F.A.; Leite, M.N.; Caetano, G.F.; Coelho, E.B.; Das, P.K.; Frade, M.A. Ex vivo Model of Human Skin (hOSEC) as Alternative to Animal use for Cosmetic Tests. Procedia Eng. 2015, 110, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Corzo-León, D.E.; Munro, C.A.; MacCallum, D.M. An ex vivo Human Skin Model to Study Superficial Fungal Infections. Front. Microbiol. 2019, 10, 1172. [Google Scholar] [CrossRef] [Green Version]
- Ogen-Shtern, N.; Chumin, K.; Silberstein, E.; Borkow, G. Copper Ions Ameliorated Thermal Burn-Induced Damage in ex vivo Human Skin Organ Culture. Ski. Pharmacol. Physiol. 2021, 34, 317–327. [Google Scholar] [CrossRef]
- Hofmann, E.; Fink, J.; Eberl, A.; Prugger, E.-M.; Kolb, D.; Luze, H.; Schwingenschuh, S.; Birngruber, T.; Magnes, C.; Mautner, S.I.; et al. A novel human ex vivo skin model to study early local responses to burn injuries. Sci. Rep. 2021, 11, 364. [Google Scholar] [CrossRef]
- Reus, A.A.; Usta, M.; Krul, C.A.M. The use of ex vivo human skin tissue for genotoxicity testing. Toxicol. Appl. Pharmacol. 2012, 261, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Eberlin, S.; Silva, M.S.D.; Facchini, G.; Silva, G.H.D.; Pinheiro, A.; Eberlin, S.; Pinheiro, A.D.S. The Ex Vivo Skin Model as an Alternative Tool for the Efficacy and Safety Evaluation of Topical Products. Altern. Lab. Anim. ATLA 2020, 48, 10–22. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | ∆Gbind a | ∆Gcoulomb b | ∆Gcovalent c | ∆GH-bond d | ∆Gsol lipo e | ∆Gsol GB f | ∆Gvdw g |
|---|---|---|---|---|---|---|---|
| SH-29 | −54.58 | 6.62 | 2.17 | −1.27 | −16.53 | −22.94 | −22.01 |
| SK-119 | −49.48 | 1.09 | 5.72 | −2.86 | −20.97 | −3.49 | −28.9 |
| Compound | HOMO (ev) | LUMO (ev) | HOMO-LUMO GAP (ev) | MESP (kcal/mol) |
|---|---|---|---|---|
| SH-29 | −0.222 | −0.113 | 0.109 | −6.94 to 150.47 |
| SK-119 | −0.127 | −0.014 | 0.114 | −167.78 to 39.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kahremany, S.; Hofmann, L.; Eretz-Kdosha, N.; Silberstein, E.; Gruzman, A.; Cohen, G. SH-29 and SK-119 Attenuates Air-Pollution Induced Damage by Activating Nrf2 in HaCaT Cells. Int. J. Environ. Res. Public Health 2021, 18, 12371. https://doi.org/10.3390/ijerph182312371
Kahremany S, Hofmann L, Eretz-Kdosha N, Silberstein E, Gruzman A, Cohen G. SH-29 and SK-119 Attenuates Air-Pollution Induced Damage by Activating Nrf2 in HaCaT Cells. International Journal of Environmental Research and Public Health. 2021; 18(23):12371. https://doi.org/10.3390/ijerph182312371
Chicago/Turabian StyleKahremany, Shirin, Lukas Hofmann, Noy Eretz-Kdosha, Eldad Silberstein, Arie Gruzman, and Guy Cohen. 2021. "SH-29 and SK-119 Attenuates Air-Pollution Induced Damage by Activating Nrf2 in HaCaT Cells" International Journal of Environmental Research and Public Health 18, no. 23: 12371. https://doi.org/10.3390/ijerph182312371
APA StyleKahremany, S., Hofmann, L., Eretz-Kdosha, N., Silberstein, E., Gruzman, A., & Cohen, G. (2021). SH-29 and SK-119 Attenuates Air-Pollution Induced Damage by Activating Nrf2 in HaCaT Cells. International Journal of Environmental Research and Public Health, 18(23), 12371. https://doi.org/10.3390/ijerph182312371