
Learning Monologues at Bedtime Improves Sleep Quality in Actors and Non-Actors

 ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Learning Task
2.3. Procedure
2.4. Sleep Recordings and Sleep Measures
- Sleep continuity: Total frequency of awakenings per hour of AST; frequency of brief (<4 epochs) and long (≥4 epochs) awakenings per hour of AST
- Sleep stability: Frequency of arousals per hour of AST (here arousals are defined as all transitions to shallower NREM sleep stages and from REM sleep to N1); frequency of state transitions (defined as all transitions from one state to another) per hour of TST; frequency of “functional uncertainty periods” (FU periods; defined as periods in which a minimum of 3 state transitions follow one another with no longer than 1.5 min intervals) per hour of TST; percentage of total time spent in FU (TFU) over TST;
- Sleep organization: Number of complete sleep cycles, defined as sequences of NREM and REM sleep (each lasting at least 10 min) not interrupted by periods of wake longer than 2 min (as in [9]); percentage of total time spent in cycles (TCT) over TST. A necessary methodological remark concerns our choice of an extremely conservative definition of the sleep cycle, based on the assumption that a non-marginal amount of each sleep state is required for the NREM-REM cycle to exert its role in sleep-dependent memory processes [36]. With such a definition, the duration of a sleep cycle is remarkably shorter, on average, than that obtained with the more common, less conservative, definitions.
2.5. Performance Measures
2.6. Data Analysis
3. Results
3.1. Sleepiness and Fatigue
3.2. Classical Sleep Variables
3.3. Sleep Continuity
3.4. Sleep Stability
3.5. Sleep Cyclic Organization
3.6. Memory Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borbély, A.A. A two process model of sleep regulation. Hum. Neurobiol. 1982, 1, 195–204. [Google Scholar]
- Feinberg, I. Changes in sleep cycle patterns with age. J. Psychiatr. Res. 1974, 10, 283–306. [Google Scholar] [CrossRef]
- Feinberg, I.; Campbell, I. Ketamine Administration during Waking Increases Delta EEG Intensity in Rat Sleep. Neuropsychopharmacology 1993, 9, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Campbell, I.G.; Feinberg, I. Noncompetitive NMDA channel blockade during waking intensely stimulates NREM delta. J. Pharmacol. Exp. Ther. 1996, 276, 737–742. [Google Scholar]
- Campbell, I.G.; Feinberg, I. NREM delta stimulation following MK-801 is a response of sleep systems. J. Neurophysiol. 1996, 76, 3714–3720. [Google Scholar] [CrossRef] [PubMed]
- Meerlo, P.; Pragt, B.J.; Daan, S. Social stress induces high intensity sleep in rats. Neurosci. Lett. 1997, 225, 41–44. [Google Scholar] [CrossRef]
- Franken, P. The quality of waking and process S. Sleep 2007, 30, 126–127. [Google Scholar] [CrossRef] [Green Version]
- Huber, R.; Tononi, G.; Cirelli, C. Exploratory behavior, cortical BDNF expression, and sleep homeostasis. Sleep 2007, 30, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Conte, F.; Carobbi, G.; Errico, B.M.; Ficca, G. The effects of pre-sleep learning on sleep continuity, stability, and organization in elderly individuals. Front. Neurol. 2012, 3, 109. [Google Scholar] [CrossRef] [Green Version]
- Haimov, I.; Shatil, E. Cognitive training improves sleep quality and cognitive function among older adults with insomnia. PLoS ONE 2013, 8, e61390. [Google Scholar] [CrossRef] [Green Version]
- Arzilli, C.; Cerasuolo, M.; Conte, F.; Bittoni, V.; Gatteschi, C.; Albinni, B.; Giganti, F.; Ficca, G. The effect of cognitive activity on sleep maintenance in a subsequent daytime nap. Behav. Sleep Med. 2019, 17, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Cerasuolo, M.; Conte, F.; Cellini, N.; Fusco, G.; Giganti, F.; Malloggi, S.; Ficca, G. The effect of complex cognitive training on subsequent night sleep. J. Sleep Res. 2020, 29, e12929. [Google Scholar] [CrossRef] [PubMed]
- Conte, F.; Ficca, G. Caveats on psychological models of sleep and memory: A compass in an overgrown scenario. Sleep Med. Rev. 2013, 17, 105–121. [Google Scholar] [CrossRef]
- Cerasuolo, M.; Conte, F.; Giganti, F.; Ficca, G. Sleep changes following intensive cognitive activity. Sleep Med. 2020, 66, 148–158. [Google Scholar] [CrossRef]
- Fogel, S.M.; Smith, C.T. Learning-dependent changes in sleep spindles and Stage 2 sleep. J. Sleep Res. 2006, 15, 250–255. [Google Scholar] [CrossRef]
- Schmidt, C.; Peigneux, P.; Muto, V.; Schenkel, M.; Knoblauch, V.; Münch, M.; de Quervain, D.J.-F.; Wirz-Justice, A.; Cajochen, C. Encoding difficulty promotes postlearning changes in sleep spindle activity during napping. J. Neurosci. 2006, 26, 8976–8982. [Google Scholar] [CrossRef]
- Johnson, L.A.; Blakely, T.; Hermes, D.; Hakimian, S.; Ramsey, N.F.; Ojemann, J.G. Sleep spindles are locally modulated by training on a brain–computer interface. Proc. Natl Acad. Sci. USA 2012, 109, 18583–18588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirov, R.; Kolev, V.; Yordanova, J. Labile sleep promotes awareness of abstract knowledge in a serial reaction time task. Front. Psychol. 2015, 6, 1354. [Google Scholar] [CrossRef] [Green Version]
- Sergeeva, V.; Viczko, J.; Ray, L.B.; Owen, A.M.; Fogel, S.M. Sleep-dependent motor sequence memory consolidation in individuals with periodic limb movements. Sleep Med. 2017, 40, 23–32. [Google Scholar] [CrossRef]
- Horne, J.A.; Minard, A. Sleep and sleepiness following a behaviourally ‘active’ day. Ergonomics 1985, 28, 567–575. [Google Scholar] [CrossRef]
- Abou-Ismail, U.A.; Burman, O.H.; Nicol, C.J.; Mendl, M. The effects of enhancing cage complexity on the behaviour and welfare of laboratory rats. Behav. Process. 2010, 85, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Hauptmann, B.; Reinhart, E.; Brandt, S.A.; Karni, A. The predictive value of the leveling off of within-session performance for procedural memory consolidation. Brain Res. Cogn. Brain Res. 2005, 24, 181–189. [Google Scholar] [CrossRef]
- Tucker, M.A.; Fishbein, W. Enhancement of declarative memory performance following a daytime nap is contingent on strength of initial task acquisition. Sleep 2008, 31, 197–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, R.; Ghilardi, M.F.; Massimini, M.; Tononi, G. Local sleep and learning. Nature 2004, 430, 78–81. [Google Scholar] [CrossRef]
- Bergmann, T.O.; Mölle, M.; Diedrichs, J.; Born, J.; Siebner, H.R. Sleep spindle-related reactivation of category-specific cortical regions after learning face-scene associations. Neuroimage 2012, 59, 2733–2742. [Google Scholar] [CrossRef]
- Curcio, G.; Tempesta, D.; Scarlata, S.; Marzano, C.; Moroni, F.; Rossini, P.M.; Ferrara, M.; De Gennaro, L. Validity of the Italian Version of the Pittsburgh Sleep Quality Index (PSQI). Neurol. Sci. 2013, 34, 511–519. [Google Scholar] [CrossRef]
- Sica, C.; Ghisi, M. The Italian versions of the Beck Anxiety Inventory and the Beck Depression Inventory-II: Psychometric properties and discriminant power. In Leading-Edge Psychological Tests and Testing Research; Lange, M.A., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2007; pp. 27–50. [Google Scholar]
- Buysse, D.J.; Reynolds, C.F.; Monk, T.H.; Berman, S.R.; Kupfer, D.J. The Pittsburgh Sleep Quality Index: A new instrument for psychiatric practice and research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef]
- Beck, A.T.; Epstein, N.; Brown, G.; Steer, R.A. An inventory for measuring clinical anxiety: Psychometric properties. J. Consult. Clin. Psychol. 1988, 56, 893–897. [Google Scholar] [CrossRef] [PubMed]
- Beck, A.T.; Steer, R.A.; Brown, G.K. Manual for the Beck Depression Inventory-II; Psychological Corporation: San Antonio, TX, USA, 1996. [Google Scholar]
- Cousse, R. Stratégie pour Deux Jambons: Roman Chochon; Flammarion: Paris, France, 1992. [Google Scholar]
- Spinnler, H.; Tognoni, G. Italian Group on the Neuropsychological Study of Ageing: Italian standardization and classification of neuropsychological tests. Ital. J. Neurol. Sci. 1987, 6 (Suppl. S8), 1–120. [Google Scholar]
- Åkerstedt, T.; Gillberg, M. Subjective and objective sleepiness in the active individual. Int. J. Neurosci. 1990, 52, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Hewlett, S.; Dures, E.; Almeida, C. Measures of fatigue: Bristol Rheumatoid Arthritis Fatigue Multi-Dimensional Questionnaire (BRAF MDQ), Bristol Rheumatoid Arthritis Fatigue Numerical Rating Scales (BRAF NRS) for Severity, Effect, and Coping, Chalder Fatigue Questionnaire (CFQ), Checklist Individual Strength (CIS20R and CIS8R), Fatigue Severity Scale (FSS), Functional Assessment Chronic Illness Therapy (Fatigue) (FACIT-F), Multi-Dimensional Assessment of Fatigue (MAF), Multi-Dimensional Fatigue Inventory (MFI), Pediatric Quality Of Life (PedsQL) Multi-Dimensional Fatigue Scale, Profile of Fatigue (ProF), Short Form 36 Vitality Subscale (SF-36 VT), and Visual Analog Scales (VAS). Arthritis Care Res. 2011, 63 (Suppl. S11), S263–S286. [Google Scholar] [CrossRef]
- Iber, C.; Ancoli-Israel, S.; Chesson, A.L.; Quan, S.F. The new sleep scoring manual—The evidence behind the rules. J. Clin. Sleep Med. 2007, 3, 107. [Google Scholar] [CrossRef] [Green Version]
- Ficca, G.; Salzarulo, P. What in sleep is for memory. Sleep Med. 2004, 5, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Jamovi, Version 1.6; Computer Software; The Jamovi Project: Sydney, Australia, 2021. Available online: https://www.jamovi.org (accessed on 12 May 2021).
- Buchegger, J.; Meier-Koll, A. Motor learning and ultradian sleep cycle: An electroencephalographic study of trampoliners. Percept. Mot. Ski. 1988, 67, 635–645. [Google Scholar] [CrossRef]
- Higginson, C.I.; Arnett, P.A.; Voss, W.D. The ecological validity of clinical tests of memory and attention in multiple sclerosis. Arch. Clin. Neuropsychol. 2000, 15, 185–204. [Google Scholar] [CrossRef]
- Swihart, B.J.; Caffo, B.; Bandeen-Roche, K.; Punjabi, N.M. Characterizing sleep structure using the hypnogram. J. Clin. Sleep Med. 2008, 4, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Kishi, A.; Natelson, B.H.; Togo, F.; Struzik, Z.R.; Rapoport, D.M.; Yamamoto, Y. Sleep stage transitions in chronic fatigue syndrome patients with or without fibromyalgia. In Proceedings of the Annual International Conference of the IEEE Engineering in Medicine and Biology, Buenos Aires, Argentina, 31 August–4 September 2010; pp. 5391–5394. [Google Scholar] [CrossRef]
- Conte, F.; Cerasuolo, M.; Fusco, G.; Giganti, F.; Inserra, I.; Malloggi, S.; Di Iorio, I.; Ficca, G. Sleep continuity, stability and organization in good and bad sleepers. J. Health Psychol. 2020, 26, 2131–2142. [Google Scholar] [CrossRef] [PubMed]
- Klerman, E.B.; Davis, J.B.; Duffy, J.F.; Dijk, D.J.; Kronauer, R.E. Older people awaken more frequently but fall back asleep at the same rate as younger people. Sleep 2004, 27, 793–798. [Google Scholar] [CrossRef] [Green Version]
- Klerman, E.B.; Wang, W.; Duffy, J.F.; Dijk, D.J.; Czeisler, C.A.; Kronauer, R.E. Survival analysis indicates that age-related decline in sleep continuity occurs exclusively during NREM sleep. Neurobiol. Aging 2013, 34, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Djonlagic, I.; Saboisky, J.; Carusona, A.; Stickgold, R.; Malhotra, A. Increased sleep fragmentation leads to impaired off-line consolidation of motor memories in humans. PLoS ONE 2012, 7, e34106. [Google Scholar] [CrossRef] [Green Version]
- Stickgold, R.; James, L.; Hobson, J. Visual discrimination learning requires sleep after training. Nat. Neurosci. 2000, 3, 1237–1238. [Google Scholar] [CrossRef]
- Ambrosini, M.V.; Giuditta, A. Learning and sleep: The sequential hypothesis. Sleep Med. Rev. 2001, 5, 477–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gais, S.; Mölle, M.; Helms, K.; Born, J. Learning-dependent increases in sleep spindle density. J. Neurosci. 2002, 22, 6830–6834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogel, S.M.; Ray, L.B.; Binnie, L.; Owen, A.M. How to become an expert: A new perspective on the role of sleep in the mastery of procedural skills. Neurobiol. Learn. Mem. 2015, 125, 236–248. [Google Scholar] [CrossRef]
- Conte, F.; Cerasuolo, M.; Giganti, F.; Ficca, G. Sleep enhances strategic thinking at the expense of basic procedural skills consolidation. J. Sleep Res. 2020, 29, e13034. [Google Scholar] [CrossRef]
- Fischer, S.; Born, J. Anticipated reward enhances offline learning during sleep. J. Exp. Psychol Learn. 2009, 35, 1586. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, I.; Diekelmann, S.; Molzow, I.; Ayoub, A.; Mölle, M.; Born, J. Sleep selectively enhances memory expected to be of future relevance. J. Neurosci. 2011, 31, 1563–1569. [Google Scholar] [CrossRef] [Green Version]
- Landmann, N.; Kuhn, M.; Piosczyk, H.; Feige, B.; Baglioni, C.; Spiegelhalder, K.; Frase, K.; Riemann, D.; Sterr, A.; Nissen, C. The reorganisation of memory during sleep. Sleep Med. Rev. 2014, 18, 531–541. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Descriptives | Statistics | ||||||
|---|---|---|---|---|---|---|---|
| Group | Condition | m ± sd | Effects | F | p | η2 | |
| Sleep Onset Latency (min) | Actors | BL | 11.4 ± 8.05 | Condition | 60.87 | <0.001 | 0.195 |
| TR | 5.7 ± 4.03 | Interaction | 5.08 | 0.036 | 0.016 | ||
| non-Actors | BL | 20.7 ± 11 | Group | 5.08 | 0.036 | 0.147 | |
| TR | 10.3 ± 5.51 | ||||||
| Time in Bed (h) | Actors | BL | 7.55 ± 1.39 | Condition | 0.23 | 0.630 | 0.004 |
| TR | 7.38 ± 0.97 | Interaction | 0.01 | 0.915 | 0.000 | ||
| non-Actors | BL | 7.36 ± 1.27 | Group | 0.17 | 0.682 | 0.005 | |
| TR | 7.25 ± 0.94 | ||||||
| Total Sleep Time (h) | Actors | BL | 7.08 ± 1.43 | Condition | 0.018 | 0.892 | 0.000 |
| TR | 7.03 ± 0.93 | Interaction | 0.085 | 0.774 | 0.001 | ||
| non-Actors | BL | 6.70 ± 1.36 | Group | 0.541 | 0.471 | 0.017 | |
| TR | 6.82 ± 0.82 | ||||||
| Actual Sleep Time (h) | Actors | BL | 6.79 ± 1.47 | Condition | 0.577 | 0.456 | 0.010 |
| TR | 6.89 ± 10 | Interaction | 0.194 | 0.665 | 0.003 | ||
| non-Actors | BL | 6.38 ± 1.54 | Group | 0.527 | 0.527 | 0.013 | |
| TR | 6.75 ± 0.78 | ||||||
| Stage1 (%) | Actors | BL | 22.6 ± 7.61 | Condition | 1.10 | 0.305 | 0.020 |
| TR | 21.1 ± 9.23 | Interaction | 0.319 | 0.579 | 0.006 | ||
| non-Actors | BL | 32.4 ± 14.7 | Group | 4.90 | 0.039 | 0.120 | |
| TR | 27.4 ± 12.1 | ||||||
| Stage2 (%) | Actors | BL | 45 ± 8.43 | Condition | 0.112 | 0.742 | 0.002 |
| TR | 46 ± 6.51 | Interaction | 0.510 | 0.483 | 0.010 | ||
| non-Actors | BL | 42.6 ± 10.1 | Group | 2.14 | 0.159 | 0.056 | |
| TR | 39.9 ± 10.7 | ||||||
| SWS (%) | Actors | BL | 13.6 ± 6.64 | Condition | 0.955 | 0.797 | 0.001 |
| TR | 14.3 ± 3.46 | Interaction | 0.1399 | 0.483 | 0.010 | ||
| non-Actors | BL | 11.2 ± 5.89 | Group | 4.09 | 0.057 | 0.098 | |
| TR | 9.58 ± 5.99 | ||||||
| REM (%) | Actors | BL | 18.2 ± 6.8 | Condition | 1.48 | 0.237 | 0.024 |
| TR | 19 ± 3.94 | Interaction | 0.450 | 0.510 | 0.007 | ||
| non-Actors | BL | 19.4 ± 4.44 | Group | 0.281 | 0.281 | 0.038 | |
| TR | 22.1 ± 6.57 | ||||||
| Wake After Sleep Onset (%) | Actors | BL | 4.2 ± 4.06 | Condition | 9.13 | 0.007 | 0.127 |
| TR | 2.1 ± 3.87 | Interaction | 1.28 | 0.272 | 0.018 | ||
| non-Actors | BL | 5.55 ± 7.06 | Group | 0.004 | 0.948 | 0.000 | |
| TR | 0.95 ± 1.4 | ||||||
| Sleep Efficiency | Actors | BL | 89.8 ± 7.92 | Condition | 8.54 | 0.008 | 0.121 |
| TR | 93.5 ± 6.78 | Interaction | 0.959 | 0.339 | 0.014 | ||
| non-Actors | BL | 85.9 ± 10.8 | Group | 0.534 | 0.473 | 0.015 | |
| TR | 93.4 ± 4.23 | ||||||
| Descriptives | Statistics | ||||||
|---|---|---|---|---|---|---|---|
| Group | Condition | m ± sd | Effects | F | p | η2 | |
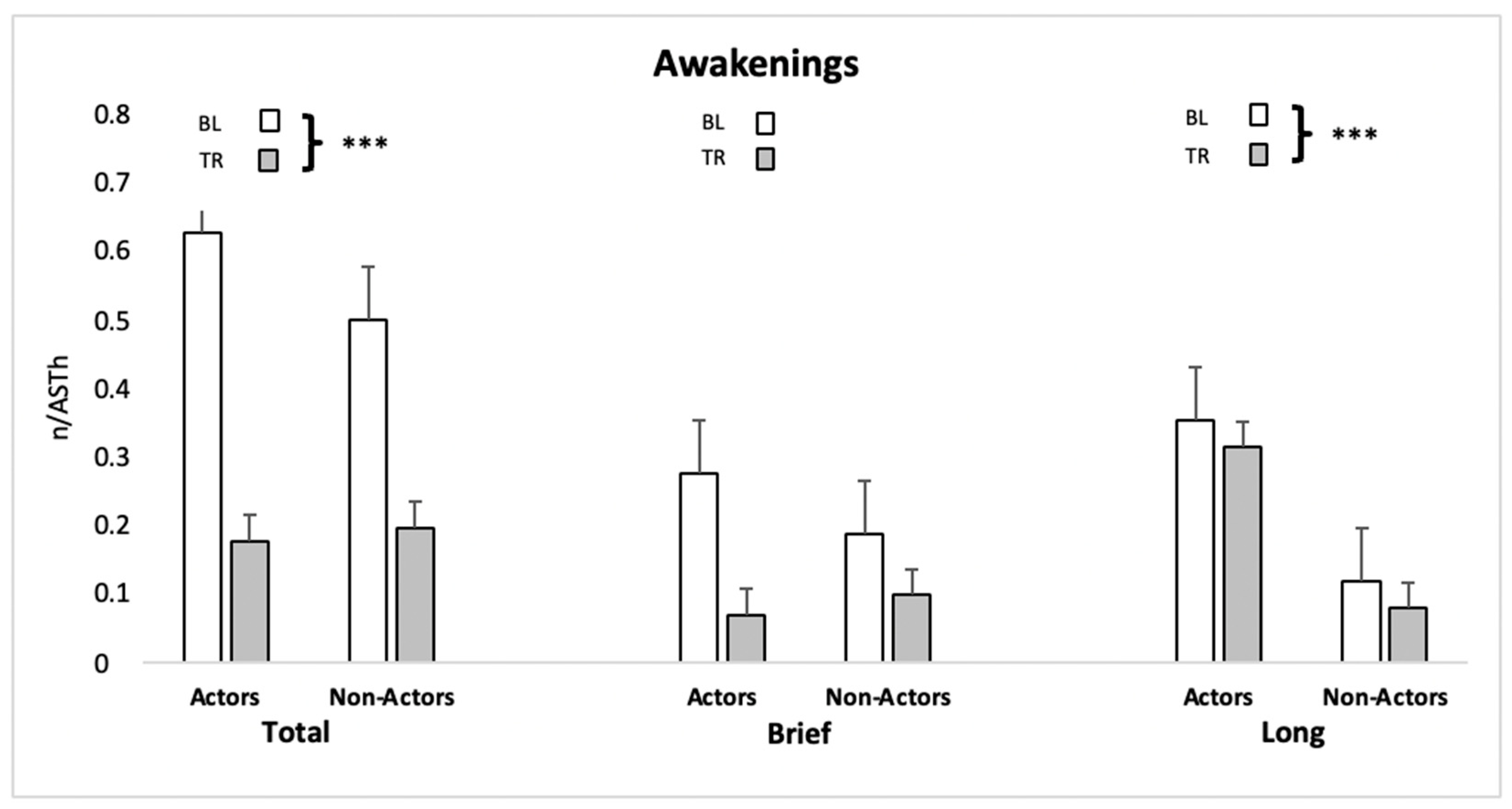
| Total Awakenings frequency | Actors | BL | 0.64 ± 0.51 | Condition | 19.94 | <0.001 | 0.231 |
| TR | 0.18 ± 0.23 | Interaction | 0.69 | 0.413 | 0.008 | ||
| non-Actors | BL | 0.51 ± 0.42 | Group | 0.146 | 0.706 | 0.004 | |
| TR | 0.20 ± 0.21 | ||||||
| Brief Awakenings frequency | Actors | BL | 0.28 ± 0.24 | Condition | 6.65 | 0.018 | 0.132 |
| TR | 0.07 ± 0.11 | Interaction | 0.097 | 0.335 | 0.019 | ||
| non-Actors | BL | 0.19 ± 0.25 | Group | 0.290 | 0.596 | 0.006 | |
| TR | 0.10 ± 0.15 | ||||||
| Long Awakenings frequency | Actors | BL | 0.36 ± 0.30 | Condition | 18.50 | <0.001 | 0.209 |
| TR | 0.12 ± 0.20 | Interaction | 0.001 | 0.967 | 0.000 | ||
| non-Actors | BL | 0.32 ± 0.28 | Group | 0.206 | 0.654 | 0.006 | |
| TR | 0.08 ± 0.12 | ||||||
| Awakenings mean duration (min) | Actors | BL | 3.76 ± 3.33 | Condition | 3.36 | 0.082 | 0.068 |
| TR | 3.39 ± 4.89 | Interaction | 2.29 | 0.146 | 0.047 | ||
| non-Actors | BL | 5.47 ± 5.13 | Group | 0.001 | 0.972 | 0.000 | |
| TR | 1.58 ± 1.86 | ||||||
| Descriptives | Statistics | ||||||
|---|---|---|---|---|---|---|---|
| Group | Condition | m ± sd | Effects | F | p | η2 | |
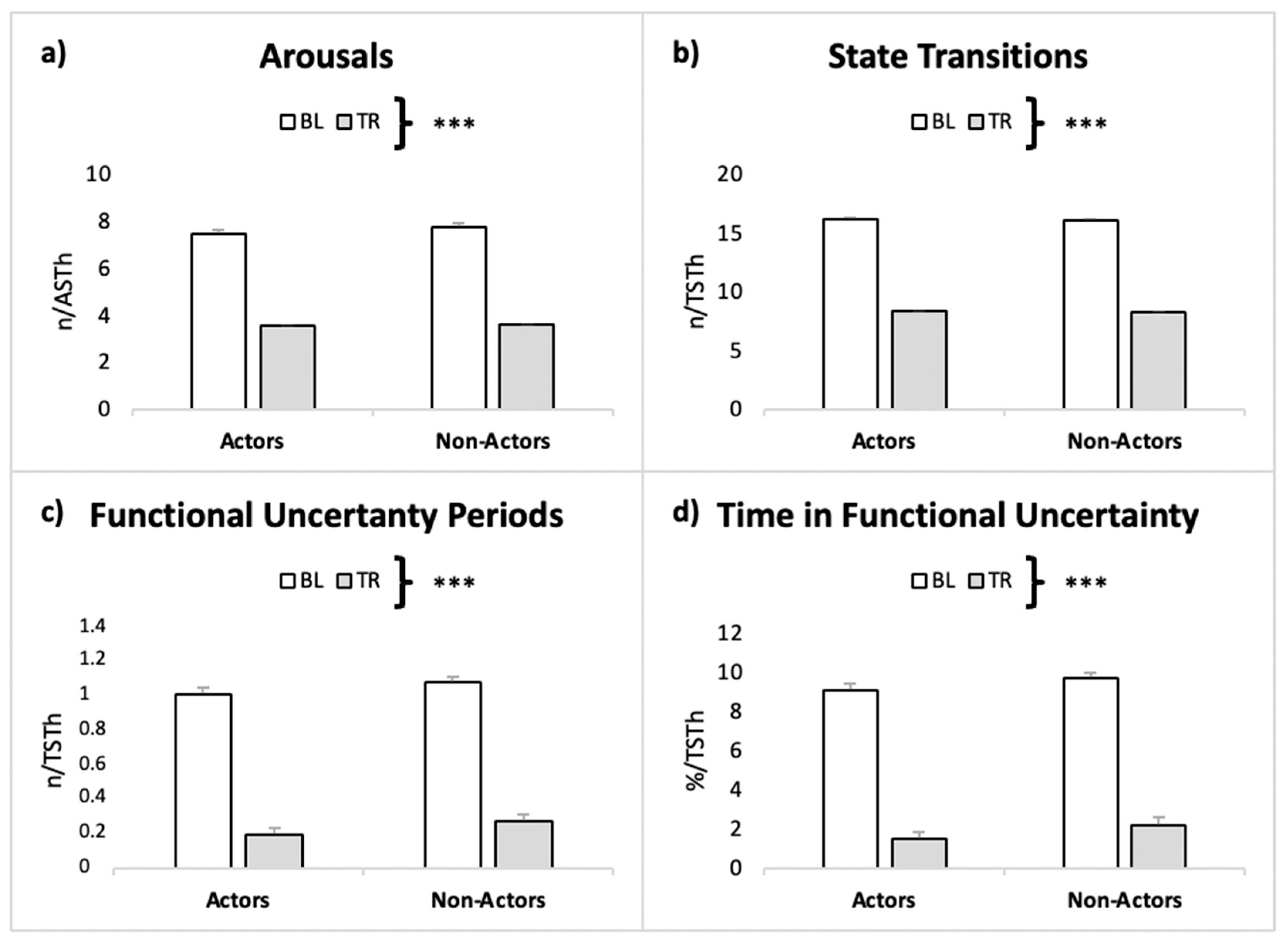
| Arousals frequency | Actors | BL | 7.49 ± 2.81 | Condition | 43.84 | <0.001 | 0.482 |
| TR | 3.55 ± 1.46 | Interaction | 0.040 | 0.842 | 0.000 | ||
| non-Actors | BL | 7.77 ± 2.39 | Group | 0.048 | 0.828 | 0.001 | |
| TR | 3.59 ± 1.94 | ||||||
| State Transitions frequency | Actors | BL | 16.2 ± 5.04 | Condition | 42.98 | <0.001 | 0.509 |
| TR | 8.31 ± 2.84 | Interaction | 0.001 | 0.972 | 0.001 | ||
| non-Actors | BL | 16.1 ± 4.17 | Group | 0.007 | 0.932 | 0.000 | |
| TR | 8.25 ± 3.86 | ||||||
| FU periods frequency | Actors | BL | 1.04 ± 0.62 | Condition | 40.1 | <0.001 | 0.485 |
| TR | 0.20 ± 0.23 | Interaction | 0.006 | 0.980 | 0.000 | ||
| non-Actors | BL | 1.11 ± 0.51 | Group | 0.271 | 0.608 | 0.004 | |
| TR | 0.28 ± 0.30 | ||||||
| TFU (%) | Actors | BL | 9.12 ± 5.72 | Condition | 36.6 | <0.001 | 0.470 |
| TR | 1.47 ± 1.79 | Interaction | 0.009 | 0.976 | 0.000 | ||
| non-Actors | BL | 9.79 ± 5.53 | Group | 0.301 | 0.589 | 0.004 | |
| TR | 2.21 ± 2.23 | ||||||
| FU periods mean duration (min) | Actors | BL | 5.12 ± 0.96 | Condition | 13.24 | 0.001 | 0.243 |
| TR | 2.96 ± 1.95 | Interaction | 0.375 | 0.547 | 0.007 | ||
| non-Actors | BL | 5.01 ± 0.86 | Group | 0.165 | 0.689 | 0.003 | |
| TR | 3.48 ± 2.46 | ||||||
| Descriptives | Statistics | ||||||
|---|---|---|---|---|---|---|---|
| Groups | Condition | m ± sd | Effects | F | p | η2 | |
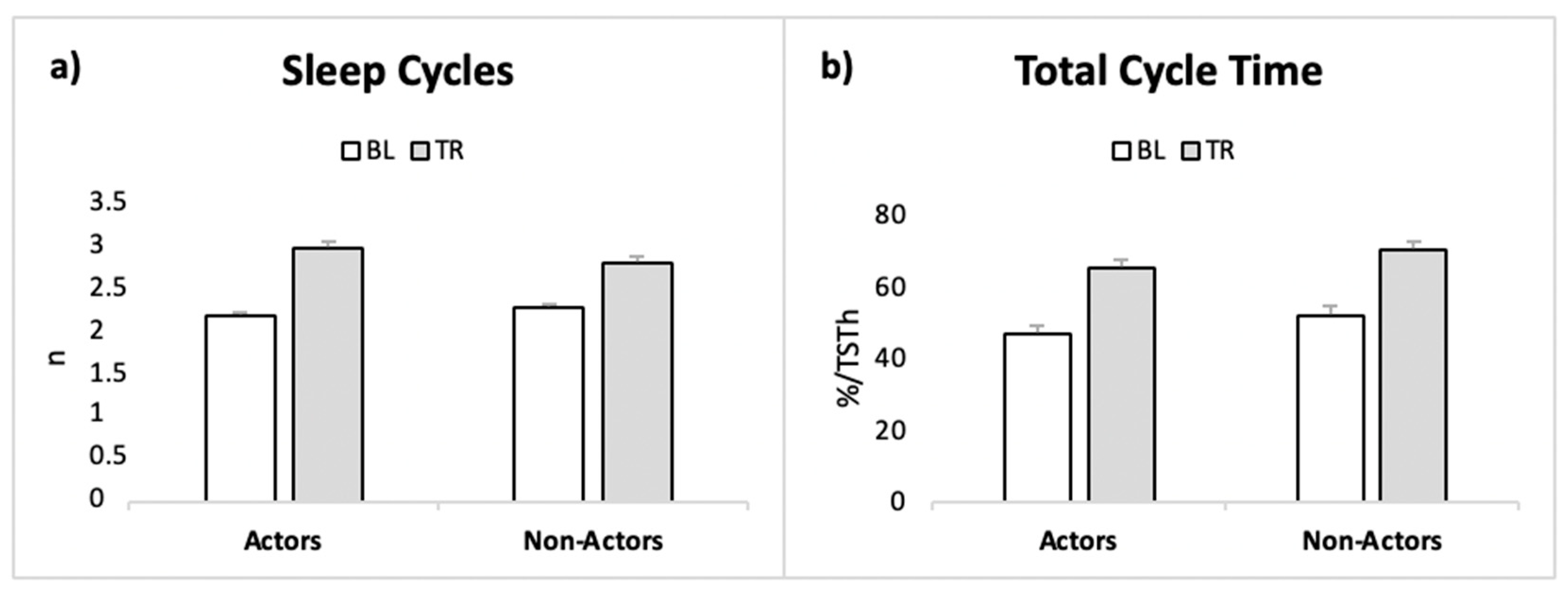
| number of Cycles | Actors | BL | 2.27 ± 1.42 | Condition | 6.32 | 0.021 | 0.078 |
| TR | 3.09 ± 1.22 | Interaction | 0.253 | 0.621 | 0.003 | ||
| non-Actors | BL | 2.36 ± 1.29 | Group | 0.010 | 0.920 | 0.000 | |
| TR | 2.91 ± 0.94 | ||||||
| TCT (%) | Actors | BL | 46.7 ± 19.9 | Condition | 6.60 | 0.018 | 0.152 |
| TR | 65 ± 21.9 | Interaction | 0.007 | 0.993 | 0.000 | ||
| non-Actors | BL | 51.9 ± 28.1 | Group | 0.642 | 0.432 | 0.012 | |
| TR | 70.1 ± 18.6 | ||||||
| Cycles mean duration (min) | Actors | BL | 91.4 ± 18.8 | Condition | 2.08 | 0.165 | 0.053 |
| TR | 91.7 ± 16.8 | Interaction | 1.96 | 0.177 | 0.050 | ||
| non-Actors | BL | 81.3 ± 33.7 | Group | 0.009 | 0.976 | 0.000 | |
| TR | 102 ± 17.1 | ||||||
| Descriptives | Statistics | ||||||
|---|---|---|---|---|---|---|---|
| Groups | Recall Phase | m ± sd | Effects | F | p | η2 | |
| Global Accuracy | Actors | immediate | 188 ± 31.3 | Recall phase | 0.060 | 0.808 | 0.000 |
| delayed | 182 ± 30 | Interaction | 1.70 | 0.206 | 0.004 | ||
| non-Actors | immediate | 167 ± 38.9 | Group | 1.39 | 0.252 | 0.061 | |
| delayed | 170 ± 34.8 | ||||||
| Synonyms | Actors | immediate | 11.1 ± 9.19 | Recall phase | 15.02 | <0.001 | 0.056 |
| delayed | 16.9 ± 12.8 | Interaction | 2.30 | 0.145 | 0.009 | ||
| non-Actors | immediate | 8.09 ± 4.74 | Group | 1.76 | 0.200 | 0.069 | |
| delayed | 10.6 ± 4.90 | ||||||
| Inversions | Actors | immediate | 0.81 ± 0.98 | Recall phase | 0.346 | 0.563 | 0.006 |
| delayed | 1.09 ± 1.76 | Interaction | 1.05 | 0.316 | 0.020 | ||
| non-Actors | immediate | 2.09 ± 3.94 | Group | 0.674 | 0.421 | 0.020 | |
| delayed | 1.09 ± 1.45 | ||||||
| Omissions | Actors | immediate | 19 ± 17.1 | Recall phase | 0.005 | 0.944 | 0.000 |
| delayed | 21.5 ± 32.8 | Interaction | 1.14 | 0.298 | 0.004 | ||
| non-Actors | immediate | 35.7 ± 26.5 | Group | 2.59 | 0.123 | 0.106 | |
| delayed | 32.8 ± 22.3 | ||||||
| Intrusions | Actors | immediate | 4.55 ± 4.91 | Recall phase | 0.525 | 0.477 | 0.004 |
| delayed | 4.09 ± 5.72 | Interaction | 1.76 | 0.199 | 0.015 | ||
| non-Actors | immediate | 2 ± 2.45 | Group | 0.907 | 0.352 | 0.035 | |
| delayed | 3.55 ± 2.77 | ||||||
| Errors | Actors | immediate | 1.18 ± 1.40 | Recall phase | 3.63 | 0.071 | 0.052 |
| delayed | 2.36 ± 2.87 | Interaction | 0.327 | 0.574 | 0.005 | ||
| non-Actors | immediate | 2 ± 1.61 | Group | 0.692 | 0.451 | 0.019 | |
| delayed | 2.64 ± 1.80 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conte, F.; De Rosa, O.; Albinni, B.; Mango, D.; Coppola, A.; Malloggi, S.; Giangrande, D.; Giganti, F.; Barbato, G.; Ficca, G. Learning Monologues at Bedtime Improves Sleep Quality in Actors and Non-Actors. Int. J. Environ. Res. Public Health 2022, 19, 11. https://doi.org/10.3390/ijerph19010011
Conte F, De Rosa O, Albinni B, Mango D, Coppola A, Malloggi S, Giangrande D, Giganti F, Barbato G, Ficca G. Learning Monologues at Bedtime Improves Sleep Quality in Actors and Non-Actors. International Journal of Environmental Research and Public Health. 2022; 19(1):11. https://doi.org/10.3390/ijerph19010011
Chicago/Turabian StyleConte, Francesca, Oreste De Rosa, Benedetta Albinni, Daniele Mango, Alessia Coppola, Serena Malloggi, Davide Giangrande, Fiorenza Giganti, Giuseppe Barbato, and Gianluca Ficca. 2022. "Learning Monologues at Bedtime Improves Sleep Quality in Actors and Non-Actors" International Journal of Environmental Research and Public Health 19, no. 1: 11. https://doi.org/10.3390/ijerph19010011
APA StyleConte, F., De Rosa, O., Albinni, B., Mango, D., Coppola, A., Malloggi, S., Giangrande, D., Giganti, F., Barbato, G., & Ficca, G. (2022). Learning Monologues at Bedtime Improves Sleep Quality in Actors and Non-Actors. International Journal of Environmental Research and Public Health, 19(1), 11. https://doi.org/10.3390/ijerph19010011