


Neurobehavioral Alterations from Noise Exposure in Animals: A Systematic Review


 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
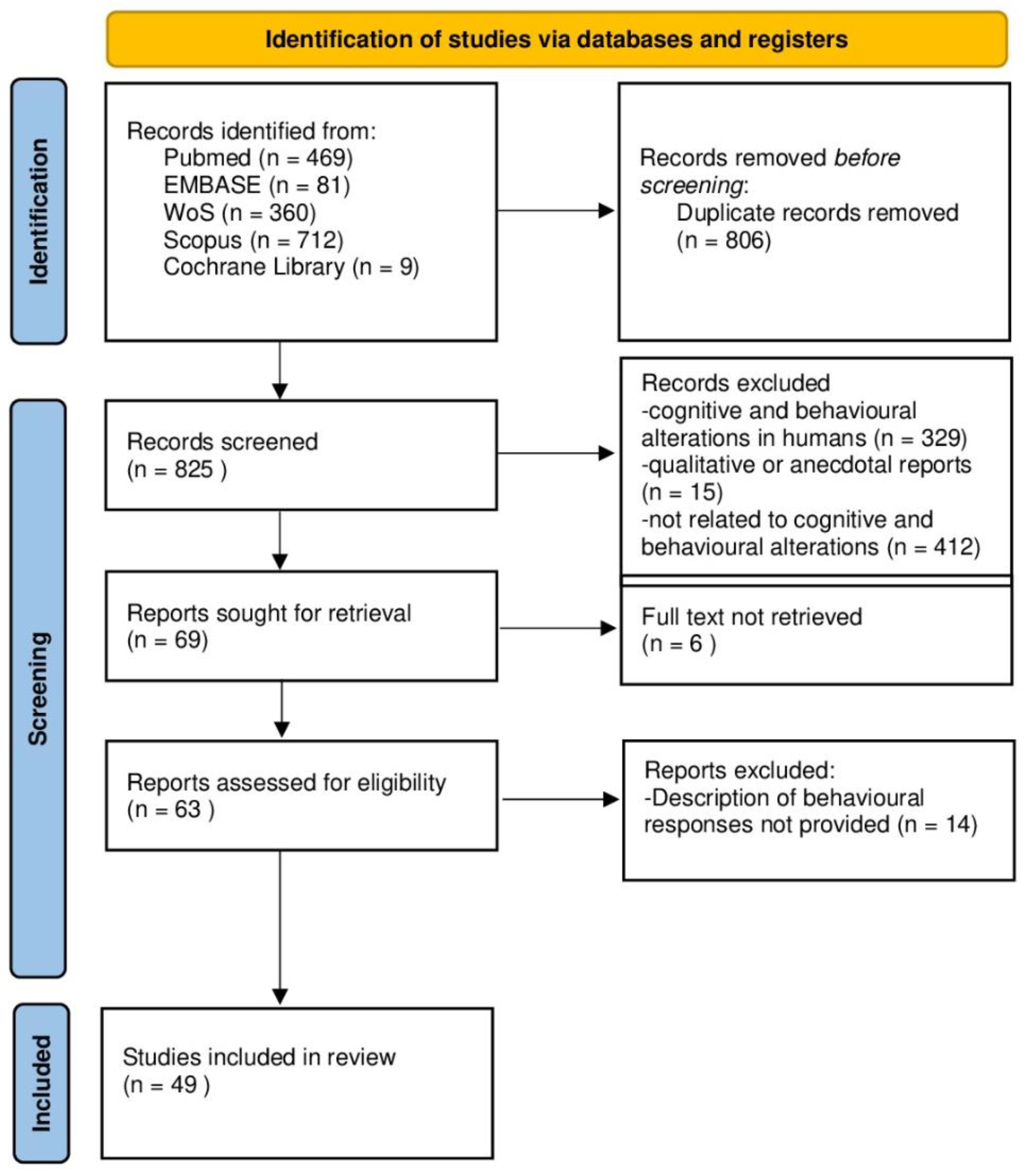
2. Materials and Methods
2.1. Literary Research
2.2. Quality Assessment
2.3. Eligibility and Inclusion Criteria
2.4. Exclusion Criteria
3. Results
3.1. Reviews
3.2. Experimental Studies
3.3. Other Articles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- González, A. What Does “Noise Pollution” Mean? J. Environ. Prot. 2014, 5, 340–350. [Google Scholar] [CrossRef] [Green Version]
- Hildebrand, J. Anthropogenic and natural sources of ambient noise in the ocean. Mar. Ecol. Prog. Ser. 2009, 395, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Frisk, G. Noiseonomics: The relationship between ambient noise levels in the sea and global economic trends. Sci. Rep. 2012, 2, 437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slabbekoorn, H.; Bouton, N.; van Opzeeland, I.; Coers, A.; Ten Cate, C.; Popper, A.N. A noisy spring: The impact of globally rising underwater sound levels on fish. Trends Ecol. Evol. 2010, 25, 419–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Franco, E.; Pierson, P.; Di Iorio, L.; Calò, A.; Cottalorda, J.; Derijard, B.; Di Franco, A.; Galvé, A.; Guibbolini, M.; Lebrun, J.; et al. Effects of marine noise pollution on Mediterranean fishes and invertebrates: A review. Mar. Pollut. Bull. 2020, 159, 111450. [Google Scholar] [CrossRef] [PubMed]
- Barber, J.R.; Crooks, K.R.; Fristrup, K.M. The costs of chronic noise exposure for terrestrial organisms. Trends Ecol. Evol. 2010, 25, 180–189. [Google Scholar] [CrossRef]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 15, 948–952. [Google Scholar] [CrossRef] [Green Version]
- Effects of Noise on Wildlife Nature Sounds. Available online: http://www.naturesounds.org/conservENW.html (accessed on 30 July 2021).
- Gianni, P. Gli effetti Del Rumore Subacqueo sui Mammiferi Marini. ACCOBAMS, n.4, 2002. Available online: http://www-9.unipv.it/cibra/edu_noise_ita.html (accessed on 30 July 2021).
- Zollinger, S.A.; Brumm, H. The Lombard effect. Curr. Biol. 2011, 21, R614–R615. [Google Scholar] [CrossRef] [Green Version]
- Gomes, D.G.E.; Page, R.A.; Geipel, I.; Taylor, R.C.; Ryan, M.J.; Halfwerk, W. Bats perceptually weight prey cues across sensory systems when hunting in noise. Science 2016, 353, 1277–1280. [Google Scholar] [CrossRef] [Green Version]
- Barton, B.T.; Hodge, M.E.; Speights, C.J.; Autrey, A.M.; Lashley, M.A.; Klink, V.P. Testing the AC/DC hypothesis: Rock and roll is noise pollution and weakens a trophic cascade. Ecol. Evol. 2018, 8, 7649–7656. [Google Scholar] [CrossRef]
- One Health. Available online: https://www.iss.it/one-health (accessed on 4 December 2022).
- Mucci, N.; Traversini, V.; Lulli, L.G.; Vimercati, L.; Rapisarda, V.; Galea, R.P.; De Sio, S.; Arcangeli, G. Neurobehavioral Alterations in Occupational Noise Exposure: A Systematic Review. Sustainability 2021, 13, 12224. [Google Scholar] [CrossRef]
- Prisma Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. The PRISMA Group. Prisma Statement. Evidence 2015, 7, e1000114. [Google Scholar]
- Torre, G.L.; Backhaus, I.; Mannocci, A. Rating for narrative reviews: Concept and development of the International Narrative Systematic Assessment tool. Senses Sci. 2015, 2, 31–35. [Google Scholar] [CrossRef]
- Shea, B.J.; Grimshaw, J.M.; Wells, G.A.; Boers, M.; Andersson, N.; Hamel, C.; Porter, A.C.; Tugwell, P.; Moher, D.; Bouter, L.M. Development of AMSTAR: A measurement tool to assess the methodological quality of systematic reviews. BMC Med. Res. Methodol. 2007, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Wells, G.A.; Shea, B.; O’Connel, D. The Newcastle-Ottawa Scale (NOS) for Assessing the Quality of Non-Randomized Studies in Meta-Analyses. 2009. Available online: http://www.ohrica/programs/clinical_epidemiology/oxford (accessed on 7 May 2020).
- Jadad, A.R.; Moore, R.A.; Carroll, D.; Jenkinson, C.; Reynolds, D.J.; Gavaghan, D.J.; Mc Quay, H.J. Assessing the quality of reports of randomized clinical trials: Is blinding necessary? Control. Clin. Trials 1996, 17, 1–12. [Google Scholar] [CrossRef]
- Shannon, G.; McKenna, M.F.; Angeloni, L.M.; Crooks, K.R.; Fristrup, K.M.; Brown, E.; Warner, K.A.; Nelson, M.D.; White, C.; Briggs, J.; et al. A synthesis of two decades of research documenting the effects of noise on wildlife. Biol. Rev. 2016, 91, 982–1005. [Google Scholar] [CrossRef]
- Kunc, H.P.; McLaughlin, K.E.; Schmidt, R. Aquatic noise pollution: Implications for individuals, populations, and ecosystems. Proc. R. Soc. B 2016, 283, 20160839. [Google Scholar] [CrossRef] [Green Version]
- De Soto, N.A. Peer-Reviewed Studies on the Effects of Anthropogenic Noise on Marine Invertebrates: From Scallop Larvae to Giant Squid. In The Effects of Noise on Aquatic Life II; Popper, A., Hawkins, A., Eds.; Advances in Experimental Medicine and Biology 875; Springer: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Cox, K.; Brennan, L.P.; Gerwing, T.G.; Dudas, S.E.; Juanes, F. Sound the alarm: A meta-analysis on the effect of aquatic noise on fish behavior and physiology. Glob Change Biol. 2018, 24, 3105–3116. [Google Scholar] [CrossRef]
- Nabi, G.; McLaughlin, R.W.; Hao, Y.; Wang, K.; Zeng, X.; Khan, S.; Wang, D. The possible effects of anthropogenic acoustic pollution on marine mammals’ reproduction: An emerging threat to animal extinction. Environ. Sci. Pollut. Res. 2018, 25, 19338–19345. [Google Scholar] [CrossRef]
- Peng, C.; Zhao, X.; Liu, G. Noise in the Sea and Its Impacts on Marine Organisms. Int. J. Environ. Res. Public Health 2015, 12, 12304–12323. [Google Scholar] [CrossRef]
- Popper, A.N.; Hawkins, A.D. An overview of fish bioacoustics and the impacts of anthropogenic sounds on fishes. J. Fish Biol. 2019, 94, 692–713. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Liu, M.; Dong, L.; Dong, J.; Wang, D. Potential impacts of shipping noise on Indo-Pacific humpback dolphins and implications for regulation and mitigation: A review. Integr. Zool. 2018, 13, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Samson, J.E.; Mooney, T.A.; Gussekloo, S.W.S.; Hanlon, R.T. A Brief Review of Cephalopod Behavioral Responses to Sound. In The Effects of Noise on Aquatic Life II: Advances in Experimental Medicine and Biology 875; Popper, A., Hawkins, A., Eds.; Springer: New York, NY, USA, 2016. [Google Scholar] [CrossRef] [Green Version]
- Kight, C.R.; Swaddle, J.P. How and why environmental noise impacts animals: An integrative, mechanistic review. Ecol. Lett. 2011, 14, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Mandel, R.; Whay, H.R.; Klement, E.; Nicol, C.J. Environmental enrichment of dairy cows and calves in indoor housing. J. Dairy Sci. 2016, 99, 1695–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akefe, I.O.; Ayo, J.O.; Sinkalu, V.O. Kaempferol and zinc gluconate mitigate neurobehavioraldecits and oxidative stress induced by noise exposure in Wistar rats. PLoS ONE 2006, 15, e0236251. [Google Scholar] [CrossRef]
- Criddle, M.W.; Godfrey, D.A.; Kaltenbach, J.A. Attenuation of noise-induced hyperactivity in the dorsal cochlear nucleus by pre-treatment with MK-801. Brain Res. 2018, 1682, 71–77. [Google Scholar] [CrossRef]
- Uran, S.L.; Aon-Bertolino, M.; Caceres, L.G.; Capani, F.; Guelman, L. Rat hippocampal alterations could underlie behavioral abnormalities induced by exposure to moderate noise levels. Brain Res. 2012, 1471, 1–12. [Google Scholar] [CrossRef]
- Koorpivaara, M.; Laapas, K.; Huhtinen, M.; Schöning, B. Overall. Dexmedetomidine oromucosal gel for noise associated acute anxiety and fear in dogs—A randomised, double-blind, placebo-controlled clinical study. Vet. Rec. 2017, 180, 356. [Google Scholar] [CrossRef] [Green Version]
- Mills, S.C.; Beldade, R.; Henry, L.; Laverty, D.; Nedelec, S.L.; Simpson, S.D.; Radford, A.N. Hormonal and behavioural effects of motorboat noise on wild coral reef fish. Environ. Pollut. 2020, 262, 114250. [Google Scholar] [CrossRef]
- Mikołajczak, J.; Borowski, S.; Marć-Pieńkowska, J.; Odrowąż-Sypniewska, G.; Bernacki, Z.; Siódmiak, J.; Szterk, P. Preliminary studies on the reaction of growing geese (Anser anser f. domestica) to the proximity of wind turbines. Pol. J. Vet. Sci. 2013, 16, 679–686. [Google Scholar] [CrossRef] [Green Version]
- Baltzer, J.; Maurer, N.; Schaffeld, T.; Ruser, A.; Schnitzler, J.G.; Siebert, U. Effect ranges of underwater noise from anchor vibration operations in the Wadden Sea. J. Sea Res. 2020, 162, 101912. [Google Scholar] [CrossRef]
- Hubert, J.; Neo, Y.Y.; Winter, H.V.; Slabbekoorn, H. The role of ambient sound levels, signal-to-noise ratio, and stimulus pulse rate on behavioural disturbance of seabass in a net pen. Behav. Process. 2020, 170, 103992. [Google Scholar] [CrossRef] [PubMed]
- Hastie, G.D.; Lepper, P.; McKnight, J.C.; Milne, R.; Russell DJ, F.; Thompson, D. Acoustic risk balancing by marine mammals: Anthropogenic noise can influence the foraging decisions by seals. J. Appl. Ecol. 2021, 58, 1854–1863. [Google Scholar] [CrossRef]
- Frouin-Mouy, H.; Tenorio-Hallé, L.; Thode, A.; Swartz, S.; Urbán, J. Using two drones to simultaneously monitor visual and acoustic behaviour of gray whales (Eschrichtius robustus) in Baja California, Mexico. J. Exp. Mar. Biol. Ecol. 2020, 525, 151321. [Google Scholar] [CrossRef]
- Grunst, M.L.; Grunst, A.S.; Pinxten, R.; Eens, M. Little parental response to anthropogenic noise in an urban songbird, but evidence for individual differences in sensitivity. Sci. Total Environ. 2021, 769. [Google Scholar] [CrossRef]
- Martin, M.; Gridley, T.; Elwen, S.H.; Charrier, I. Assessment of the impact of anthropogenic airborne noise on the behaviour of Cape fur seals during the breeding season in Namibia. J. Exp. Mar. Biol. Ecol. 2022, 550, 151721. [Google Scholar] [CrossRef]
- Leduc, A.O.; Costa, J.S.O.; Do Nascimento Silva, R.R.; Winandy, G.S.; De Araújo, C.B. Spatial cognitive abilities of a tide-pool fish show resilience to noise pollution. J. Ethol. 2021, 39, 225–234. [Google Scholar] [CrossRef]
- Issad, S.M.; Benhafri, N.; El Allali, K.; Farsi, H.; Ouali-Hassenaoui, S. Effects of prolonged night-time light exposure and traffic noise on the behavior and body temperature rhythmicity of the wild desert rodent, Gerbillus tarabuli. Chronobiol. Int. 2021, 38, 415–425. [Google Scholar] [CrossRef]
- Abdullah; Shabrina, A.; Khairil; Saputri, M.; Syafrianti, D.; Ala, N. The effect of antrophogenic noise on Sumatran Elephant’s anti-predator behavior in the Elephant Conservation Center. In Journal of Physics: IOP Conference Series Series, Proceedings of the the 1st Annual International Conference on Mathematics, Science and Technology Education, Kota Banda Aceh, Indonesia, 14–15 September 2019; IOP Publishing: Bristol, UK, 2020; Volume 1460, p. 012067. [Google Scholar] [CrossRef]
- Blanchett, M.K.S.; Finegan, S.; Atkinson, J. The Effects of Increasing Visitor and Noise Levels on Birds Within a Free-flight Aviary Examined Through Enclosure Use and Behavior. ABC 2020, 7, 49–69. [Google Scholar] [CrossRef]
- Senzaki, M.; Barber, J.R.; Phillips, J.N.; Carter, N.H.; Cooper, C.B.; Ditmer, M.A.; Fristrup, K.M.; McClure, C.J.W.; Mennitt, D.J.; Tyrrell, L.P.; et al. Francis. Sensory pollutants alter bird phenology and fitness across a continent. Nature 2020, 587, 605–609. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, L.; Cai, J.; Jiang, B.; Xu, W. Behavioral Response of Bean Goose (Anser fabalis) to Simulated Ship Noises at Lake. Animals 2022, 12, 465. [Google Scholar] [CrossRef] [PubMed]
- Amorim, M.C.P.; Vieira, M.; Meireles, G.; Novais, S.C.; Lemos, M.F.; Modesto, T.; Alves, D.; Zuazu, A.; Lopes, A.F.; Matos, A.B.; et al. Boat noise impacts Lusitanian toadfish breeding males and reproductive outcome. Sci. Total Environ. 2022, 830, 154735. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.J.O.; Isojunnoa, S.; Siegala, E.; Lamb, F.-P.A.; Kvadsheimc, P.H.; Cure, C. Behavioral responses to predatory sounds predict sensitivity of cetaceans to anthropogenic noise within a soundscape of fear. PNAS 2022, 119, e2114932119. [Google Scholar] [CrossRef]
- Van der Knaap, I.; Ashe, E.; Hannay, D.; Bergman, A.G.; Nielsen, K.A.; Lo, C.F.; Williams, R. Behavioural responses of wild Pacific salmon and herring to boat noise. Mar. Pollut. Bull. 2022, 174, 113257. [Google Scholar] [CrossRef]
- van der Knaap, I.; Slabbekoorn, H.; Moens, T.; Van den Eynde, D.; Reubens, J. Effects of pile driving sound on local movement of free-ranging Atlantic cod in the Belgian North Sea. Environ. Pollut. 2022, 300, 118913. [Google Scholar] [CrossRef] [PubMed]
- Pirotta, E.; Milor, R.; Quick, N.; Moretti, D.; Di Marzio, N.; Tyack, P.; Boyd, I.; Hastie, G. Vessel Noise Aects Beaked Whale Behavior: Results of a Dedicated Acoustic Response Study. PLoS ONE 2012, 7, e42535. [Google Scholar] [CrossRef] [Green Version]
- Williams, T.M.; Blackwell, S.B.; Tervo, O.; Garde, E.; Sinding, M.-H.; Richter, B.; HeideJørgensen, M.P. Physiological responses of narwhals to anthropogenic noise: A case study with seismic airguns and vessel traffic in the Arctic. Funct. Ecol. 2022, 36, 1–16. [Google Scholar] [CrossRef]
- Durbach, I.N.; Harris, C.M.; Martin, C.; Helble, T.A.; Henderson, E.E.; Ierley, G.; Thomas, L.; Martin, S.W. Changes in the Movement and Calling Behavior of Minke Whales (Balaenoptera acutorostrata) in Response to Navy Training. Front. Mar. Sci. 2021, 8, 660122. [Google Scholar] [CrossRef]
- Pellegrini, A.Y.; Romeu, B.; Ingram, S.N.; Daura-Jorge, F.G. Boat disturbance affects the acoustic behaviour of dolphins engaged in a rare foraging cooperation with fishers. Anim. Conserv. 2021, 24, 613–625. [Google Scholar] [CrossRef]
- Lara, R.A.; Vasconcelos, R.O. Impact of noise on development, physiological stress and behavioural patterns in larval zebrafsh. Sci. Rep. 2021, 11, 6615. [Google Scholar] [CrossRef]
- Manukyan, A.; Grigoryan, A.; Hunanyan, L.; Harutyunyanc, H.; Manukyand, M.V.; Mkrtchyan, V. Melkonyan, MAlfa2-adrenoblockers attenuate the elevated plasma cholesterol, anxiety levels and restore impaired spatial memory of rats under the chronic noise exposure. Sci. Total Environ. 2020, 740, 140390. [Google Scholar] [CrossRef] [PubMed]
- Mulders, W.H.A.M.; Robertson, D. Development of hyperactivity after acoustic trauma in the guinea pig Inferior colliculus. Hear. Res. 2013, 298, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Longenecker, R.J.; Galazyuk, A.V. Variable Effects of Acoustic Trauma on Behavioral and Neural Correlates of Tinnitus In Individual Animals. Front. Behav. Neurosci. 2016, 10, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landsberg, G.M.; Beck, A.; Lopez, A.; Deniaud, M.; Araujo, J.A.; Milgram, N.W. Dog-appeasing pheromone collars reduce sound-induced fear and anxiety in beagle dogs: A placebo-controlled study. Vet. Rec. 2015, 177, 260. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-K.; Do, Y. Wind Turbine Noise Behaviorally and Physiologically Changes Male Frogs. Biology 2022, 11, 516. [Google Scholar] [CrossRef]
- Sun, G.; Lin, X.; Yi, X.; Zhang, P.; Liu, R.; Fu, B.; Sun, Y.; Li, J.; Jiao, S.; Tian, T.; et al. Aircraft noise, like heat stress, causes cognitive impairments via similar mechanisms in male mice. Chemosphere 2021, 274, 129739. [Google Scholar] [CrossRef]
- Wieczerzak, K.B.; Patel, S.V.; MacNeil, H.; Scott, K.E.; Schormans, A.L.; Hayes, S.H.; Herrmann, B.; Brian, L. Allman. Differential Plasticity in Auditory and Prefrontal Cortices, and Cognitive-Behavioral Deficits Following Noise-Induced Hearing Loss. Neuroscience 2021, 455, 1–18. [Google Scholar] [CrossRef]
- McClure, C.J.; Ware, H.E.; Carlisle, J.; Kaltenecker, G.; Barber, J.R. An experimental investigation into the effects of traffic noise on distributions of birds: Avoiding the phantom road. Proc. R. Soc. B: Biol. Sci. 2013, 280, 20132290. [Google Scholar] [CrossRef]
- Slabbekoorn, H.; Peet, M. Birds sing at a higher pitch in urban noise. Nature 2003, 424, 267. [Google Scholar] [CrossRef]
- Kerbiriou, C.; Bas, Y.; Le Viol, I.; Lorrilliere, R.; Mougnot, J.; Julien, J.F. Evidence of suboscine song plasticity in response to traffic noise fluctuations and temporary road closures. Bioacoustics 2018, 27, 165–181. [Google Scholar] [CrossRef]
- Kareklas, K.; Wilson, J.; Kunc, H.P.; Arnott, G. Signal complexity communicates aggressive intent during contests, but the process is disrupted by noise. Biol. Letters. 2019, 15, 20180841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ETC; ICM. Pressures and their Effects Upon Europe’s Marine Environment, No Eionet Report ETC/ICM No 4/2019, European Topic Centre for Inland, Coastal and Marine Waters. 2019. Available online: https://www.eionet.europa.eu/etcs/etc-icm/products/etc-icm-report-4-2019-multiple-pressures-and-their-combined-effects-in-europes-seas (accessed on 30 July 2021).
- Weilgart, L. The impact of Ocean Noise Pollution on Fish and Invertebrates. OceanCare and Dalhousie University. 2018. Available online: https://www.oceancare.org/wp-content/uploads/2017/10/OceanNoise_FishInvertebrates_May2018.pdf (accessed on 30 July 2021).
- United Nations Convention on the Law of the Sea (UNCLOS); United Nations: New York, NY, USA, 1982; Volume VII, ISBN 978-90-04-19117-4.
- Commissione Europea. Linee Guida Per L’istituzione della Rete Natura 2000 Nell’ambiente Marino, Maggio 2007; European Comision: Brussels, Beligium, 2000; p. 108. Available online: www.ec.europa.eu/environment/nature/natura2000/marine (accessed on 30 July 2021).
- EEA Report. In Environmental Noise in Europe 2020; No 22/2019; European Environment Agency: Copenhagen, Denmark, 2020. [CrossRef]
- Jepson, P.D. Gas-bubble lesions in stranded cetaceans. Nature 2003, 425, 575. [Google Scholar] [CrossRef] [PubMed]
- Intergovernmental Panel on Climate Change. WGI, Quarto Rapporto di Valutazione 2007. Available online: https://www.ipcc.ch/site/assets/uploads/2021/03/ar4-wg1-spm-1.pdf (accessed on 30 July 2021).
- Heinrichs, S.C. Neurobehavioral consequences of stressor exposure in rodent models of epilepsy. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2010, 34, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Tai, S.K.; Huang, F.D.; Moochhala, S.; Khanna, S. Hippocampal theta state in relation to formalin nociception. Pain 2006, 1211, 29–42. [Google Scholar] [CrossRef]
- Nitz, D.A.; McNaughton, B.L. Hippocampal EEG and unit activity responses to modulation of serotonergic median raphe neurons in the freely behaving rat. Learn. Mem. 1999, 6, 153–167. [Google Scholar] [CrossRef]
- Koob, G.F. Corticotropin-releasing factor, norepinephrine, and stress. Biol. Psychiatry 1999, 46, 1167–1180. [Google Scholar] [CrossRef]
- Di, G.; Zhou, B.; Lin, Q. The effects of aircraft noise exposure on rat behavior and serum neurotransmitter expression. Noise Control Eng. J. 2011, 59, 514–518. [Google Scholar] [CrossRef]
- Aston-Jones, G.; Bloom, F.E. Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. J. Neurosci. 1981, 1, 876–886. [Google Scholar] [CrossRef] [Green Version]
- Aston-Jones, G.; Bloom, F.E. Nonrepinephrine-containing locus coeruleus neurons in behaving rats exhibit pronounced responses to non-noxious environmental stimuli. J. Neurosci. 1981, 1, 887–900. [Google Scholar] [CrossRef] [Green Version]
- Sara, S.J.; Vankov, A.; Herve, A. Locus coeruleus-evoked responses in behaving rats: A clue to the role of noradrenaline in memory. Brain. Res. Bull. 1994, 35, 457–465. [Google Scholar] [CrossRef]
- Walling, S.G.; Harley, C.W. Locus ceruleus activation initiates delayed synaptic potentiation of perforant path input to the dentate gyrus in awake rats: A novel betaadrenergic- and protein synthesis-dependent mammalian plasticity mechanism. J. Neurosci. 2004, 24, 598–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkowski, J.; Kubiak, P.; Aston-Jones, G. Locus coeruleus activity in monkey: Phasic and tonic changes are associated with altered vigilance. Brain. Res. Bull. 1994, 35, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Aston-Jones, G.; Rajkowski, J.; Kubiak, P. Conditioned responses of monkey locus coeruleus neurons anticipate acquisition of discriminative behavior in a vigilance task. Neuroscience 1997, 80, 697–715. [Google Scholar] [CrossRef] [PubMed]
- Kitchigina, V.; Vankov, A.; Harley, C.; Sara, S.J. Novelty-elicited, noradrenaline-dependent enhancement of excitability in the dentate gyrus. Eur. J. NeuroSci. 1997, 9, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Michaud, D.S.; McLean, J.; Keith, S.E.; Ferrarotto, C.; Hayley, S.; Khan, S.A.; Anisman, H.; Merali, Z. Differential impact of audiogenic stressors on Lewis and Fischer rats: Behavioral, neurochemical, and endocrine variations. Neuropsychopharmacology 2003, 28, 1068–1081. [Google Scholar] [CrossRef]
- Codocedo, J.F.; Inestrosa, N.C. Environmental control of microRNAs in the nervous system: Implications in plasticity and behavior. Neurosci. Biobehav. Rev. 2016, 60, 121–138. [Google Scholar] [CrossRef]
- Di, G.; Zheng, Y. Effects of high-speed railway noise on the synaptic ultrastructure and phosphorylated-CaMKII expression in the central nervous system of SD rats. Environ. Toxicol. Pharmacol. 2013, 35, 93–99. [Google Scholar] [CrossRef]
- Di, G.Q.; Xu, Y.Q. Influences of combined traffic noise on anxiety in mice. Sci. Total Environ. 2017, 579, 1439–1445. [Google Scholar] [CrossRef]
- Di, G.Q.; Liu, G.X.; Xu, Y.Q.; Kim, J.H. Effects of combined traffic noise on the synaptic ultrastructure and expresstions of p-CaMKII and NMDAR1 in the hippocampus of young SD rats. Environ. Sci. Pollut. Res. 2019, 26, 22030–22039. [Google Scholar] [CrossRef]
- Di, G.Q.; Qin, Z.Q. Influences of combined traffic noise on the ability of learning and memory in mice. Noise Health 2018, 20, 9–15. [Google Scholar]
- Raman, M.; Earnest, S.; Zhang, K.; Zhao, Y.; Cobb, M.H. TAO kinases mediate activation of p38 in response to DNA damage. EMBO J. 2007, 26, 2005–2014. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.F.; Wang, S.G. Human TAO kinase 1 induces apoptosis in SH-SY5Y cells. Cell Biol. Int 2008, 32, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Pienkowski, M.; Eggermont, J.J. Cortical tonotopic map plasticity and behavior. Neurosci. Biobehav. Rev. 2011, 35, 2117–2128. [Google Scholar] [CrossRef] [PubMed]
- Eggermont, J.J.; Komiya, H. Moderate noise trauma in juvenile cats results in profound cortical topographic map changes in adulthood. Hear. Res. 2000, 142, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Kujala, T.; Shtyrov, Y.; Winkler, I.; Saher, M.; Tervaniemi, M.; Sallinen, M.; Teder-Sälejärvi, W.; Alho, K.; Reinikainen, K.; Näätänen, R. Long-term exposure to noise impairs cortical sound processing and attention control. Psychophysiology 2004, 41, 875–881. [Google Scholar] [CrossRef]
- Brattico, E.; Kujala, T.; Tervaniemi, M.; Alku, P.; Ambrosi, L.; Monitillo, V. Longterm exposure to occupational noise alters the cortical organization of sound processing. Clin. Neurophysiol. 2005, 116, 190–203. [Google Scholar]
- Dutilleux, G. Anthropogenic outdoor sound and wildlife: It’s not just bioacoustics! In Proceedings of the Acoustics Conference, Nantes, France, 23–27 April 2012; pp. 2301–2306. Available online: Hal.archives-ouvertes.fr/docs/00/81/07/95/PDF/hal-00810795.pdf (accessed on 30 July 2021).
- EEA Report. In Quiet Areas in Europe—The Environment Unaffected by Noise Pollution; No 14/2016; European Environment Agency: Copenhagen, Denmark, 2016; ISBN 978-92-9213-745-8. [CrossRef]


{kind=link}
| Population | Animal with No Differences in Species, Habitat or Geolocation. |
|---|---|
| Interventions | Noise exposure from any source |
| Comparison | N.A. |
| Outcomes | Neurobehavioral or neurological alterations |
| First Author. | Year | Country | Type of Study | Species | Alterations | Score |
|---|---|---|---|---|---|---|
| Abdullah | 2020 | Indonesia | observational study | Elephants | altered anti-predatory’reaction | N.6 |
| Akefe | 2020 | Nigeria | experimental study | Rats | learning, short-term memory, sensorimotor reflex | J.2 |
| Amorim | 2022 | Portugal | Case-control | Fish | Behavioral and reproductive responses | N.5 |
| Baltzer | 2020 | Wadden sea | experimental study | marine mammals, fishes | altered movements, swimming speed, anti-predatory reaction | n.a. |
| Blanchett | 2020 | USA | observational study | Birds | aggression, pacing, nesting etc | N.6 |
| Codocedo | 2016 | Australia/Chile | narrative review | mice, rats | anhedonia, anxiety, social-avoidance behaviors | I.6 |
| Cox | 2017 | Canada | meta-analysis | Fishes | complex movements and swimming abilities | A.6 |
| Criddle | 2018 | USA | experimental study | Hamsters | hyperactivity | J.2 |
| De Soto | 2016 | Spain | narrative review | marine invertebrates | altered movements, swimming speed, metabolic parameters | I.5 |
| Di Franco | 2020 | Italy/France | systematic review | marine invertebrates, fishes | altered movements, swimming speed, anti-predatory reaction | A.4 |
| Durbach | 2021 | UK | Observational study | Whales | Behavior responses | N.6 |
| Frouin-Mouy | 2020 | Mexico | experimental study | Whales | resting, interaction mother-calf | n.a. |
| Gang | 2021 | China | Case-control | Mice | Stress response, cognitive capacities, neuroinflammation | N.5 |
| Grunst | 2021 | Belgium | Experimental study | birds | Parental behaviors | J.3 |
| Hastie | 2021 | UK | Experimental study | Grey seals | Foraging behavior | n.a. |
| Heinrichs | 2010 | USA | narrative review | Rodents | anxiety, hyperactivity | I.5 |
| Hubert | 2020 | North sea | experimental study | Fishes | changed swimming | n.a. |
| Issad | 2021 | Algeria | Experimental study | Gerbils | Circadian rhythm and anxious behavior | n.a. |
| Kight | 2011 | USA | narrative review | rats, zebra | cognition, sleep | I.5 |
| Koorpivaara | 2017 | USA | experimental study | Dogs | anxiety, fear | J.4 |
| Kunc | 2016 | Uk | narrative review | marine species | aggression, hunting, movements, anti-predatory reaction | I.5 |
| Landsberg | 2015 | Canada | case-control | Dogs | anxiety, fear | N.6 |
| Lara | 2021 | China | Case-control | Larval zebrafish | stronger dark avoidance, scotophobia, movements and swimming alterations | N.5 |
| Leduc | 2021 | Brazil | Experimental study | Fish | Cognitive performance | J.2 |
| Li | 2018 | Indo-Pacific sea | narrative review | Dolphins | altered movements and vocals | I.4 |
| Longenecker | 2016 | USA | cohort study | Mice | hyperactivity | N.6 |
| Mandel | 2016 | Israel, Uk | narrative review | cows, calves | various | I.4 |
| Manukyan | 2020 | Armenia | case-control | Rats | anxiety, memory | N.6 |
| Martin | 2022 | France | Experimental study | Cape fur seals | Behavioral responses | J.2 |
| Mikolajczak | 2013 | Poland | experimental study | Geese | movements, stress | J.2 |
| Miller | 2022 | UK | Observational study | Cetaceans | Foraging behavior | N.a. |
| Mills | 2020 | Polynesia | experimental study | Fishes | hiding, distance, aggression | J.2 |
| Mulders | 2013 | Australia | case-control | Pig | hyperactivity | N.5 |
| Nabi | 2018 | China/USA | narrative review | marine mammals | masking, altered reproduction | I.5 |
| Park | 2022 | Korea | Case-control | Frogs | behavioral–physiological–immunological response | N.6 |
| Pellegrini | 2020 | Brazil | Observational study | Dolphins | Foraging behavior | N.6 |
| Peng | 2015 | China | narrative review | marine species | nesting, aggression, anti-predatory reaction | I.6 |
| Pienkowski | 2011 | Canada | narrative review | rats, cats | cortical plasticity | I.4 |
| Pirotta | 2012 | USA | case-control | Whales | foraging, movements | N.6 |
| Popper | 2019 | USA/Uk | narrative review | Fishes | impairment of spawning, interference with foraging, disruption in migration-habitat selection | I.4 |
| Samson | 2016 | USA/Netherland | narrative review | cephalopods | escape, inking, altered speed | I.3 |
| Senzaki | 2020 | USA | Observational study | Birds | Reproductive behaviors | N.6 |
| Shannon | 2016 | USA | systematic review | Wildlife | vocals, movements, foraging, escape, vigilance, mating | A.5 |
| Uran | 2012 | Argentina | experimental study | Rats | recognition, memory | J.2 |
| Van der Knapp | 2021 | Netherlands | Observational study | Fish | Behavioral responses | N.5 |
| Van der knapp(b) | 2021 | Netherlands | Observational study | Fish | Movement behavior | N.6 |
| Wang | 2022 | China | Observational study | waterbirds | Flight pattern | N.6 |
| Wieczerzak | 2021 | Canada | Cohort study | Mice | Cognitive behavior | N.6 |
| Williams | 2022 | USA | Case-control | narwhales | Locomotor reactions | N.6 |
| First Author | Included Articles | Level’ Noise | Results |
|---|---|---|---|
| Codocedo | narrative | not specified | In rats, noise exposure for 24 h generates a decrease in several miRNAs, including miR-183, leading to adecrease in the level of the target TaoK1, which participates in the activation of the MAPK pathway and the induction of cell apoptosis |
| Cox | 42 | not specified | Increased hearing thresholds and cortisol levels were associated with an increase in stress-related hormones, and suggest that anthropogenic noise has the potential to cause both short- and long-term physiological effects |
| De Soto | 15 | 157/136–162/156–168 dB re 1 μPa | noise interferes with growth larvae, metabolism, reproductive rates, changes in swimming and movements |
| Di franco | 57 | not specified | acute and chronic marine noise can cause a wide variety of effects on marine invertebrates and vertebrates, such as swimming and gregarious patterns, anti-predator responses, mating and spawning patterns, auditory damage, communication masking, changes in habitat use, migration and displacement, stress-related physiological responses |
| Heinrichs | narrative | 120 dB–12 kHz | mechanisms that can induce hyperactivity in animals exposed to stressors, such as loud noises, are related to hippocampal changes, in the locus coerulus or to activation of adrenocortical hormones. |
| Kight | narrative | 65–95–110 dB | noise stressed animals are not able to reproduce species-appropriate vocalisations, they do spatial errors and stress during pregnancy but noise might act as a beneficialstimulant of brain activity, such as white noise during sleep |
| Kunc | narrative | not specified | Noise may also negatively affect the social structure between pairs and groups, can impede defence against predators, reduce the ability to maintain territories or alter the reproductive behavior |
| Li | narrative | pulse with sound exposure levels (SELs) > 183 dB re: 1 μPa2 and nonpulses > 195 dB re: 1 μPa2s | dolphins with vessel noise change their fluke, rate, heading, dive depth and reduced their sounds |
| Mandel | narrative | not specified | in cows white noise or classical music decreases stress level |
| Nabi | narrative | not specified | masking can compromise reproduction, mother-offspring bonding, foraging and survival because animals are unable to interpret and respond to mating calls, offspring calls, prey sounds or predator sound |
| Peng | narrative | 119–250 dB re 1 μPa | the effects of anthropogenic noise on marine organisms are dependent on the species investigated and both the levels of impulsive and stationary noise |
| Pienkowski | narrative | 68–72 dB spl | sounds can lead to a reorganization of auditory cortex not unlike that following restricted hearing loss but different from that learning-induced |
| Popper | narrative | 20–50 Hz (bulk), 180 to 200 dB re 1 μPa2 s−1 (pile drivers), <1 Hz (vessel) | change in behavior from small and short-duration movements to changes in migration routes and leaving a feeding or breeding site; decrease in detectability ofbiologically relevant sounds (e.g., sounds of predators and prey, sounds of conspecifics, acoustic cues used for orientation) |
| Samson | narrative | 20–1000 Hz | in cephalopods, reactions considered to be escape and/or startle behavior (blanching, jetting, inking) mostly occurred at low frequencies and high sound levels |
| Shannon | 188 | 52 and 68 dBA SPL re 20 μPa (terrestrial)/67–195 dB SPL re 1 μPa (acquatic) | noise cause increased stress levels, decreased reproductive efficiency, impacted the vocal behavior and reduced the foraging efficiency |
| First Author | Sample | Level of Noise | Aim | Lenght of Study | Results |
|---|---|---|---|---|---|
| Abdullah | 2 | 20–75 dB | exposure to various noise for 15 min | 3 repetitions in each day for 5 days | noise interferes with prey perceptions of predators |
| Akefe | 30 | 100 dB | exposed to noise, with or not kaempferol + zinc gluconate | 48 days | noise interferes with oxidative stress |
| Amorim | 16 | 104–140 dB re. 1 μPa | impacts of boat noise exposure in the reproductive success of wild toadfish | 2 weeks | Noise affected reproductive success by decreasing the likelihood of receiving eggs, the number of live eggs and increasing the number of dead eggs |
| Baltzer | not specified | 120–99 dB re 1 μPa2s | effects of underwater noise on marine mammals | 1 day | anchor pipe vibration embedment noise might induce a behavioral reaction (changes in movements) |
| Blanchett | 98 | 51.5–66.6 dB | associations between visitor numbers, noise levels and stress or critical behavior | 12 days | lack of association between visitor numbers and stress or critical behavior |
| Criddle | 24 | 85–115 dB | NMDA receptor blocker and sound exposure | 4 h + 28 days | treated animals show lower hyperactivity |
| Durbach | Not specified | approximately 3 kHz and a nominal source level of 235 dB re 1 µPa | investigate the effect of sonar activity on movement behaviors | 3–4 days for 3 years | Faster and more directed movement during sonar exposure; animals were more likely to cease calling during exposure |
| Frouin-Mouy | 2 | 94.8–110.2 dB re 1 μPa | measuring the underwater source levels, behavioral vocal and non-vocal marine mammal signals | 1 month | noise can interfere communications between group |
| Gang | 120 | mean sound pressure level of 72 dB (A) | Associations between aircraft noise and cognitive functions | 2 h daily for 4 days | Changes in spatial recognition memory |
| Grunst | 34 pairs | 60 dB | altered parental behaviors in response to consistent freeway noise and a diverse anthropogenic noise | 2 weeks | no population-level changes in nestling provisioning behavior during noise but individual differences in noise sensitivity |
| Hastie | 5 | 148 dB re 1 µPa | measuring the relative influence of a sound (silence, pile driving, and a tidal turbine) on decision-making and foraging success in grey seals | 8 days | Foraging success was significantly reduced (16%–28% lower) when the speaker was located at the Low Density prey patch |
| Hubert | 64 | mean SPLs 128.3 or 119.0 dB re 1 μPa | exposed seabass to different impulsive sound treatments (pulse level, elevated background level) | 3 sound treatment in each day for 2 days | upon sound exposure, fishes increased their swimming depth |
| Issad | 32 | 80 dB | Effects of light and noise pollution on body temperature and anxious behavior | 3–4 weeks | significant decrease in the number of line crossings and time spent in the open field test. |
| Koorpivaara | 182 | not specified | dexmedetomidine for noise-associated acute anxiety and fear in dogs | 3 months | noise can caused hyperactivity by locus coerulus’activation |
| Landsberg | 24 | average 83.9 dB | two treatment groups (DAP and placebo) in response to a thunderstorm recording | a week | pheromones reduce anxiety and fear by noise |
| Lara | Not specified | 130 and 150 dB re 1 μPa | Shipping activity can altered fish’behavior | 5 days | continuous noise can increase dark avoidance in anxiety-related dark/light preference test and impaired spontaneous alternation behavior |
| Leduc | 32 | 45–100 dB | Noise can reduce the available cognitive processing capacity | 3 weeks | fish exposed to noise playbacks require additional time to reach this target and reduce exploratory behavior |
| Longenecker | 16 | 116 dB | relationship between tinnitus, hearing loss, hyperactivity and bursting activity post noise trauma | 1 h | noise increase tinnitus and hyperactivity |
| Manukyan | 24 | 91 dB | monitoring hdl, ldl, cholesterol, cognitive functions post noise exposure | 60 days | chronic noise altered behavioral activity, delay in movement and orientation, increased anxiety, deficit spatial memory |
| Martin | 35 groups (369 individuals) | 60.9–64.4 (low)/64.4–70.5 (medium)/70.5–80 dB re 20 μPa RMS SPL (high) | Effect of car and boat noise on marine mammals behavior | 1 month | detriment of vital activities such as resting and nursing that decreased considerably (from 5.9 to 45% decrease) |
| Mikolajczak | 40 | 94–104 dB | effect of noise by wind turbines on the stress parameters (cortisol) | 17 weeks | lower activity, some disturbing changes in behavior, increased cortisol |
| Miller | 43 | 1–4 kHz | Effect of sonar noise on foraging | 13 h | whales ceased foraging completely during killer whale and sonar exposures |
| Mills | 28/20 | 120–70 dB re 1 μPa2s/Hz | short-longer effect motorboat-noise playback on the behavior, cortisol, androgens of anemonefish | 30 min/48 h | in short term, hiding aggression, androgen level increased |
| Mulders | 24 | 20–120 dB | monitoring hyperactivity post noise exposure | 2 weeks | hyperactivity in the colliculus begins at some time between 4 and 12 h post trauma |
| Park | 27/24 | 41.3–57.60 dB | Effects of wind turbine on frog’s behavior | 2 days | Call rate increased after 1 h of exposure |
| Pellegrini | 122 groups | 180 dB re: 1 lPa V−1 | Effects of boat noise on foraging | 9 months | cooperative foraging may potentially be reducedor interrupted by the presence of boats, in response to the number, type and speed, indicating a behavioral change and acoustic masking |
| Pirotta | 32 | 50–200 dB re 1 μPa2s/Hz | How vessel noise influenced foraging behavior | 5 days | ship noise caused a significant change in whale behavior up to at least 5.2 km away from the vessel. |
| Senzaki | 142 species (58,506 nest) | Not specified | Effect of light and noise on reproductive success | 14 years | Closed-habitat, but not open-habitat, birds also tended to experience a decline in clutch size with noise exposure |
| Uran | 30 | 95–97 dB SPL | Monitoring hippocampal-related behavioral alterations | 15–30–45 postnatal day | moderate intensity can changed hippocampus, with observed behavioral effects |
| Van der Knapp | 49–250 | 123–140 dB (re 1 μPa) | Effect of boat noise on behavior | 6 months | in presence of boat noise) fishes spent more time in behaviorsconsidered to be a response to predators |
| Van der knapp(b) | 14 | 114–138 dB (re 1 μPa) | Effect of wind turbine on movement behavior of free swimming | 4 months | cod did not increase their net movementactivity, but moved closer to the scour-bed (i.e., hard substrate), surrounding their nearest turbine, |
| Wang | 60 | 60–100 dB | investigate the effects of ship noise on foraging, vigilance and flight behaviors | 1 month | As the noise level increased, foraging behavior decreased and vigilance and flight behaviors increased, particularly above 70 dB |
| Wieczerzak | 10/11 | 40 Hz | Investigated neural plasticity in the auditory and prefrontal cortices in the days following noise exposure | 1 month | noise exposure impaired spatial learning and reference memory |
| Williams | 13 | 241 dB re 1 μPa-m | Investigated reactions to anthropogenic noise by this deep-diving cetacean | 5 years | movement from surface to depth (descent) was often more gradual for control dives than for noise exposed dives which showed shorter, more rapid ‘directed’ descents |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arcangeli, G.; Lulli, L.G.; Traversini, V.; De Sio, S.; Cannizzaro, E.; Galea, R.P.; Mucci, N. Neurobehavioral Alterations from Noise Exposure in Animals: A Systematic Review. Int. J. Environ. Res. Public Health 2023, 20, 591. https://doi.org/10.3390/ijerph20010591
Arcangeli G, Lulli LG, Traversini V, De Sio S, Cannizzaro E, Galea RP, Mucci N. Neurobehavioral Alterations from Noise Exposure in Animals: A Systematic Review. International Journal of Environmental Research and Public Health. 2023; 20(1):591. https://doi.org/10.3390/ijerph20010591
Chicago/Turabian StyleArcangeli, Giulio, Lucrezia Ginevra Lulli, Veronica Traversini, Simone De Sio, Emanuele Cannizzaro, Raymond Paul Galea, and Nicola Mucci. 2023. "Neurobehavioral Alterations from Noise Exposure in Animals: A Systematic Review" International Journal of Environmental Research and Public Health 20, no. 1: 591. https://doi.org/10.3390/ijerph20010591
APA StyleArcangeli, G., Lulli, L. G., Traversini, V., De Sio, S., Cannizzaro, E., Galea, R. P., & Mucci, N. (2023). Neurobehavioral Alterations from Noise Exposure in Animals: A Systematic Review. International Journal of Environmental Research and Public Health, 20(1), 591. https://doi.org/10.3390/ijerph20010591