
Prevalence of Multidrug-Resistant Bacteria (Enteropathogens) Recovered from a Blend of Pig Manure and Pinewood Saw Dust during Anaerobic Co-Digestion in a Steel Biodigester


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Determination of the Counts of Viable Bacteria
2.3. Isolation, Biochemical Characterisation and Storage of the Bacteria
2.4. Determination of Antibiotic Resistance Phenotypes
2.5. Calculating Multiple Antibiotic Resistance (MAR) Index and Presentation of Their Resistance Patterns
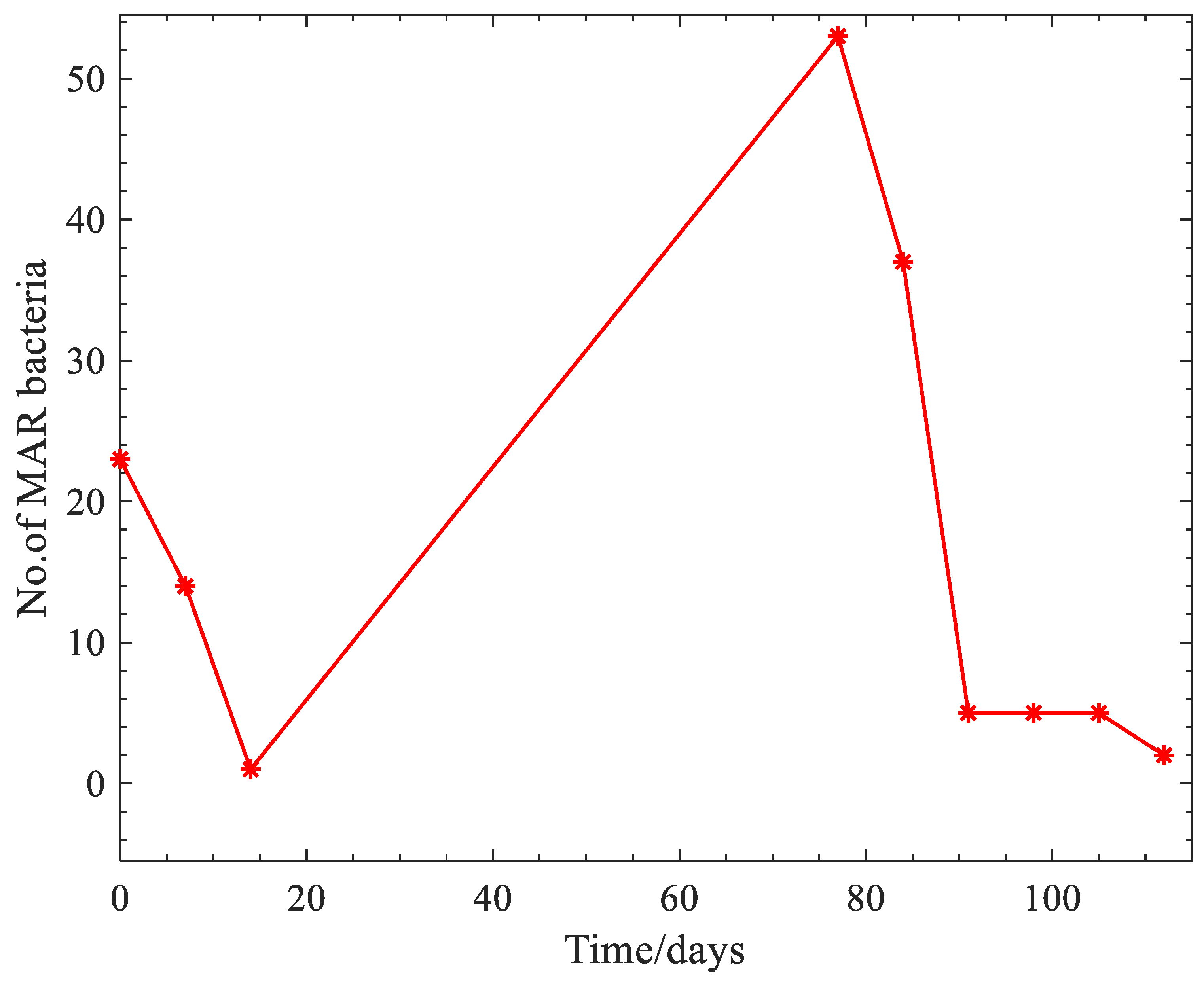
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mmbengwa, V.; Nyhodo, B.; Myeki, L.; Ngethu, X.X.; van Schalkwyk, H. Communal livestock farming in South Africa: Does this farming system create jobs for poverty stricken rural areas? Sylwan 2015, 159, 176–192. [Google Scholar]
- Scholtz, N.M.; van Ryssen, J.B.J.; Meissner, H.H.; Laker, M.C. A South African perspective on livestock production in relation to greenhouse gases and water usage. S. Afr. J. Anim. Sci. 2013, 43, 247. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Yu, Z.; Gao, H.; Yan, X.; Chang, J.; Wang, C.; Hu, J.; Zhang, L. Chemical structures and characteristics of animal manures and composts during composting and assessment of maturity indices. PLoS ONE 2017, 12, e0178110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngumbela, X.G. Unique Challenges of the Poverty Dilemma in the Eastern Cape Province of South Africa. Afr. J. Public Aff. 2021, 12, 114–131. [Google Scholar]
- Ngumbela, X.G.; Khalema, E.N.; Nzimakwe, T.I. Local worlds: Vulnerability and food insecurity in the Eastern Cape province of South Africa. Jàmbá J. Disaster Risk Stud. 2020, 12, a830. [Google Scholar] [CrossRef]
- Kostic, B.; Stevanovic, G.; Lutovac, M.; Lutovac, B.; Ketin, S.; Biocanin, R. Animal manure and environment. Fresenius Environ. Bull. 2020, 29, 1289–1296. [Google Scholar]
- Blaiotta, G.; Di Cerbo, A.; Murru, N.; Coppola, R.; Aponte, M. Persistence of bacterial indicators and zoonotic pathogens in contaminated cattle wastes. BMC Microbiol. 2016, 16, 87. [Google Scholar] [CrossRef] [Green Version]
- Noll, M.; Kleta, S.; Al Dahouk, S. Antibiotic susceptibility of 259 Listeria monocytogenes strains isolated from food, food-processing plants and human samples in Germany. J. Infect. Public Health 2018, 11, 572–577. [Google Scholar] [CrossRef]
- Van, T.T.H.; Yidana, Z.; Smooker, P.M.; Coloe, P.J. Antibiotic use in food animals worldwide, with a focus on Africa: Pluses and minuses. J. Glob. Antimicrob. Resist. 2020, 20, 170–177. [Google Scholar] [CrossRef]
- Meissner, H.H.; Scholtz, M.M.; Palmer, A.R. Sustainability of the South African Livestock Sector towards 2050 Part 1: Worth and impact of the sector. S. Afr. J. Anim. Sci. 2013, 43, 283–297. [Google Scholar] [CrossRef]
- Doidge, C.; Ruston, A.; Lovatt, F.; Hudson, C.; King, L.; Kaler, J. Farmers’ Perceptions of Preventing Antibiotic Resistance on Sheep and Beef Farms: Risk, Responsibility, and Action. Front. Vet. Sci. 2020, 7, 524. [Google Scholar] [CrossRef] [PubMed]
- Finley, R.L.; Collignon, P.; Joakim Larsson, D.G.; McEwen, S.A.; Li, X.-Z.; Gaze, W.H.; Reid-Smith, R.; Timinouni, M.; Graham, D.W.; Topp, E. The Scourge of Antibiotic Resistance: The Important Role of the Environment. Clin. Infect. Dis. 2013, 57, 704–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhao, Z.; Orfe, L.; Subbiah, M.; Call, D.R. Soil-borne reservoirs of antibiotic-resistant bacteria are established following therapeutic treatment of dairy calves. Environ. Microbiol. 2016, 18, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Dadgostar, P. Antimicrobial Resistance: Implications and Costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, N.D.; Temkin, E.; Carmeli, Y. The negative impact of antibiotic resistance. Clin. Microbiol. Infect. 2016, 22, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Mnyambwa, N.P.; Mahende, C.; Wilfred, A.; Sandi, E.; Mgina, N.; Lubinza, C.; Kahwa, A.; Petrucka, P.; Mfinanga, S.; Ngadaya, E.; et al. Antibiotic Susceptibility Patterns of Bacterial Isolates from Routine Clinical Specimens from Referral Hospitals in Tanzania: A Prospective Hospital-Based Observational Study. Infect. Drug Resist. 2021, 14, 869–878. [Google Scholar] [CrossRef]
- Bayot, M.L.; Bragg, B.N. Antimicrobial Susceptibility Testing; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK539714/ (accessed on 28 March 2022).
- Leopold, S.J.; van Leth, F.; Tarekegn, H.; Schultsz, C. Antimicrobial drug resistance among clinically relevant bacterial isolates in sub-Saharan Africa: A systematic review. J. Antimicrob. Chemother. 2014, 69, 2337–2353. [Google Scholar] [CrossRef] [Green Version]
- Low, A.; Gavriilidis, G.; Larke, N.; B-Lajoie, M.-R.; Drouin, O.; Stover, J.; Muhe, L.; Easterbrook, P. Incidence of Opportunistic Infections and the Impact of Antiretroviral Therapy Among HIV-Infected Adults in Low- and Middle-Income Countries: A Systematic Review and Meta-analysis. Clin. Infect. Dis. 2016, 62, 1595–1603. [Google Scholar] [CrossRef] [Green Version]
- Kirchhelle, C. Pharming animals: A global history of antibiotics in food production (1935–2017). Palgrave Commun. 2018, 4, 96. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.; Szamosi, Z.; Siménfalvi, Z. Impact of mixing intensity and duration on biogas production in an anaerobic digester: A review. Crit. Rev. Biotechnol. 2020, 40, 508–521. [Google Scholar] [CrossRef] [Green Version]
- Poudel, R.M.; Joshi, D.R.; Dhakal, N.R.; Karki, A.B. Anaerobic digestion of sewage sludge mixture for the reduction of indicator and pathogenic microorganisms. Sci. World 2010, 8, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Thaker, H.C.; Brahmbhatt, M.N.; Nayak, J.B. Study on occurrence and antibiogram pattern of Escherichia coli from raw milk samples in Anand, Gujarat, India. Vet. World 2012, 5, 556–559. [Google Scholar] [CrossRef]
- Omolajaiye, S.A.; Afolabi, K.O.; Iweriebor, B.C. Pathotyping and Antibiotic Resistance Profiling of Escherichia coli Isolates from Children with Acute Diarrhea in Amatole District Municipality of Eastern Cape, South Africa. BioMed Res. Int. 2020, 2020, 4250165. [Google Scholar] [CrossRef] [PubMed]
- Onohuean, H.; Igere, B.E. Occurrence, Antibiotic Susceptibility and Genes Encoding Antibacterial Resistance of Salmonella spp. and Escherichia coli From Milk and Meat Sold in Markets of Bushenyi District, Uganda. Microbiol. Insights 2022, 15, 11786361221088992. [Google Scholar] [CrossRef]
- Morka, K.; Bystroń, J.; Bania, J.; Korzeniowska-Kowal, A.; Korzekwa, K.; Guz-Regner, K.; Bugla-Płoskońska, G. Identification of Yersinia enterocolitica isolates from humans, pigs and wild boars by MALDI TOF MS. BMC Microbiol. 2018, 18, 86. [Google Scholar] [CrossRef]
- Sithole, V.; Amoako, D.G.; Abia, A.L.K.; Perrett, K.; Bester, L.A.; Essack, S.Y. Occurrence, Antimicrobial Resistance, and Molecular Characterization of Campylobacter spp. in Intensive Pig Production in South Africa. Pathogens 2021, 10, 439. [Google Scholar] [CrossRef]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Characterisation and antibiotic resistance of selected bacterial pathogens recovered from dairy cattle manure during anaerobic mono-digestion in a balloon-type digester. Appl.Sci. 2018, 8, 2088. [Google Scholar] [CrossRef] [Green Version]
- Nassar, M.S.M.; Hazzah, W.A.; Bak, W.M.K. Evaluation of antibiotic susceptibility test results: How guilty a laboratory could be? J. Egypt Public Health Assoc. 2019, 94, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.-Q.; Tan, P.-C.; Gao, Y.-M.; Zhang, X.-J.; Xie, Y.; Zheng, D.-N.; Zhou, S.-B.; Li, Q.-F. The effect of glycerol as a cryoprotective agent in the cryopreservation of adipose tissue. Stem Cell Res. Ther. 2022, 13, 152. [Google Scholar] [CrossRef] [PubMed]
- Cheesbrough, M. District Laboratory Practice in Tropical Countries, Part 2: Microbiology; Cambridge University Press: Cambridge, UK, 2000; pp. 234–280. ISBN 9780521676311. [Google Scholar]
- Clinical Laboratory Standard Institute. M100. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; Clinical & Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Resende, J.A.; Silva, V.L.; de Oliveira, T.L.R.; de Oliveira Fortunato, S.; da Costa Carneiro, J.; Otenio, M.H.; Diniz, C.G. Prevalence and persistence of potentially pathogenic and antibiotic resistant bacteria during anaerobic digestion treatment of cattle manure. Bioresour. Technol. 2014, 153, 284–291. [Google Scholar] [CrossRef] [Green Version]
- Davis, R.; Brown, P.D. Multiple antibiotic resistance index, fitness and virulence potential in respiratory Pseudomonas aeruginosa from Jamaica. J. Med. Microbiol. 2016, 65, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, J.S.; Jeggo, M. The One Health Approach—Why Is It So Important? Trop. Med. Infect. Dis. 2019, 4, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawangpa, A.; Lertwatcharasarakul, P.; Boonsoongnern, A.; Ratanavanichrojn, N.; Sanguankiat, A.; Pinniam, N.; Jala, S.; Laopiem, S.; Tulayakul, P. Multidrug resistance problems targeting piglets and environmental health by Escherichia coli in intensive swine farms. Emerg. Contam. 2022, 8, 123–133. [Google Scholar] [CrossRef]
- Manyi-Loh, C.; Lues, R. Reduction in Bacterial Pathogens in a Single-Stage Steel Biodigester Co-Digesting Saw Dust and Pig Manure at Psychrophilic Temperature. Appl. Sci. 2022, 12, 10071. [Google Scholar] [CrossRef]
- Strauch, D. Survival of pathogenic micro-organisms and parasites in excreta, manure and sewage sludge. Rev. Sci. Tech. Off. 1991, 10, 813–846. [Google Scholar] [CrossRef]
- Matjuda, D.S.-M.; Aiyegoro, O.A. Analysis of bacteriological pollution and the detection of antibiotic resistance genes of prevailing bacteria emanating from pig farm seepage. MicrobiologyOpen 2019, 8, e737. [Google Scholar] [CrossRef]
- Peng, Z.; Zou, M.; Li, M.; Liu, D.; Guan, W.; Hao, Q.; Xu, J.; Zhang, S.; Jing, H.; Li, Y.; et al. Prevalence, antimicrobial resistance and phylogenetic characterization of Yersinia enterocolitica in retail poultry meat and swine feces in parts of China. Food Control 2018, 93, 121–128. [Google Scholar] [CrossRef]
- Venglovsky, J.; Sasakova, N.; Gregova, G.; Papajova, I.; Toth, F.; Szaboova, T. Devitalisation of pathogens in stored pig slurry and potential risk related to its application to agricultural soil. Environ. Sci. Pollut. Res. 2018, 25, 21412–21419. [Google Scholar] [CrossRef] [Green Version]
- Milford, A.B.; Le Mouël, C.; Bodirsky, B.L.; Rolinski, S. Drivers of meat consumption. Appetite 2019, 141, 104313. [Google Scholar] [CrossRef]
- South African Veterinary Association (SAVA). Guidelines for the Use of Antimicrobials in the South African Pig Industry. Available online: https://www.medbox.org/document/guidelines-for-the-use-of-antimicrobials-in-the-south-african-pig-industry#GO (accessed on 28 October 2022).
- Musonye, H.A.; Hassanali, A.; Nonoh, J.; Njeu, E.M.; Langata, L.M. Molecular identification and antibiotic susceptibility patterns of bacterial isolates from urine samples of African buffalo, eland and cattle. Int. J. Appl. Biol. 2021, 5, 59–63. [Google Scholar]
- Moyen, R.; Ngoulou, T.B.; Nguimbi, E.; Ahombo, G. Antibiotic Resistance Phenotypes of Enterobacteriaceae Isolated from Household Wastewater in Brazzaville, Republic of Congo. Adv. Microbiol. 2021, 11, 27–36. [Google Scholar] [CrossRef]
- Kilonzo-Nthenge, A.; Rotich, E.; Nahashon, S.N. Evaluation of drug-resistant Enterobacteriaceae in retail poultry and beef. Poult. Sci. 2013, 92, 1098–1107. [Google Scholar] [CrossRef]
- Hennessey, M.; Whatford, L.; Payne-Gifford, S.; Johnson, K.F.; Van Winden, S.; Barling, D.; Häsler, B. Antimicrobial & antiparasitic use and resistance in British sheep and cattle: A systematic review. Prev. Vet. Med. 2020, 185, 105174. [Google Scholar] [CrossRef] [PubMed]
- Abdus Sobur, M.; Sabuj, A.A.M.; Sarker, R.; Tauqifur Rahman, A.M.M.; Lutful Kabir, S.M.; Tanvir Rahman, M. Antibiotic-resistant Escherichia coli and Salmonella spp. associated with dairy cattle and farm environment having public health significance. Vet. World 2019, 12, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Rasschaert, G.; Van Elst, D.; Colson, L.; Herman, L.; de Carvalho Ferreira, H.C.; Dewulf, J.; Decrop, J.; Meirlaen, J.; Heyndrickx, M.; Daeseleire, E. Antibiotic Residues and Antibiotic-Resistant Bacteria in Pig Slurry Used to Fertilize Agricultural Fields. Antibiotics 2020, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, S. Understanding the contribution of environmental factors in the spread of antimicrobial resistance. Environ. Health Prev. Med. 2015, 20, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Lekagul, A.; Tangcharoensathien, V.; Mills, A.; Rushton, J.; Yeung, S. How antibiotics are used in pig farming: A mixed-methods study of pig farmers, feed mills and veterinarians in Thailand. BMJ Glob. Health 2020, 5, e001918. [Google Scholar] [CrossRef] [Green Version]
- Lekagul, A.; Tangcharoensathien, V.; Yeung, S. Patterns of antibiotic use in global pig production: A systematic review. Vet. Anim. Sci. 2019, 7, 100058. [Google Scholar] [CrossRef]
- Singh, A.K.; Das, S.; Kumar, S.; Gajamer, V.R.; Najar, I.N.; Lepcha, Y.D.; Tiwari, H.K.; Singh, S. Distribution of Antibiotic-Resistant Enterobacteriaceae Pathogens in Potable Spring Water of Eastern Indian Himalayas: Emphasis on Virulence Gene and Antibiotic Resistance Genes in Escherichia coli. Front. Microbiol. 2020, 11, 581072. [Google Scholar] [CrossRef]
- Ogbor, O.; Ajayi, A.; Zautner, A.E.; Smith, S.I. Antibiotic susceptibility profiles of Campylobacter coli isolated from poultry farms in Lagos Nigeria—A pilot study. Eur. J. Microbiol. Immunol. 2019, 9, 32–34. [Google Scholar] [CrossRef]
- Sibanda, N.; McKenna, A.; Richmond, A.; Ricke, S.C.; Callaway, T.; Stratakos, A.C.; Gundogdu, O.; Corcionivoschi, N. A Review of the Effect of Management Practices on Campylobacter Prevalence in Poultry Farms. Front. Microbiol. 2018, 9, 2002. [Google Scholar] [CrossRef] [PubMed]
- Zhen, X.; Chen, J.; Sun, X.; Sun, Q.; Guo, S.; Lundborg, C.S. Socioeconomic Factors Contributing to Antibiotic Resistance in China: A Panel Data Analysis. Antibiotics 2021, 10, 994. [Google Scholar] [CrossRef] [PubMed]
- Alividza, V.; Mariano, V.; Ahmad, R.; Charani, E.; Rawson, T.M.; Holmes, A.H.; Castro-Sánchez, E. Investigating the impact of poverty on colonization and infection with drug-resistant organisms in humans: A systematic review. Infect. Dis. Poverty 2018, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Kobeissi, E.; Menassa, M.; Moussally, K.; Repetto, E.; Soboh, I.; Hajjar, M.; Saleh, S.; Abu-Sittah, G. The socioeconomic burden of antibiotic resistance in conflict-affected settings and refugee hosting countries: A systematic scoping review. Confl. Health 2021, 15, 21. [Google Scholar] [CrossRef]
- Tegner, C.; Chandra, N.P.S.; Wijesooriya, W.R.P.L.I.; Perera, B.V.; Hansson, I.; Fahlman, Å. Detection, identification and antimicrobial susceptibility of Campylobacter spp. and Salmonella spp. from Free-ranging nonhuman primates in Sri Lanka. J. Wild Dis. 2019, 55, 879–884. [Google Scholar] [CrossRef]
- Miller, J.H.; Novak, J.T.; Knocke, W.R.; Pruden, A. Survival of Antibiotic Resistant Bacteria and Horizontal Gene Transfer Control Antibiotic Resistance Gene Content in Anaerobic Digesters. Front. Microbiol. 2016, 7, 263. [Google Scholar] [CrossRef] [Green Version]
- Manyi-Loh, C.E.; Mamphweli, S.N.; Meyer, E.L.; Okoh, A.I.; Makaka, G.; Simon, M. Inactivation of Selected Bacterial Pathogens in Dairy Cattle Manure by Mesophilic Anaerobic Digestion (Balloon Type Digester). Int. J. Environ. Res. Public Health 2014, 11, 7184–7194. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Xie, S.H.; Dennehy, C.; Lawlor, P.G.; Hu, Z.H.; Wu, G.X.; Zhan, X.M.; Gardiner, G.E. Inactivation of pathogens in anaerobic digestion systems for converting biowastes to bioenergy: A review. Renew. Sustain. Energy Rev. 2020, 120, 109654. [Google Scholar] [CrossRef]
- Katada, S.; Fukuda, A.; Nakajima, C.; Suzuki, Y.; Azuma, T.; Takei, A.; Takada, H.; Okamoto, E.; Kato, T.; Tamura, Y.; et al. Aerobic Composting and Anaerobic Digestion Decrease the Copy Numbers of Antibiotic-Resistant Genes and the Levels of Lactose-Degrading Enterobacteriaceae in Dairy Farms in Hokkaido, Japan. Front. Microbiol. 2021, 12, 737420. [Google Scholar] [CrossRef]
- Chala, G.; Eguale, T.; Abunna, F.; Asrat, D.; Stringer, A. Identification and characterisation of Campylobacter species in livestock, humans and water in livestock owning households of Peri-Urban Addis Ababa, Ethiopia: A one health approach. Front. Public Health 2021, 9, 750551. [Google Scholar] [CrossRef]


{kind=link}
| Class of Antibiotics | Examples of Antibiotics | Total Number of Bacterial Isolates Investigated in This Study, n = 159 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Yersinia spp. (n = 26) | Escherichia coli (n = 32) | Salmonella spp. (n = 45) | Campylobacter spp. (n = 56) | ||||||||||
| Sensitive | Intermediate | Resistant | Sensitive | Intermediate | Resistant | Sensitive | Intermediate | Resistant | Sensitive | Intermediate | Resistant | ||
| Sulphonamides | Trimethoprim- Sulfamethoxazole (Co-trimoxazole) | 16 (61.54) | 3 (11.54) | 7 (26.92) | 20 (62.50) | 3 (9.38) | 9 (28.13) | 7 (15.56) | 2 (4.44) | 36 (80.00) | 31 (55.36) | 2 (3.57) | 23 (41.07) |
| Sulfamethoxazole | 0 (0.00) | 2 (7.69) | 24 (92.31) | 0 (0.00) | 14 (43.75) | 18 (56.25) | 1 (2.22) | 0 (0.00) | 44 (97.78) | 25 (44.64) | 1 (1.79) | 30 (53.57) | |
| Quinolones | Ciprofloxacin | 21 (80.76) | 3 (11.54) | 2 (7.69) | 13 (40.63) | 14 (43.75) | 5 (15.63) | 1 (2.22) | 16 (35.56) | 28 (62.22) | 17 (30.36) | 9 (16.07) | 30 (53.57) |
| Nalidixic acid | 4 (15.38) | 20 (76.92) | 2 (7.69) | 4 (12.50) | 22 (68.75) | 6 (18.75) | 2 (4.44) | 39 (86.67) | 4 (8.89) | 4 (7.14) | 15 (26.79) | 37 (66.07) | |
| Macrolides | Erythromycin | 0 (0.00) | 0 (0.00) | 26 (100) | 0 (0.00) | 3 (9.38) | 29 (90.63) | 0 (0.00) | 0 (0.00) | 45 (100) | 2 (3.57) | 10 (17.86) | 44 (78.57) |
| Phenicols | Chloramphenicol | 8 (30.77) | 8 (30.77) | 10 (38.46) | 22 (68.75) | 5 (15.63) | 5 (15.63) | 7 (15.56) | 0 (0.00) | 38 (84.44) | 17 (30.36) | 2 (3.57) | 37 (66.07) |
| Tetracyclines | Tetracycline | 9 (34.62) | 5 (19.23) | 12 (46.15) | 4 (12.50) | 2 (6.25) | 26 (81.25) | 0 (0.00) | 0 (0.00) | 45 (100) | 19 (33.93) | 7 (12.5) | 30 (53.57) |
| Penicllins | Amoxicillin-Clavulanate (Augmentin) | 4 (15.38) | 0 (0.00) | 22 (84.62) | 1 (3.13) | 14 (43.77) | 17 (50.13) | 0 (0.00) | 9 (20.00) | 36 (80.00) | 40 (71.43) | 12 (21.43) | 4 (7.14) |
| Ampicillin | 4 (15.38) | 4 (15.38) | 18 (69.23) | 2 (6.25) | 7 (21.88) | 23 (71.88) | 8 (17.78) | 29 (64.44) | 8 (17.78) | 38 (67.86) | 11 (19.64) | 7 (12.5) | |
| Amoxicillin | 1 (3.85) | 2 (7.69) | 23 (88.46) | 32 (100) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 45 (100) | 23 (41.07) | 9 (16.07) | 24 (42.86) | |
| Aminoglycosides | Gentamicin | 20 (76.92) | 5 (19.23) | 1 (3.85) | 12 (37.50) | 14 (43.75) | 6 (18.75) | 31 (68.89) | 12 (26.67) | 2 (4.44) | 22 (39.29) | 2 (3.57) | 32 (57.14) |
| Streptomycin | 25 (96.15) | 1 (3.85) | 0 (0.00) | 21 (65.63) | 10 (31.25) | 1 (3.13) | 42 (93.33) | 3 (6.67) | 0 (0.00) | 26 (46.43) | 3 (5.36) | 27 (48.21) | |
| Nitrofurans | Nitrofurantoin | 12 (46.15) | 12 (46.15) | 2 (7.69) | 5 (15.63) | 10 (31.25) | 17 (53.13) | 2 (4.44) | 11 (24.44) | 32 (71.11) | 42 (75.00) | 2 (3.57) | 12 (21.43) |
| Cephalosporins | Cefotaxime | 8 (30.77) | 12 (46.15) | 6 (23.08) | 0 (0.00) | 9 (28.13) | 23 (71.88) | 1 (2.22) | 3 (6.67) | 41 (91.11) | 0 (0.00) | 1 (1.79) | 55 (98.21) |
| Bacterial Isolates | Range of MAR Indices | Number of Bacterial Isolates with MAR > 0.2 (n = 145) | Percentage (%) |
|---|---|---|---|
| Yersinia spp. | 0.3–0.7 | 26 | 17.93 |
| E. coli | 0.1–0.8 | 29 | 20.00 |
| Salmonella spp. | 0.4–0.9 | 45 | 31.03 |
| Campylobacter spp. | 0.1–0.9 | 45 | 31.03 |
| Total | 145 | 100 |
| Yersinia spp. (n = 26) | ||
|---|---|---|
| No. of Antibiotics | MAR Phenotypes (n= 19) | Frequency of Isolates in Percentages (%) |
| 5 | E, AUG, AP, AMOX, SMX | 4 (15.38%) |
| 5 | E, AUG, AP, AMOX, NA | 1 (3.85%) |
| 6 | E, AUG, AP, AMOX, SMX, NI | 2 (7.69%) |
| 6 | E, SMX, TET, TS, C, NI | 2 (7.69%) |
| 6 | E, AUG, AP, AMOX, SMX, NI | 1 (3.85%) |
| 6 | E, AUG, AP, AMOX, SMX, NA | 1 (3.85%) |
| 6 | E, AP, AMOX, SMX, CTX, CIP | 1 (3.85%) |
| 7 | E, AUG, AP, AMOX, SMX, CTX, C | 1 (3.85%) |
| 7 | E, AUG, AP, AMOX, SMX, NI, CIP | 1 (3.85%) |
| 7 | E, SMX, TET, TS, C, NI, GM | 1 (3.85%) |
| 7 | E, AUG, AP, AMOX, SMX, CTX, NI | 1 (3.85%) |
| 7 | E, AUG, AP, AMOX, SMX, TET, NI | 1 (3.85%) |
| 7 | E, AUG, AMOX, SMX, TET, TS, C | 1 (3.85%) |
| 8 | E, AUG, AP, AMOX, SMX, TET, NI, C, | 2 (7.69%) |
| 8 | E, AUG, AP, AMOX, SMX, TET, TS, C | 1 (3.85%) |
| 8 | E, AUG, AMOX, SMX, CTX, TET, TS, C | 1 (3.85%) |
| 10 | E, AUG, AP, AMOX, SMX, CTX, TET, TS, C, NI | 1 (3.85%) |
| Total of MDR isolates | 26 | |
| Percentage MDR | 100% | |
| Escherichia coli(spp.) (n = 32) | ||
| No. of Antibiotics | MAR Phenotypes (n = 29) | Frequency of Isolates in Percentages (%) |
| 3 | AMOX, TET, AP | 1 (3.13%) |
| 3 | AMOX, TET, CTX | 1 (3.13%) |
| 4 | E, AMOX, SMX, TET | 1 (3.13%) |
| 5 | E, AMOX, CTX, TET, NI | 1 (3.13%) |
| 5 | E, AUG, AP, AMOX, TET | 1 (3.13%) |
| 6 | E, AP, AMOX, SMX, CTX, NA | 1 (3.13%) |
| 6 | E, AUG, AP, AMOX, CTX, TET | 1 (3.13%) |
| 6 | E, AMOX, SMX, CIP, NA, GM | 1 (3.13%) |
| 6 | E, AP, AMOX, CTX, TET, NI | 2 (6.25%) |
| 6 | E, AMOX, SMX, CTX, TS, C | 1 (3.13%) |
| 6 | E, AP, AMOX, SMX, TET, NI | 1 (3.13%) |
| 7 | E, AUG, AP, AMOX, SMX, CTX, TET | 1 (3.13%) |
| 7 | E, AUG, AP, AMOX, CTX, TET, NI | 1 (3.13%) |
| 7 | E, AP, AMOX, SMX, CIP, GM, NI | 1 (3.13%) |
| 8 | E, AUG, AP, AMOX, SMX, CTX, TET, TS | 1 (3.13%) |
| 8 | E, AUG, AP, AMOX, CTX, TET, NA, NI | 2 (6.25%) |
| 8 | E, AUG, AP, AMOX, SMX, CTX, TET, CIP | 1 (3.13%) |
| 8 | E, AUG, AP, AMOX, SMX, CTX, TET, NI | 1 (3.13%) |
| 8 | E, AUG, AP, AMOX, CTX, TET, CIP, NI | 1 (3.13%) |
| 8 | E, AUG, AP, AMOX, SMX, TET, NI, TS | 1 (3.13%) |
| 9 | E, AMOX, SMX, CTX, TET, NI, TS, C, GM | 1 (3.13%) |
| 9 | E, AUG, AP, AMOX, SMX, CTX, NI, C, TS | 1 (3.13%) |
| 9 | E, AUG, AP, AMOX, SMX, CTX, TET, NA, NI | 1 (3.13%) |
| 10 | E, AUG, AP, AMOX, SMX, CTX, TET, TS, S, GM | 1 (3.13%) |
| 10 | E, AMOX, SMX, CTX, TET, NA, NI, GM, C, TS | 1 (3.13%) |
| 10 | E, AUG, AP, AMOX, SMX, CTX, TET, TS, NI, CIP | 1 (3.13%) |
| 11 | E, AUG, AP, AMOX, SMX, CTX, TET, TS, NI, C, GM | 1 (3.13%) |
| Total of MDR isolates | 29 | |
| Percentage MDR | 90.63% | |
| Salmonella spp. (n = 45) | ||
| No. of Antibiotics | MAR Phenotypes (n = 22) | Frequency of Isolates in Percentages (%) |
| 5 | E, AMOX, TET, NA, NI | 1 (2.22%) |
| 6 | E, AUG, AMOX, SMX, TET, NI | 2 (4.44%) |
| 7 | E, AUG, AMOX, SMX, CTX, TET, NI | 2 (4.44%) |
| 7 | E, AMOX, SMX, CTX, TET, TS, C | 1 (2.22%) |
| 8 | E, AMOX, SMX, CTX, TET, TS, C, CIP | 3 (6.67%) |
| 8 | E, AUG, AMOX, SMX, CTX, TET, TS, C | 1 (2.22%) |
| 9 | E, AUG, AMOX, SMX, CTX, TET, TS, C, NI | 4 (8.89%) |
| 9 | E, AUG, AMOX, SMX, CTX, TET, TS, C, CIP | 2 (4.44%) |
| 9 | E, AMOX, SMX, CTX, TET, TS, C, CIP, NI | 2 (4.44%) |
| 9 | E, AP, AMOX, SMX, CTX, TET, TS, C, CIP | 1 (2.22%) |
| 9 | E, AUG, AP, AMOX, SMX, CTX, TET, TS, C | 1 (2.22%) |
| 9 | E, AUG, AMOX, SMX, CTX, TET, TS, C, NI | 2 (4.44%) |
| 9 | E, AUG, AP, AMOX, SMX, CTX, TET, C, NI | 1 (2.22%) |
| 9 | E, AMOX, SMX, CTX, TET, TS, C, CIP, NA | 1 (2.22%) |
| 9 | E, AUG, AMOX, SMX, CTX, TET, CIP, GM, NI | 1 (2.22%) |
| 10 | E, AUG, AMOX, SMX, CTX, TET, TS, C, CIP, NI | 13 (28.89%) |
| 10 | E, AUG, AP, AMOX, SMX, CTX, TET, TS, C, CIP | 2 (4.44%) |
| 10 | E, AUG, AP, AMOX, SMX, CTX, TET, CIP, C, NI | 1 (2.22%) |
| 11 | E, AUG, AMOX, SMX, CTX, TET, TS, C, CIP, NA, NI | 1 (2.22%) |
| 11 | E, AUG, AP, AMOX, SMX, CTX, TET, TS, C, GM, NI | 1 (2.22%) |
| 12 | E, AUG, AP, AMOX, SMX, CTX, TET, TS, C, CIP, NA, NI | 1 (2.22%) |
| Total MDR isolates | 45 | |
| Percentage MDR | 100% | |
| Campylobacter spp. (n = 56) | ||
| No. of Antibiotics | MAR Phenotypes (n = 27) | Frequency of Isolates in Percentages (%) |
| 3 | E, CTX, AMOX | 3 (5.36%) |
| 3 | E, CTX, NA, | 1 (1.79%) |
| 3 | AMOX, CTX, AUG | 1 (1.79%) |
| 3 | AMOX, CTX, NA | 2 (3.57%) |
| 4 | AMOX, CTX, TET, AP | 1 (1.79%) |
| 5 | CTX, C, NI, E, AMOX | 4 (7.14%) |
| 5 | CTX, C, NI, E, SMX, | 1 (1.79%) |
| 5 | CTX, C, NI, AP, AMOX | 2 (3.57%) |
| 5 | CTX, CIP, NA, GM, SMX | 1 (1.79%) |
| 6 | CTX, CIP, NA, GM, E, C | 2 (3.57%) |
| 6 | CTX, C, NI, AP, AMOX, E | 2 (3.57%) |
| 6 | E, CTX, CIP, NA, GM, S | 1 (1.79%) |
| 6 | E, CTX, CIP, NA, GM, SMX | 1 (1.79%) |
| 6 | CTX, C, NI, E, AMOX, NA | 1 (1.79%) |
| 6 | CTX, S, NI, AMOX, AUG, TET | 1 (1.79%) |
| 8 | E, SMX, CTX, TET, C, NA, GM, TS | 1 (1.79%) |
| 8 | E, SMX, CTX, TET, C, NA, GM, S | 1 (1.79%) |
| 8 | E, SMX, CTX, TET, AMOX, NA, GM, S | 1 (1.79%) |
| 8 | E, SMX, CTX, TET, CIP, NA, GM, TS | 1 (1.79%) |
| 9 | E, SMX, TET, CIP, NA, GM, TS, S, C | 1 (1.79%) |
| 9 | E, SMX, CTX, TET, CIP, NA, GM, S, C | 1 (1.79%) |
| 10 | E, SMX, CTX, TET, CIP, NA, GM, S, C, TS | 15 (26.79%) |
| 11 | E, SMX, CTX, TET, CIP, NA, GM, S, C, TS, AMOX | 3 (5.36%) |
| 11 | E, SMX, CTX, TET, CIP, NA, GM, S, C, TS, NI | 1 (1.79%) |
| 11 | E, SMX, CTX, TET, CIP, NA, GM, S, C, TS, AUG | 1 (1.79%) |
| 13 | E, SMX, CTX, TET, CIP, NA, GM, S, C, TS, AUG, AP, AMOX | 1 (1.79%) |
| Total MDR isolates | 45 | |
| Percentage MDR | 80.36% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manyi-Loh, C.E.; Okoh, A.I.; Lues, R. Prevalence of Multidrug-Resistant Bacteria (Enteropathogens) Recovered from a Blend of Pig Manure and Pinewood Saw Dust during Anaerobic Co-Digestion in a Steel Biodigester. Int. J. Environ. Res. Public Health 2023, 20, 984. https://doi.org/10.3390/ijerph20020984
Manyi-Loh CE, Okoh AI, Lues R. Prevalence of Multidrug-Resistant Bacteria (Enteropathogens) Recovered from a Blend of Pig Manure and Pinewood Saw Dust during Anaerobic Co-Digestion in a Steel Biodigester. International Journal of Environmental Research and Public Health. 2023; 20(2):984. https://doi.org/10.3390/ijerph20020984
Chicago/Turabian StyleManyi-Loh, Christy Echakachi, Anthony Ifeanyin Okoh, and Ryk Lues. 2023. "Prevalence of Multidrug-Resistant Bacteria (Enteropathogens) Recovered from a Blend of Pig Manure and Pinewood Saw Dust during Anaerobic Co-Digestion in a Steel Biodigester" International Journal of Environmental Research and Public Health 20, no. 2: 984. https://doi.org/10.3390/ijerph20020984
APA StyleManyi-Loh, C. E., Okoh, A. I., & Lues, R. (2023). Prevalence of Multidrug-Resistant Bacteria (Enteropathogens) Recovered from a Blend of Pig Manure and Pinewood Saw Dust during Anaerobic Co-Digestion in a Steel Biodigester. International Journal of Environmental Research and Public Health, 20(2), 984. https://doi.org/10.3390/ijerph20020984