
Relationship Functioning and Gut Microbiota Composition among Older Adult Couples
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Self-Report Measures
2.2. Fecal Sample Collection, Sequencing, and Microbiota Analysis
2.3. Analytic Plan
3. Results
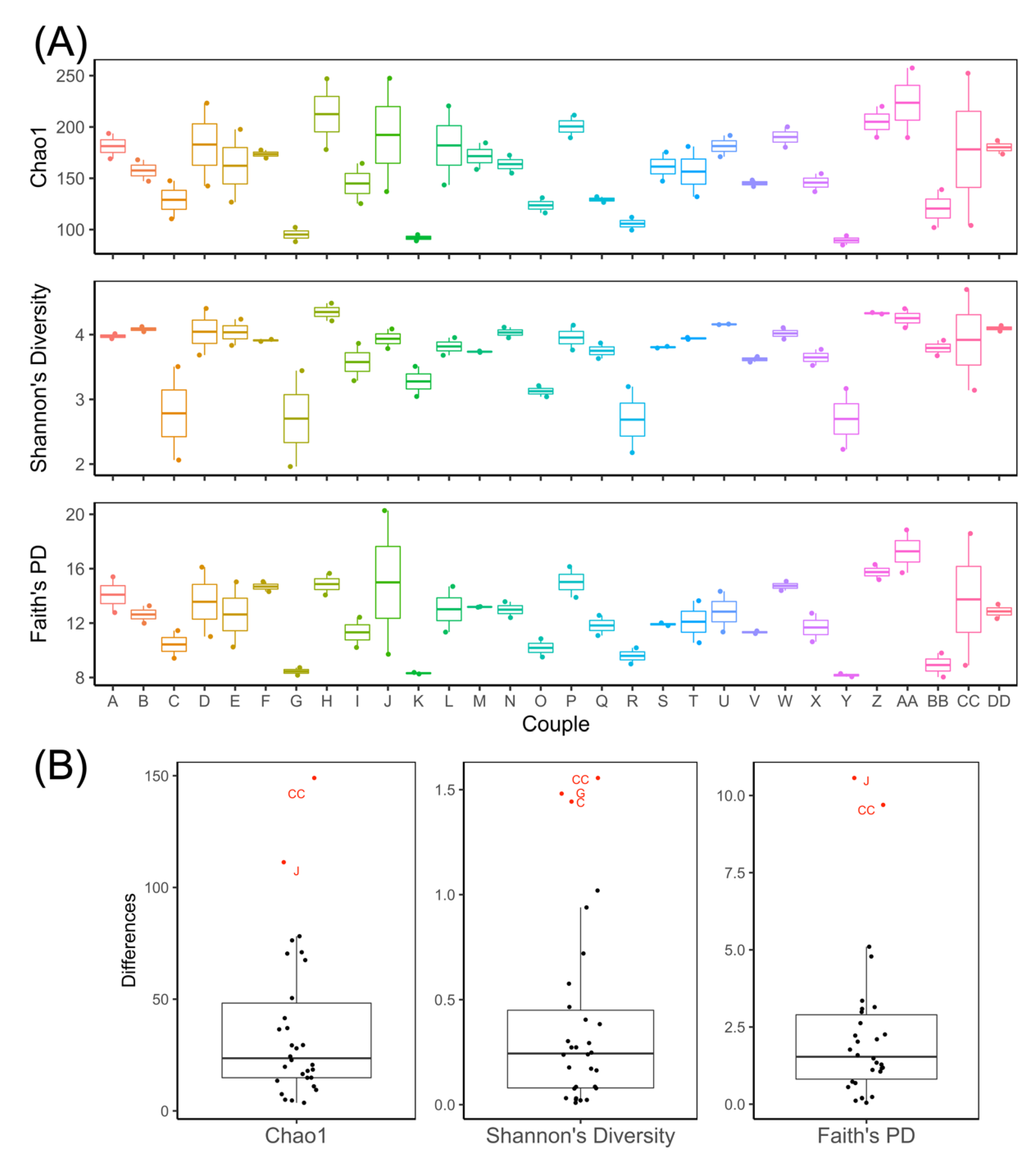
3.1. Remote Recruitment and Assessment of Older Adult Couples, including Collection of Fecal Samples, Is Feasible
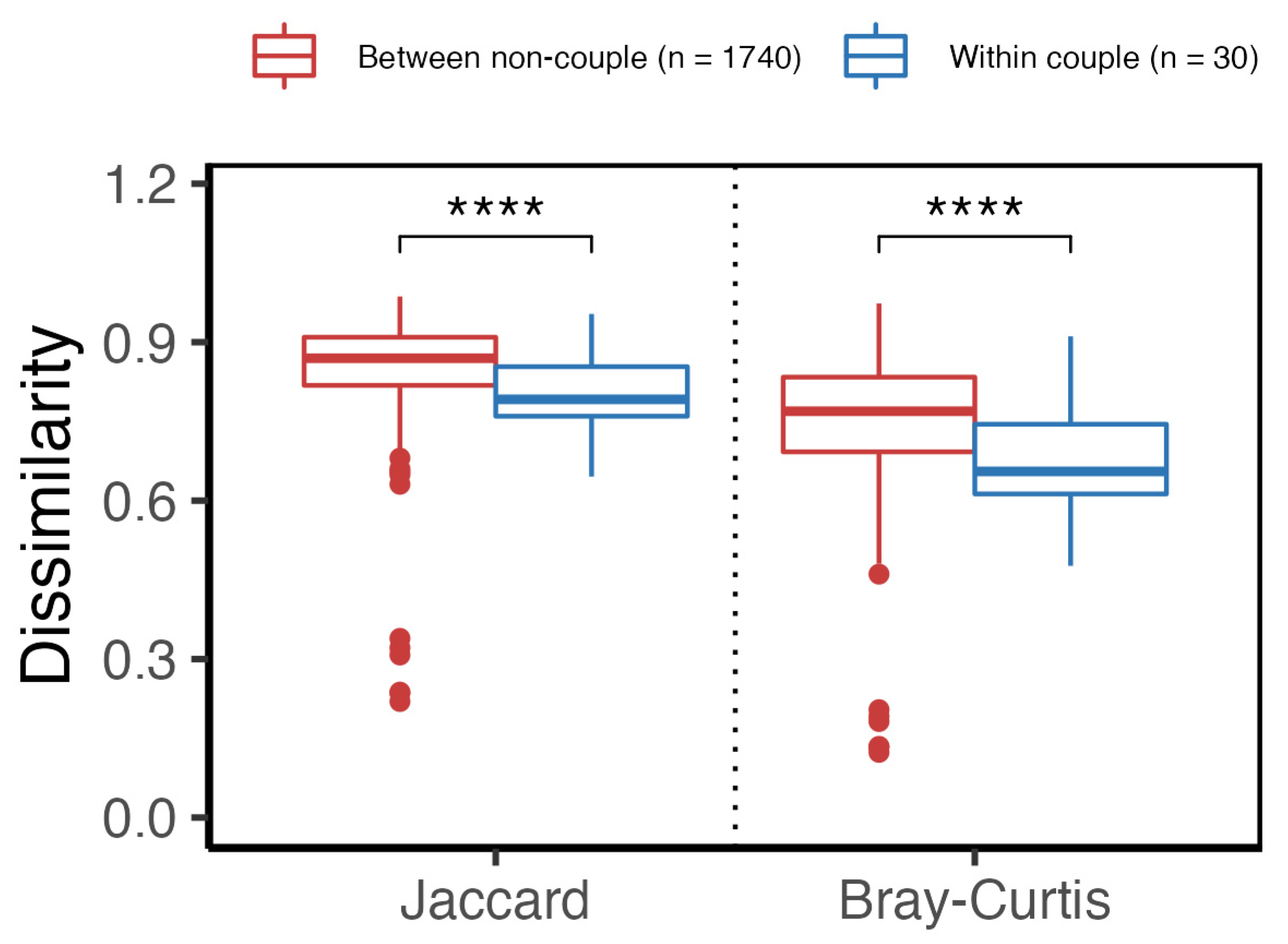
3.2. Gut Microbiota of Couples Share More Similarity Than Those of Non-Couples
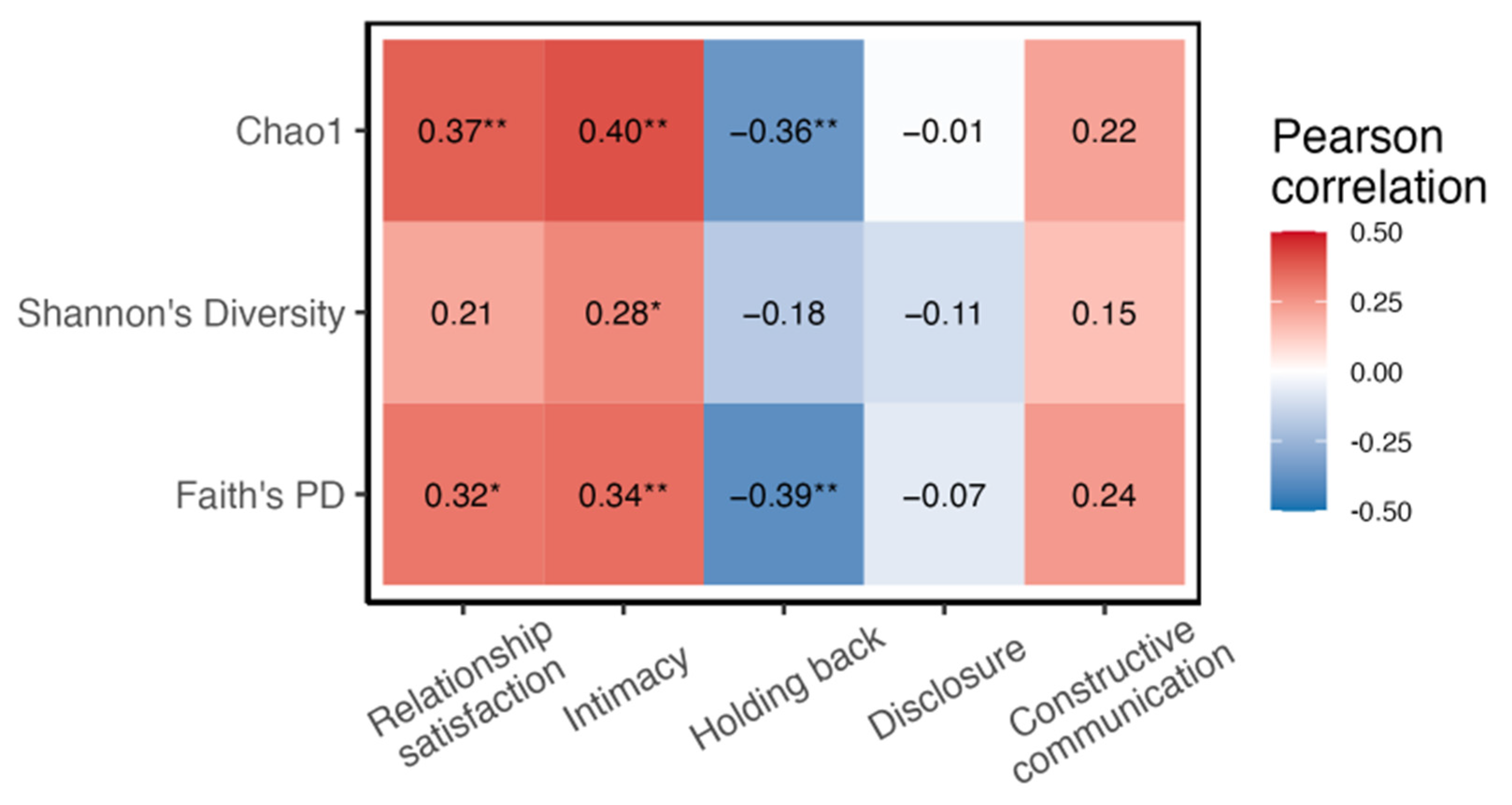
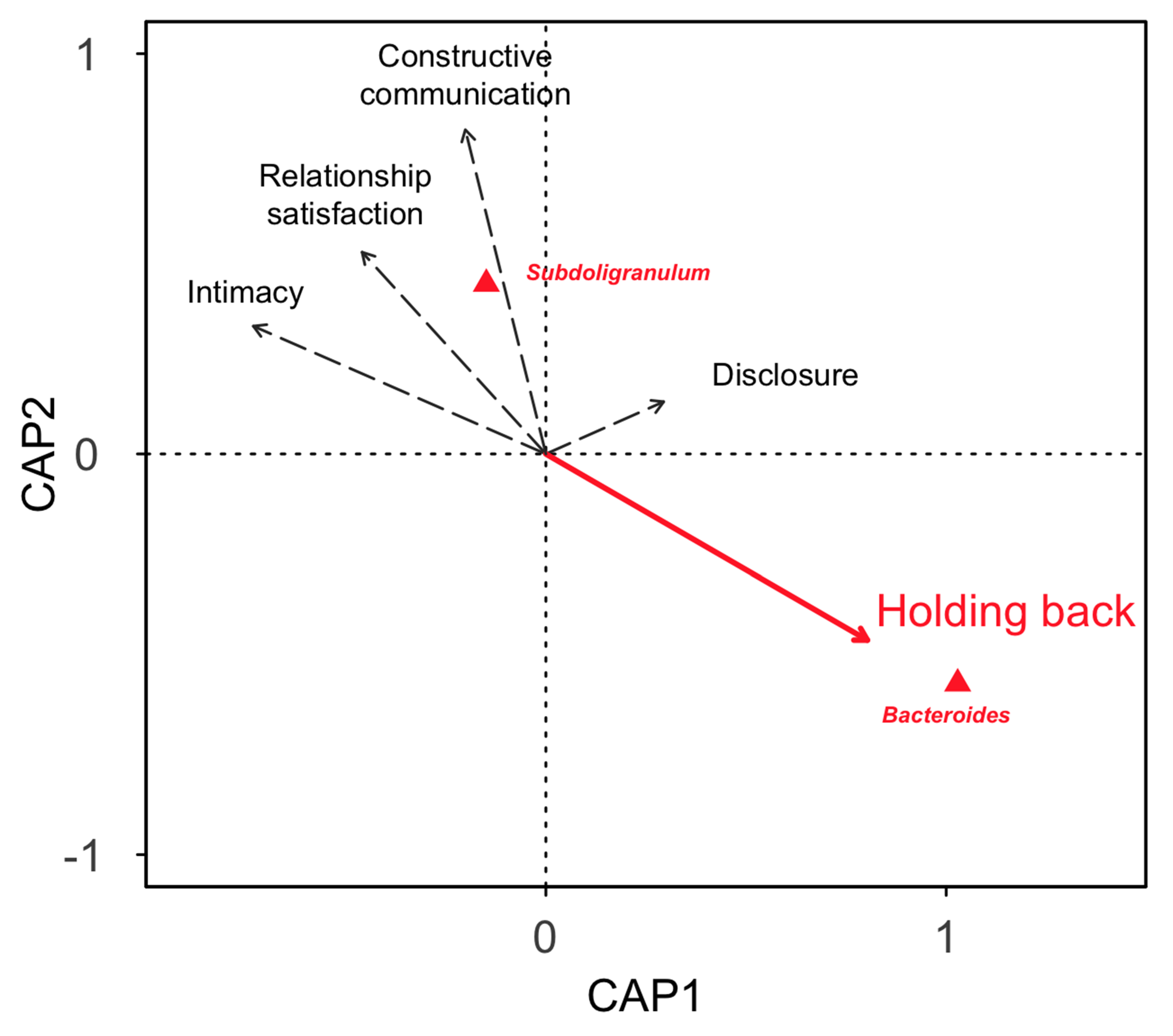
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holt-Lunstad, J. Loneliness and social isolation as risk factors: The power of social connection in prevention. Am. J. Lifestyle Med. 2021, 15, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Doyle, W.; Skoner, D.; Rabin, B.; Gwaitney, J. Social ties and susceptibility to the common cold. J. Am. Med. Assoc. 1997, 277, 1940–1944. [Google Scholar] [CrossRef] [Green Version]
- Holt-Lunstad, J.; Smith, T.; Layton, J. Social relationships and mortality risk: A meta-analytic review. PLoS Med. 2010, 7, e1000316. [Google Scholar] [CrossRef]
- House, J.; Landis, K.; Umberson, D. Social relationships and health. Science 1988, 241, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Robles, T.; Slatcher, R.; Trombello, J.; McGinn, M. Marital quality and health: A meta-analytic review. Psychol. Bull. 2014, 140, 140–187. [Google Scholar] [CrossRef] [Green Version]
- Aizer, A.; Chen, M.-H.; McCarthy, E.; Mendu, M.; Koo, S.; Wilhite, T.; Graham, P.; Choueiri, T.; Hoffman, K.; Martin, N.; et al. Marital status and survival in patients with cancer. J. Clin. Oncol. 2013, 31, 3869–3876. [Google Scholar] [CrossRef]
- Kiecolt-Glaser, J.; Newton, T. Marriage and health: His and hers. Psychol. Bull. 2001, 127, 472–503. [Google Scholar] [CrossRef]
- Cornelius, T.; Birk, J.; Edmondson, D.; Schwartz, J. Romantic relationship satisfaction and ambulatory blood pressure during social interactions: Specificity or spillover effects? Ann. Behav. Med. 2019, 53, 223–231. [Google Scholar] [CrossRef]
- Whisman, M.; Gilmour, A.; Salinger, J. Marital satisfaction and mortality in the United States adult population. Health Psychol. 2018, 37, 1041–1044. [Google Scholar] [CrossRef]
- Di Castelnuovo, A.; Quacquaruccio, G.; Donati, M.; de Gaetano, G.; Iacoviello, L. Spousal concordance for major coronary risk factors: A systematic review and meta-analysis. Am. J. Epidemiol. 2008, 169, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Macken, L.; Yates, B.; Blancher, S. Concordance of risk factors in female spouses of male patients with coronary heart disease. J. Cardiopulm. Rehabil. 2000, 20, 361–368. [Google Scholar] [CrossRef]
- Bookwala, J.; Schulz, R. Spousal similarity in subjective well-being: The Cardiovascular Health Study. Psychol. Aging 1996, 11, 582–590. [Google Scholar] [CrossRef]
- Meyler, D.; Stimpson, J.; Peek, M. Health concordance within couples: A systematic review. Soc. Sci. Med. 2007, 64, 2297–2310. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Bolnick, D.; Kirkpatrick, M. Assortative mating in animals. Am. Nat. 2013, 181, E125–E138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichter, D.; Qian, Z. The study of assortative mating: Theory, data, and analysis. In Analytical Family Demography; The Springer Series on Demographic Methods and Population, Analysis; Schoen, R., Ed.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Umberson, D. Gender, marital status and the social control of health behavior. Soc. Sci. Med. 1992, 34, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickard, J.; Zeng, M.; Caruso, R.; Nunez, G. Gut microbiota: Role in pathogen colonization, immune responses and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef]
- Durack, J.; Lynch, S. The gut microbiome: Relationships with disease and opportunities for therapy. J. Exp. Med. 2019, 216, 20–40. [Google Scholar] [CrossRef] [Green Version]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Finnicum, C.T.; Beck, J.J.; Dolan, C.V.; Davis, C.; Willemsen, G.; Ehli, E.A.; Boomsma, D.I.; Davies, G.E.; de Geus, E.J.C. Cohabitation is associated with a greater resemblance in gut microbiota which can impact cardiometabolic and inflammatory risk. BMC Microbiol. 2019, 19, 230. [Google Scholar] [CrossRef]
- Amato, K.R.; Van Belle, S.; Di Fiore, A.; Estrada, A.; Stumpf, R.; White, B.; Nelson, K.E.; Knight, R.; Leigh, S.R. Patterns in gut microbiota similarity associated with degree of sociality among sex classes of a neotropical primate. Microb. Ecol. 2017, 74, 250–258. [Google Scholar] [CrossRef]
- Grieneisen, L.E.; Livermore, J.; Alberts, S.; Tung, J.; Archie, E.A. Group living and male dispersal predict the core gut microbiome in wild baboons. Integr. Comp. Biol. 2017, 57, 770–785. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.; Godneva, A.; Kalka, I.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Tung, J.; Barreiro, L.B.; Burns, M.B.; Grenier, J.C.; Lynch, J.; Grieneisen, L.E.; Altmann, J.; Alberts, S.C.; Blekhman, R.; Archie, E.A. Social networks predict gut microbiome composition in wild baboons. Elife 2015, 4, e05224. [Google Scholar] [CrossRef]
- Song, S.J.; Lauber, C.; Costello, E.K.; Lozupone, C.A.; Humphrey, G.; Berg-Lyons, D.; Caporaso, J.G.; Knights, D.; Clemente, J.C.; Nakielny, S.; et al. Cohabiting family members share microbiota with one another and with their dogs. Elife 2013, 2, e00458. [Google Scholar] [CrossRef]
- Valles-Colomer, M.; Blanco-Miguez, A.; Manghi, P.; Asnicar, F.; Dubois, L.; Golzato, D.; Armanini, F.; Cumbo, F.; Huang, K.; Manara, S.; et al. The person-to-person transmission landscape of the gut and oral microbiomes. Nature, 2023; Epub ahead of print. [Google Scholar]
- Lax, S.; Smith, D.; Hampton-Marcell, J.; Owens, S.; Handley, K.; Scott, N.; Gibbons, S.; Larsen, P.; Shogan, B.; Weiss, S.; et al. Longitudinal analysis of microbial interaction between humans and the indoor environment. Science 2014, 345, 1048–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dill-McFarland, K.; Tang, Z.-Z.; Kernis, J.; Kerby, R.; Chen, G.; Palloni, A.; Sorenson, T.; Rey, F.; Herd, P. Close social relationships correlate with human gut microbiota composition. Sci. Rep. 2019, 9, 703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baucom, D.; Porter, L.; Kirby, J.; Hudepohl, J. Couple-based interventions for medical problems. Behav. Ther. 2012, 43, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Bookwala, J.; Gaugler, T. Relationship quality and 5-year mortality risk. Health Psychol. 2020, 39, 633–641. [Google Scholar] [CrossRef]
- Kiecolt-Glaser, J.; Wilson, S.; Madison, A. Marriage and gut (microbiome) feelings: Tracing novel dyadic pathways to accelerated aging. Psychosom. Med. 2019, 81, 704–710. [Google Scholar] [CrossRef] [Green Version]
- Langer, S.; Romano, J.; Todd, M.; Strauman, T.; Keefe, F.; Syrjala, K.; Bricker, J.; Ghosh, N.; Burns, J.; Bolger, N.; et al. Links between communication and relationship satisfaction among patients with cancer and their spouses: Results of a fourteen-day smartphone-based ecological momentary assessment study. Front. Psychol. 2018, 9, 1843. [Google Scholar] [CrossRef] [PubMed]
- Helgeson, V. The effects of masculinity and social support on recovery from myocardial infarction. Psychosom. Med. 1991, 53, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Courtin, E.; Knapp, M. Social isolation, loneliness and health in old age: A scoping review. Health Soc. Care Community 2017, 25, 799–812. [Google Scholar] [CrossRef]
- Spanier, G. Measuring dyadic adjustment: New scales for assessing the quality of marriage and similar dyads. J. Marriage Fam. 1976, 38, 15–28. [Google Scholar] [CrossRef]
- Miller, R.S.; Lefcourt, H.M. Miller Social Intimacy Scale. In Measures for Clinical Practice: A Sourcebook, 3rd ed.; Corcoran, K., Fischer, J., Eds.; Free Press: New York, NY, USA, 2000. [Google Scholar]
- Pistrang, N.; Barker, C. Disclosure of concerns in breast cancer. Psycho-Oncology 1992, 1, 183–192. [Google Scholar] [CrossRef]
- Christensen, A. Detection of conflict patterns in couples. In Understanding Major Mental Disorder: The Contribution of Family Interaction Research; Hahlweg, K., Goldstein, M., Eds.; Family Process Press: New York, NY, USA, 1987; pp. 250–265. [Google Scholar]
- Division of Cancer Control and Population Sciences, National Cancer Institute, Dietary Screener Questionnaire (DSQ) in the NHANES 2009–2010. Available online: https://epi.grants.cancer.gov/nhanes/dietscreen/questionnaires.html (accessed on 24 July 2021).
- Chaplin, A.; Parra, P.; Laraichi, S.; Serra, F.; Palou, A. Calcium supplementation modulates gut microbiota in a prebiotic manner in dietary obese mice. Mol. Nutr. Food Res. 2016, 60, 468–480. [Google Scholar] [CrossRef]
- Di Rienzi, S.; Britton, R. Adaptation of the gut microbiota to modern dietary sugars and sweeteners. Adv. Nutr. 2020, 11, 616–629. [Google Scholar] [CrossRef] [Green Version]
- Makki, K.; Deehan, E.; Walter, J.; Backhed, F. The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Oliver, A.; Chase, A.; Weihe, C.; Orchanian, S.; Riedel, S.; Hendrickson, C.; Lay, M.; Sewall, J.; Martiny, J.; Whiteson, K. High-fiber, whole-food dietary intervention alters the human gut microbiome but not fecal short-chain fatty acids. mSystems 2021, 6, e00115-21. [Google Scholar] [CrossRef]
- Ramne, S.; Brunkwall, L.; Ericson, U.; Gray, N.; Kuhnle, G.; Nilsson, P.; Orho-Melander, M.; Sonestedt, E. Gut microbiota composition in relation to intake of added sugar, sugar-sweetened beverages and artificially sweetened beverages in the Malmo Offspring Study. Eur. J. Nutr. 2021, 60, 2087–2097. [Google Scholar] [CrossRef]
- Charlson, M.; Pompei, P.; Ales, K.; MacKenzie, C. A new method of classifying prognostic comorbidity in longitudinal studies: Development and validation. J. Chronic Dis. 1987, 40, 373–383. [Google Scholar] [CrossRef]
- Caporaso, J.; Lauber, C.; Walters, W.; Berg-Lyons, D.; Lozupone, C.; Turnbaugh, P.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilhan, Z.; DiBaise, J.; Dautel, S.; Isern, N.; Kim, Y.; Hoyt, D.; Schepmoes, A.; Brewer, H.; Weitz, K.; Metz, T.; et al. Temporospatial shifts in the human gut microbiome and metabolome after gastric bypass surgery. NPJ Biofilms Microbiomes 2020, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Bisanz, J. qiime2R: Importing QIIME2 Artifacts and Associated Data into R Sessions; R Package Version 0.99.6. 2018. Available online: https://github.com/jbisanz/qiime2R (accessed on 30 January 2023).
- McMurdie, P.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A. Ecological Diversity and Its Measurement; Springer: Dordrecht, The Netherlands, 1988. [Google Scholar]
- Lahti, L.; Shetty, S. microbiome; R Package Version 1.18.0; 2012-2019. Available online: http://microbiome.github.io (accessed on 30 January 2023).
- Kembel, S. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [Green Version]
- Smith, D. rbiom: Read/Write, Transform, and Summarize ‘BIOM’ Data; R Package Version 1.0.3. 2021. Available online: https://CRAN.R-project.org/package=rbiom (accessed on 30 January 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Ara, T. brunnermunzel: (Permuted) Brunner-Munzel Test; R Package Version 2.0. 2022. Available online: https://CRAN.R-project.org/package=brunnermunzel (accessed on 30 January 2023).
- Neubert, K.; Brunner, E. A studentized permutation test for non-parametric Behrens-Fisher problem. Comput. Stat. Data Anal. 2007, 51, 5192–5204. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. vegan: Community Ecology Package; R Package Version 2.6-2. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 30 January 2023).
- Revelle, W. psych: Procedures for Personality and Psychological Research; R Package Version 2.2.5. 2022. Available online: https://CRAN.R-project.org/package=psych (accessed on 30 January 2023).
- Chao, A. Non-parametric estimation of the classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Shannon, C.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Champaign, IL, USA, 1949. [Google Scholar]
- Faith, D. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Jaccard, P. Nouvelles recherches sur la distribution florale. Bull. De La Soc. Vaud. Des Sci. Nat. 1908, 44, 223–270. [Google Scholar]
- Sorensen, T.; I kommission hos, E. A Method of Establishing Groups of Equal Amplitude in Plant Sociology Based on Similarity of Species Content and Its Application to Analyses of the Vegetation on Danish Commons; I kommission hos E. Munksgaard: København, Denmark, 1948. [Google Scholar]
- Blanchet, F.; Legendre, P.; Borcard, D. Forward selection of explanatory variables. Ecology 2008, 89, 2623–2632. [Google Scholar] [CrossRef] [PubMed]
- Raulo, A.; Allen, B.E.; Troitsky, T.; Husby, A.; Firth, J.A.; Coulson, T.; Knowles, S.C.L. Social networks strongly predict the gut microbiota of wild mice. ISME J. 2021, 15, 2601–2613. [Google Scholar] [CrossRef] [PubMed]
- Mosca, A.; Leclerc, M.; Hugot, J.P. Gut Microbiota Diversity and Human Diseases: Should We Reintroduce Key Predators in Our Ecosystem? Front. Microbiol. 2016, 7, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmu, J.; Lahti, L.; Niiranen, T. Targeting gut microbiota to treat hypertension: A systematic review. Int. J. Environ. Res. Public Health 2021, 18, 1248. [Google Scholar] [CrossRef]
- Zafar, H.; Saier, M. Gut Bacteroides species in health and disease. Gut Microbes 2021, 13, e1848158. [Google Scholar] [CrossRef]
- Holmstrom, K.; Collins, M.; Moller, T.; Falsen, E.; Lawson, P. Subdoligranulum variabile gen. nov., sp. nov. from human feces. Anaerobe 2004, 10, 197–203. [Google Scholar] [CrossRef]
- Van Hul, M.; Le Roy, T.; Prifti, E.; Dao, M.; Paquot, A.; Zucker, J.; Delzenne, N.; Muccioli, G.; Clement, K.; Cani, P. From correlation to causality: The case of Subdoligranulum. Gut Microbes 2020, 12, e1849998. [Google Scholar] [CrossRef]
- Adams, R.; Winger, J.; Mosher, C. A meta-analysis of the relationship between social constraints and distress in cancer patients. J. Behav. Med. 2015, 38, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Hasson-Ohayon, I.; Goldzweig, G.; Braun, M.; Hagedoorn, M. Beyond “being open about it”: A systematic review on cancer related communication within couples. Clin. Psychol. Rev. 2022, 96, 102176. [Google Scholar] [CrossRef]
- Traa, M.; De Vries, J.; Bodenmann, G.; Den Oudsten, B. Dyadic coping and relationship functioning in couples coping with cancer: A systematic review. Br. J. Health Psychol. 2015, 20, 85–114. [Google Scholar] [CrossRef] [PubMed]
- Gross, J. Antecedent- and response-focused emotion regulation: Divergent consequences for experience, expression, and physiology. J. Personal. Soc. Psychol. 1998, 74, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Butler, E.; Egloff, B.; Wilhelm, F.; Smith, N.; Erickson, E.; Gross, J. The social consequences of expressive suppression. Emotion 2003, 3, 48–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina-Torres, G.; Rodriguez-Arrastia, M.; Roman, P.; Sanchez-Labraca, N.; Cardona, D. Stress and the gut microbiota-brain axis. Behav. Pharmacol. 2019, 30, 187–200. [Google Scholar] [CrossRef]
- Langer, S.; Romano, J.; Keefe, F.; Baucom, D.; Strauman, T.; Syrjala, K.; Bolger, N.; Burns, J.; Bricker, J.; Todd, M.; et al. Couple communication in cancer: Protocol for a multi-method examination. Front. Psychol. 2022, 12, 769407. [Google Scholar] [CrossRef]
- McBurney, M.; Davis, C.; Fraser, C.; Schneeman, B.; Huttenhower, C.; Verbeke, K.; Walter, J.; Latulippe, M. Establishing what constitutes a healthy human gut microbiome: State of the science, regulatory considerations, and future directions. J. Nutr. 2019, 149, 1882–1895. [Google Scholar] [CrossRef] [Green Version]
- Ragonnaud, E.; Biragyn, A. Gut microbiota as the key controllers of “healthy” aging of elderly people. Immun. Ageing 2021, 18, 1–11. [Google Scholar] [CrossRef]
- Holt-Lunstad, J. Social connection as a public health issue: The evidence and a systemic framework for prioritizing the “social” in social determinants of health. Annu. Rev. Public Health 2022, 43, 193–213. [Google Scholar] [CrossRef]
- Somes, J. The loneliness of aging. J. Emerg. Nurs. 2021, 47, 469–475. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | 60 |
| Age, M (SD) | 66.57 (4.78) |
| Age, range | 60–79 |
| Gender, n (%) | |
| Male | 28 (46.7) |
| Female | 32 (53.3) |
| Ethnicity, n (%) | |
| Hispanic or Latinx | 1 (1.7) |
| Not Hispanic or Latinx | 58 (96.7) |
| Unknown | 1 (1.7) |
| Race, n (%) | |
| Asian | 4 (6.7) |
| White | 55 (91.7) |
| More than one race | 1 (1.7) |
| Educational status, n (%) | |
| ≤High school or GED | 2 (3.4) |
| Some college or technical school | 11 (18.3) |
| 4-year college degree | 18 (30.0) |
| Post-baccalaureate degree | 29 (48.3) |
| Income, n (%) | |
| USD 20k–USD 39,999 | 2 (3.3) |
| USD 40k–USD 59,999 | 10 (16.7) |
| USD 60k–USD 79,999 | 16 (26.7) |
| USD 80k–USD 99,999 | 2 (3.3) |
| USD 100k–USD 120,999 | 10 (16.7) |
| USD 121k+ | 20 (33.3) |
| Measure | Total |
|---|---|
| N | 60 |
| DAS relationship satisfaction (0–50 scale), M (SD) | 39.50 (4.09) |
| Miller Social Intimacy Scale (1–10 scale), M (SD) | 8.13 (1.15) |
| Emotional Disclosure Scale: disclosure (1–5 scale), M (SD) | 2.35 (0.86) |
| Emotional Disclosure Scale: holding back (1–5 scale), M (SD) | 1.66 (0.73) |
| Communication Patterns Questionnaire: constructive communication (1–9 scale), M (SD) | 6.54 (1.39) |
| Sexual satisfaction in past month (0–10 scale), M (SD) | 5.86 (3.65) |
| Comorbidities, n (%) chronic conditions | |
| None | 34 (56.7) |
| One | 20 (33.3) |
| Two | 5 (8.3) |
| Three | 1 (1.7) |
| Current medication use, n (%) | |
| Prilosec | 8 (13.3) |
| Nexium | 2 (3.3) |
| Dexilant | 1 (1.7) |
| Pepcid | 5 (8.3) |
| Current supplement use, n (%) | |
| Prebiotic | 2 (3.3) |
| Probiotic | 9 (15.0) |
| Dietary intake per Dietary Screener Questionnaire, M (SD) | |
| Fiber, g/day | 16.97 (2.61) |
| Calcium, mg/day | 985.80 (202.33) |
| Added sugars, teaspoon equivalents/day | 15.62 (6.15) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Q.; Krajmalnik-Brown, R.; DiBaise, J.K.; Maldonado, J.; Guest, M.A.; Todd, M.; Langer, S.L. Relationship Functioning and Gut Microbiota Composition among Older Adult Couples. Int. J. Environ. Res. Public Health 2023, 20, 5435. https://doi.org/10.3390/ijerph20085435
Cheng Q, Krajmalnik-Brown R, DiBaise JK, Maldonado J, Guest MA, Todd M, Langer SL. Relationship Functioning and Gut Microbiota Composition among Older Adult Couples. International Journal of Environmental Research and Public Health. 2023; 20(8):5435. https://doi.org/10.3390/ijerph20085435
Chicago/Turabian StyleCheng, Qiwen, Rosa Krajmalnik-Brown, John K. DiBaise, Juan Maldonado, M. Aaron Guest, Michael Todd, and Shelby L. Langer. 2023. "Relationship Functioning and Gut Microbiota Composition among Older Adult Couples" International Journal of Environmental Research and Public Health 20, no. 8: 5435. https://doi.org/10.3390/ijerph20085435
APA StyleCheng, Q., Krajmalnik-Brown, R., DiBaise, J. K., Maldonado, J., Guest, M. A., Todd, M., & Langer, S. L. (2023). Relationship Functioning and Gut Microbiota Composition among Older Adult Couples. International Journal of Environmental Research and Public Health, 20(8), 5435. https://doi.org/10.3390/ijerph20085435