Ethanol Consumption by Wistar Rat Dams Affects Selenium Bioavailability and Antioxidant Balance in Their Progeny

Abstract
:1. Introduction
2. Results and Discussion
2.1. Dams' Weight Gain and Se Homeostasis
2.2. Pups' Weight at Birth and After Breastfeeding, and Pups' Se Homeostasis
2.3. Selenium Deposits in Different Tissues of Dams and Offspring (μg/g Dry Weight)
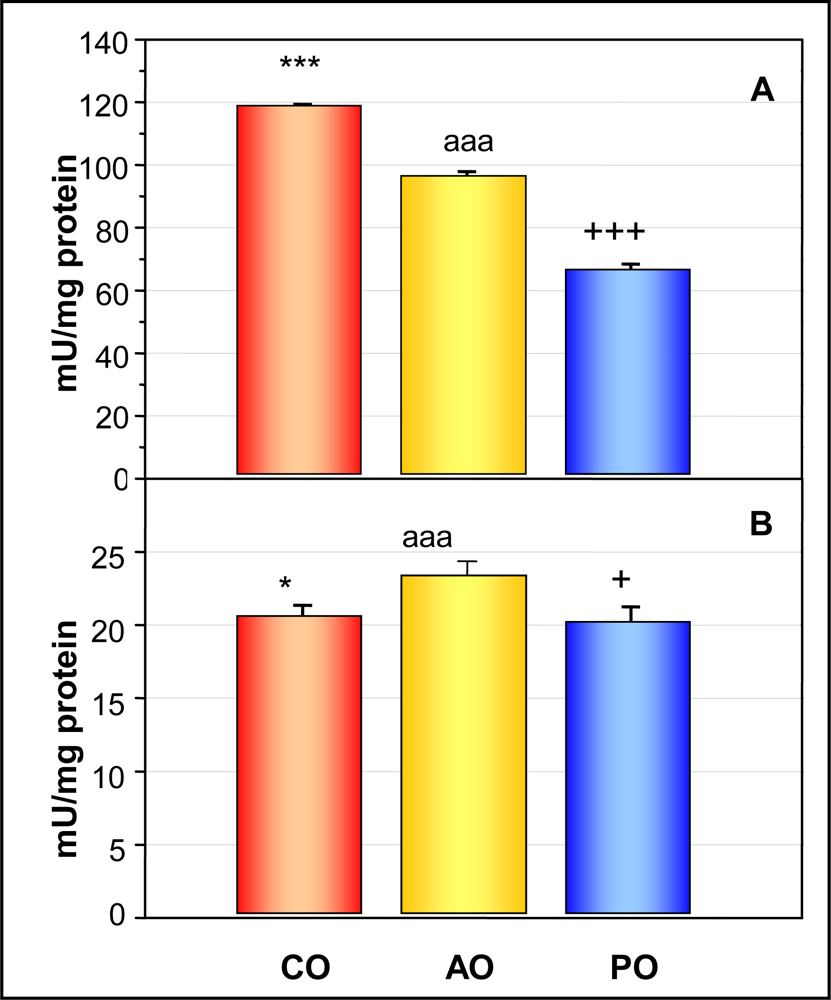
2.4. Offspring Hepatic Glutathione Peroxidase Activity (A) and Serum Glutathione Peroxidase Activity (B)
3. Experimental Section
4. Conclusions
Acknowledgments
References and Notes
- Bondy, SC; Pearson, KR. Ethanol-induced oxidative stress and nutritional status. Alcohol. Clin. Exp. Res 1993, 17, 651–654. [Google Scholar]
- Moreno Otero, R; Cortés, JR. Nutrition and chronic alcohol abuse. Nutr. Hosp 2008, 23, 3–7. [Google Scholar]
- Zhang, X; Sliwowska, JH; Weinberg, J. Prenatal alcohol exposure and foetal programming: effects on neuroendocrine and immune function. Exp. Biol. Med 2005, 230, 376–388. [Google Scholar]
- Youngentob, SL; Glendinning, JI. Foetal ethanol exposure increases ethanol intake by making it smell and taste better. Proc. Natl. Acad. Sci. USA 2009, 106, 5359–5364. [Google Scholar]
- Murillo-Fuentes, ML; Artillo, R; Ojeda, ML; Delgado, MJ; Murillo, ML; Carreras, O. Effects of prenatal or postnatal ethanol consumption on Zn intestinal absorption and excretion in rats. Alcohol Alcoholism 2007, 42, 3–10. [Google Scholar]
- Przybylska, J; Albera, E; Kankofer, M. Antioxidants in bovine colostrum. Reprod. Domest. Anim 2007, 42, 402–409. [Google Scholar]
- Rayman, MP. The importance of selenium to human health. The Lancet 2000, 356, 233–241. [Google Scholar]
- Bosl, MR; Takadu, K; Oshima, M; Nishimura, S; Taketo, MM. Early embryonic lethality caused by targeted disruption of the mouse selenocysteine tRNA gene (trsp). Proc. Natl. Acad. Sci. USA 1997, 94, 5531–5534. [Google Scholar]
- Dorea, JG. Selenium and breastfeeding. Br. J. Nut 2002, 88, 443–461. [Google Scholar]
- Ojeda, ML; Nogales, F; Vázquez, B; Delgado, MJ; Murillo, ML; Carreras, O. Alcohol, Gestation and Breastfeeding: Selenium as an Antioxidant Therapy. Alcohol Alcoholism 2009, 44, 272–277. [Google Scholar]
- Martín-Algarra, RV; Polache, A; Fernández-Villalba, E; Plá Delfina, JM; Guerri, C. Influence of chronic alcohol intake on intestinal taurine and antipyrine transport in pregnant rats. Alcohol. Clin. Exp. Res 1998, 22, 463–467. [Google Scholar]
- Harsley, JW; Oostdyk, TS; Keliher, PN. Determination of arsenic and selenium in environmental and agricultural samples by hydride generation atomic absorption spectrometry. J. Assc. Off. Anal. Chem 1988, 71, 1090–1093. [Google Scholar]
- Griffiths, NM; Stewart, RD; Robinson, MF. The metabolism of [75Se] selenomethionine in four women. Br. J. Nutr 1976, 35, 373–382. [Google Scholar]
- Martin, RF; Janghorbani, M; Young, VR. Experimental selenium restriction in healthy adult humans: changes in selenium metabolism studied with stable-isotope methodology. Am. J. Clin. Nutr 1989, 49, 854–61. [Google Scholar]
- Lorenzo Alonso, MJ; Bermejo Barrera, A; Cocho de Juan, JA; Fraga Bermúdez, JM; Bermejo Barrera, P. Selenium levels in related biological samples: human placenta, maternal and umbilical cord blood, hair and claws. J. Trace. Elem. Med. Biol 2005, 19, 49–54. [Google Scholar]
- Özdemir, HS; Karadas, F; Pappas, AC; Cassey, P; Oto, G; Tuncer, O. The selenium levels of mothers and their neonates using hair, breast milk, meconium, and maternal and umbilical cord blood in van basin. Biol. Trace. Elem. Res 2008, 122, 206–215. [Google Scholar]
- Salbe, AD; Levander, OA. Effect of various dietary factors on the deposition of selenium in the hair and claws of rats. J. Nutr 1990, 120, 200–206. [Google Scholar]
- Behne, D; Gessner, H; Kyriakopoulos, A. Information on the selenium status of several body compartments of rats from the selenium concentrations in blood fractions, hair and claws. J. Trace. Elem. Med. Biol 1996, 10, 174–179. [Google Scholar]
- Watzl, B; Watson, RR. Role of nutrients in alcohol-induced immunomodulation. Alcohol Alcoholism 1993, 28, 89–95. [Google Scholar]
- Kyriakopoulos, A; Richter, A; Pohl, T; Wolf, C; Grbavac, I; Plotnikov, A; Kühbacher, M; Bertelsmann, H; Behne, D. Protein expression in the tissues of the cardiovascular system of the rat under selenium deficiency and adequate conditions. Ann. N. Y. Acad. Sci 2007, 1095, 300–304. [Google Scholar]
- Kimura, T; Kawasaki, Y; Okumura, F; Sone, T; Natsuki, R; Isobe, M. Ethanol-induced expression of glutamate-cysteine ligase catalytic subunit gene is mediated by NF-kappaB. Toxicol. Lett 2009, 185, 110–115. [Google Scholar]
- McConnell, KP; Roth, DM. Respiratory excretion of selenium. Proc. Soc. Exp. Biol. Med 1966, 123, 919–921. [Google Scholar]
- Wentzel, P; Eriksson, UJ. Ethanol-induced foetal dysmorphogenesis in the mouse is diminished by high antioxidative capacity of the mother. Toxicol. Sci 2006, 92, 416–422. [Google Scholar]
- Payne, RL; Southern, LL. Changes in glutathione peroxidase and tissue selenium concentrations of broilers after consuming a diet adequate in selenium. Poult. Sci 2005, 84, 1268–1276. [Google Scholar]
- Ojeda, ML; Delgado-Villa, MJ; Llopis, R; Murillo, ML; Carreras, O. Lipid metabolism in ethanol-treated rat pups and adults: Effects of folic acid. Alcohol Alcoholism 2008, 43, 544–550. [Google Scholar]
- Lawrence, RA; Burk, RF. Glutathione peroxidase activity in selenium-deficient rat liver. Biochem. Biophys. Res. Commun 1976, 71, 952–958. [Google Scholar]
- Lowry, OH; Rosebrough, NJ; Farr, AL; Randall, RJ. Protein measurement with the Folin phenol reagent. J. Biol. Chem 1951, 1, 265–275. [Google Scholar]


{kind=link}
| CD (n:6) | AD (n:6) | PD (n:6) | |
|---|---|---|---|
| Weight gain (g) | 78.3±7.1** | 46.6±4.2 | 49.5±5.1++ |
| Se intake during gestation (μg/d) | 1.89±0.21* | 1.43±0.02 | 1.45±0.04+ |
| Se intake during lactation (μg/d) | 3.12±0.33* | 2.23±0.03 | 2.22±0.05+ |
| Se (μg/d) in faeces | 0.26±0.02*** | 0.14±0.001 | 0.12±0.01+++ |
| Se (μg/d) in urine | 0.18±0.01 | 0.23±0.02aaa | 0.11±0.01++ |
| Se (μg/l) in serum | 245 ± 15.2 | 259 ± 21.6aaa | 150 ± 5.1+++ |
| Se (ng/g dry weight) in claws | 30 ± 3.6 | 30 ± 1.7aaa | 66 ± 2.5+++ |
| Se (ng/g dry weight)in hair | 50 ± 0.5*** | 16 ± 1.0aaa | 113 ± 8.0+++ |
| Apparent Se absorption rate (%) | 91.6±0.74 | 93.3±0.64 | 94.45±0.55+ |
| Apparent Se balance (μg/d) | 2.67±0.07*** | 1.85±0.008aa | 2.01±0.013+++ |
| CO (n:8) | AO (n:8) | PO (n:8) | |
|---|---|---|---|
| Weight at birth (g) | 5.53 ± 0.09 | 5.02 ± 0.18 | 5.14 ± 0.16 |
| Weight at 21 days (g) | 31.60 ± 1.05** | 22.00 ± 1.67a | 27.80 ± 1.15+ |
| Nº of offspring/ litter | 11.0±0.9* | 7.8±0.6 | 9.0 ± 0.8 |
| Se (p.p.m.) in milk | 20 ± 1.2*** | 12.6 ± 0.6 | 13.7 ± 0.7+++ |
| Milk intake during 30 min suckling (mL) | 0.85 ±0.045*** | 0.54 ± 0.03 | 0.60 ± 0.04+++ |
| Se intake during 30 min suckling (ng) | 17.05 ± 0.1*** | 6.85 ± 0.03aaa | 8.63 ± 0.04+++ |
| Se (ng/d) in faeces | 14 ± 0.1 | 18 ± 0.1 | 13 ± 0.1 |
| Se (ng/d) in urine | 15 ± 0.1 | 18 ± 0.1aaa | 5 ± 0.05+++ |
| Se (μg/L) in serum | 117 ± 3.4** | 137 ± 6.9aaa | 71.01±1.5+++ |
| Se (ng/g dry weight)in hair | 88 ± 0.7* | 67 ± 0.5aaa | 108 ± 0.7 |
| Se levels (μg/g dry weight) | CD (n: 6) | AD (n: 6) | PD (n: 6) | CO (n: 8) | AO (n: 8) | PO (n: 8) |
|---|---|---|---|---|---|---|
| Heart | 0.22±0.03** | 0.27±0.05aaa | 0.14±0.02+++ | 0.24 ± 0.01** | 0.19 ± 0.01 | 0.13± 0.01+++ |
| Liver | 0.21±0.02* | 0.28±0.02a | 0.23±0.01 | 0.39±0.02*** | 0.30 ±0.01aaa | 0.15±0.01+++ |
| Kidney | 0.66±0.04 | 0.63±0.04aaa | 0.37±0.0+++ | 0.46±0.01*** | 0.37 ± 0.02aaa | 0.26±0.01+++ |
| Lung | 0.14±0.008 | 0.13±0.003 | 0.12±0.007+ | 0.29±0.02 | 0.29±0.02 | 0.33±0.02 |
| Spleen | 0.28±0.01* | 0.37±0.02aa | 0.26±0.01 | 0.24±0.02 | 0.24±0.02 | 0.24±0.01 |
| Muscle | 0.15±0.01** | 0.12±0.003 | 0.1±0.006+++ | |||
| Mammary gland | 0.18±0.01** | 0.13±0.005 | 0.14±0.005+ | |||
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ojeda, M.L.; Vázquez, B.; Nogales, F.; Murillo, M.L.; Carreras, O. Ethanol Consumption by Wistar Rat Dams Affects Selenium Bioavailability and Antioxidant Balance in Their Progeny. Int. J. Environ. Res. Public Health 2009, 6, 2139-2149. https://doi.org/10.3390/ijerph6082139
Ojeda ML, Vázquez B, Nogales F, Murillo ML, Carreras O. Ethanol Consumption by Wistar Rat Dams Affects Selenium Bioavailability and Antioxidant Balance in Their Progeny. International Journal of Environmental Research and Public Health. 2009; 6(8):2139-2149. https://doi.org/10.3390/ijerph6082139
Chicago/Turabian StyleOjeda, María Luisa, Beatriz Vázquez, Fátima Nogales, María Luisa Murillo, and Olimpia Carreras. 2009. "Ethanol Consumption by Wistar Rat Dams Affects Selenium Bioavailability and Antioxidant Balance in Their Progeny" International Journal of Environmental Research and Public Health 6, no. 8: 2139-2149. https://doi.org/10.3390/ijerph6082139
APA StyleOjeda, M. L., Vázquez, B., Nogales, F., Murillo, M. L., & Carreras, O. (2009). Ethanol Consumption by Wistar Rat Dams Affects Selenium Bioavailability and Antioxidant Balance in Their Progeny. International Journal of Environmental Research and Public Health, 6(8), 2139-2149. https://doi.org/10.3390/ijerph6082139