Updating the Northern Tsetse Limit in Burkina Faso (1949–2009): Impact of Global Change

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
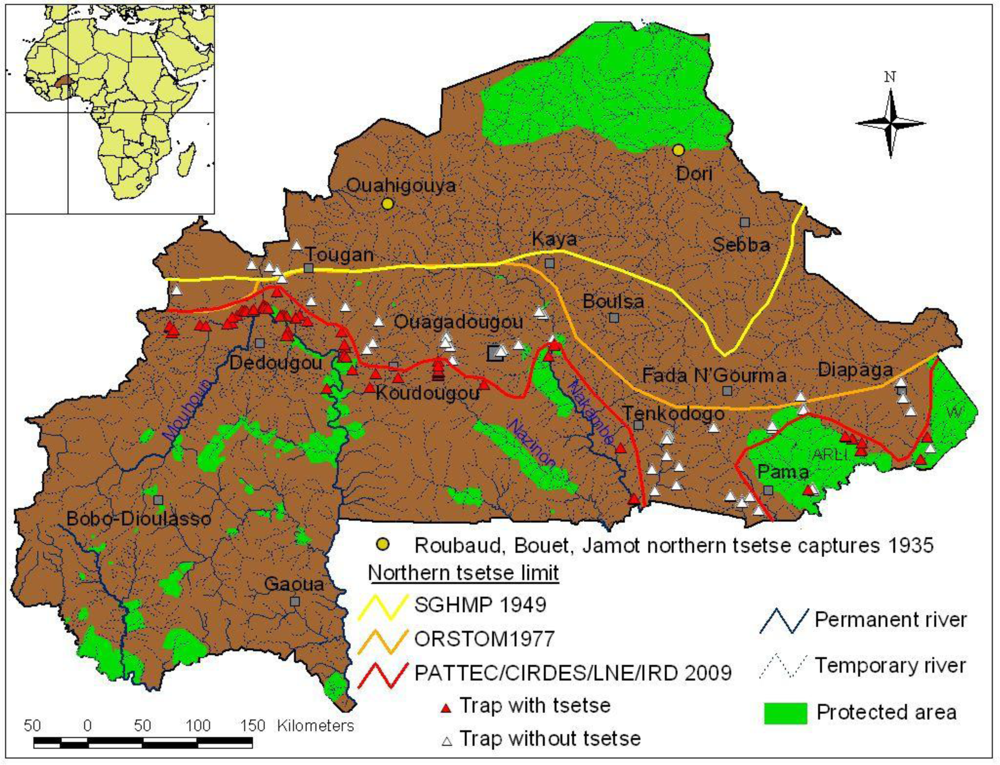
2.1. Previous Northern Tsetse Limits
2.2. Current Northern Tsetse Limit in Burkina Faso
2.3. Impact of Global Change on Tsetse Distribution
3. Experimental Section
4. Conclusions
Acknowledgments
References
- Desquesnes, M; Bossard, G; Patrel, D; Herder, S; Patout, O; Lepetitcolin, E; Thevenon, S; Berthier, D; Pavlovic, R; Brugidou, R; Jacquiet, P; Schelcher, F; Faye, B; Touratier, L; Cuny, G. First outbreak of Trypanosoma evansi in camels in metropolitan France. Vet. Rec 2008, 162, 750–752. [Google Scholar]
- Budd, L. Economic analysis. In DFID-funded Tsetse and Trypanosomiasis Research and Development Since 1980; Department for International Development: Chatham, UK, 1999. [Google Scholar]
- Solano, P; Ravel, S; de Meeûs, T. How can tsetse population genetics contribute to African Trypanosomiasis control. Trends Parasitol 2010, in press. [Google Scholar]
- Kabayo, JP. Aiming to eliminate tsetse from Africa. Trends Parasitol 2002, 18, 473–475. [Google Scholar]
- Nash, TAM. Tsetse Flies in British West Africa; H.M.S.O.: London, UK, 1948. [Google Scholar]
- Solano, P; Bouyer, J; Itard, J; Cuisance, D. Cyclical vectors of Trypanosomosis. In Infectious and Parasitic Diseases of Livestock; Lefèvre, B, Chermette, U, Eds.; Lavoisier Tec&Doc: Paris, France, 2009. [Google Scholar]
- Bouyer, J; Balenghien, T; Ravel, S; Vial, L; Sidibé, I; Thévenon, S; Solano, P; De Meeûs, T. Population sizes and dispersal patterns of tsetse flies: rolling on the river? Mol. Ecol 2009, 18, 2787–2797. [Google Scholar]
- Challier, A. Ecologie de Glossina palpalis gambiensis Vanderplank, 1949 (Diptera-Muscidae) en savane d’Afrique occidentale; ORSTOM: Paris, France, 1973. [Google Scholar]
- Gruvel, J. Données générales sur l’écologie de Glossina tachinoides Westwood, 1850, dans la réserve de Kalamaloué, vallée du Bas-Chari. Rev. Elev. Méd. vét. Pays trop 1975, 28, 27–40. [Google Scholar]
- Bouyer, J; Guerrini, L; César, J; De La Rocque, S; Cuisance, D. A phyto-sociological analysis of the distribution of riverine tsetse flies in Burkina Faso. Med. Vet. Entomol 2005, 19, 372–378. [Google Scholar]
- De la Rocque, S; Rioux, JA. Influence des changements climatiques sur l’épidémiologie des maladies transmissibles. Bull. Soc. Path. Exo 2008, 101, 213–219. [Google Scholar]
- Githeko, AK; Lindsay, SW; Confalonieri, UE; Patz, JA. Climate change and vector borne disease: a regional analysis. Bull. WHO 2000, 78, 1136–1147. [Google Scholar]
- Rogers, DJ; Randolph, SE. Climate change and vector-borne diseases. Adv. Parasitol 2006, 62, 346–381. [Google Scholar]
- Courtin, F; Jamonneau, V; Duvallet, G; Garcia, A; Coulibaly, B; Doumenge, JP; Cuny, G; Solano, P. Sleeping sickness in West Africa (1906–2006): changes in spatial repartition and lessons from the past. Trop. Med. Int. Health 2008, 13, 334–344. [Google Scholar]
- Rouamba, J; Jamonneau, V; Sidibé, I; Solano, P; Courtin, F. Impact de la dynamique de peuplement sur la distribution des glossines dans la boucle du Mouhoun (Burkina Faso). Parasite 2009, 16, 11–19. [Google Scholar]
- Rayaisse, JB; Courtin, F; Akoudjim, M; Cesar, J; Solano, P. Influence de l’anthropisation sur la végétation et l’abondance des tsé-tsé au sud du Burkina Faso. Parasite 2009, 16, 21–28. [Google Scholar]
- Courtin, F; Sidibé, I; Rouamba, J; Jamonneau, V; Gouro, A; Solano, P. Impacts observés des évolutions démo-climatiques sur la répartition spatiale des hommes, des tsé-tsé et des trypanosomoses en Afrique de l’Ouest. Parasite 2009, 16, 3–10. [Google Scholar]
- Ford, J; Katondo, KM. The Distribution of Tsetse Flies (Glossina) in Africa; Organization of African Unity, Scientific and Technical Research Commission (OAU/STRC): Nairobi, Kenya, 1973. [Google Scholar]
- Wint, W; Rogers, D. Predicted Distributions of Tsetse in Africa Consultancy Report for the Animal Health Service of the Animal Production and Health Division of the Food and Agriculture Organization of the United Nations; FAO: Rome, Italy, 2000. Available online: http://www.fao.org/ag/againfo/programmes/en/paat/documents/maps/pdf/tserep.pdf (accessed on 15 December 2009).
- Cecchi, G; Mattioli, RC; Slingenbergh, J; de la Rocque, S. Land cover and tsetse fly distributions in sub-Saharan Africa. Med. Vet. Entomol 2008, 22, 364–373. [Google Scholar]
- Cecchi, G; Mattioli, RC. Global geospatial datasets for African trypanosomiasis management: a review. Program against African Trypanosomiasis Technical and Scientific Series 2009, 9, 1–39. [Google Scholar]
- Guerrini, L; Sidibé, I; Bouyer, J. Tsetse distribution in the Mouhoun river basin (Burkina Faso): the role of global and local geospatial datasets. Program against African Trypanosomiasis Technical and Scientific Series 2009, 9, 41–52. [Google Scholar]
- Challier, A; Laveissière, C. Carte de répartition des glossines en Haute-Volta; ORSTOM: Bondy, France, 1977. Available online: http://www.cartographie.ird.fr/sphaera/images/telechargement/00891.pdf (accessed on 4 December 2009).
- Gouzien, P. La maladie du sommeil dans le Haut-Sénégal et Niger. n° 490/; Document Technique OCCGE: Bobo-Dioulasso, Burkina Faso, 1908; p. 25. [Google Scholar]
- Roubaud, E; Bouët, G; Jamot, E. Distribution des maladies à trypanosomoses et des mouches tsé-tsé en Afrique Occidentale Française: Carte au 1/ 4 000 000; dressée par le Service Géographique de l’Afrique Occidentale Française: Paris, France, 1935. [Google Scholar]
- Service Général d’Hygiène Mobile et de Prophylaxie. Limite des aires de distribution des Glossina palpalis, Glossina tachinoides et des Glossina morsitans submorsitans en Afrique Occidentale Française, carte au 1 : 3,000,000 dressée, dessinée, imprimée et publiée par le Service Géographique de l’AOF: Paris, France, 1949.
- Barclay, HJ; Hargrove, JW. Probability models to facilitate a declaration of pest-free status, with special reference to tsetse (Diptera: Glossinidae). Bull. Ent. Res 2005, 95, 1–11. [Google Scholar]
- Gado, BA. Une histoire des famines au Sahel, étude des grandes crises alimentaires (XIXe-XXe siècles); Eds l’Harmattan: Paris, France, 1993. [Google Scholar]
- L’Hôte, Y; Mahé, G. Afrique de l’Ouest et Centrale, précipitations moyennes annuelles (période 1951–1989); ORSTOM: Bondy, France, 1996. Available online: http://www.cartographie.ird.fr/images/pluvio_afrique/pluvio3.gif (accessed on 15 September 2009).
- Mahé, G; Olivry, JC. Changements climatiques et variations des écoulements en Afrique occidentale et centrale, du mensuel à l’interannuel. Hydrology for the Water Management of Large River Basins 1991, 201, 163–171. [Google Scholar]
- Paturel, JE; Servat, E; Delattre, MO. Analyse de séries pluviométriques de longue durée en Afrique de l’Ouest et Centrale non sahélienne dans un contexte de variabilité climatique. Hydrol. Sci 1998, 43, 937–946. [Google Scholar]
- Laveissière, C. Répartition des glossines en Haute-Volta : effets de la grande sécheresse de 1972–1973. Cahiers ORSTOM, sér Ent Méd Parasitol 1976, XIV, 293–299. [Google Scholar]
- Mandé, I. Les migrations du travail en Haute-Volta (actuel Burkina Faso), mise en perspective historique (1919–1960); PhD thesis; Université Denis Diderot: Paris, France, 1997; p. 490. [Google Scholar]
- Guengant, JP. La démographie entre convergences et divergences. Ed. B. Ferry; In l’Afrique face à ses défis démographiques, un avenir incertain; AFD-CEPED-Karthala: Paris, France, 2007; pp. 27–121. [Google Scholar]
- Capron, J; Kohler, JM. Répartition de la population mossi en Haute-Volta. ORSTOM, Office de la recherche scientifique et technique outre mer: Bondy, France, 1996.
- .
- Ben Yahmed, D; Houstin, N; Delaroche, F; Ménager, MT; Meunier-Nikiéma, A; Arnaud, JC; Devey, M; Fournet, F; Goama Nacoulma, A; Ouattara, A; Toguyéni, R. Atlas de l’Afrique; Jeune Afrique: Paris, France, 2005; p. 115. [Google Scholar]
- Reid, SR; Kruska, RL; Deichmann, U; Thornton, PK; Leak, S. Human population growth and the extinction of the tsetse fly. Agric. Ecosyst. Env 2000, 77, 227–236. [Google Scholar]
- Laveissière, C; Traoré, T. Enquête entomologique dans le foyer de trypanosomiase humaine de la Somone (République du Sénégal, mai 1979); Centre Muraz, rapport de mission n° 7166/79, Document Technique OCCGE: Bobo-Dioulasso, Burkina Faso, 1976; p. 12. [Google Scholar]
- Kubi, C; Van Den Abbeele, J; De Deken, R; Marcotty, T; Dorny, P; Van den Bossche, P. The effect of starvation on the susceptibility of teneral and non teneral tsetse flies to trypanosome infection. Med. Vet. Entomol 2006, 20, 388–392. [Google Scholar]
- D’Orgeval, T. Impact du changement climatique sur la saison des pluies en Afrique de l’Ouest : que nous disent les modèles de climat actuels? Sécheresse 2008, 19, 79–85. [Google Scholar]
- AMMA. Devic, MP, Roussot, O, Janicot, S, Thorncroft, C, Eds.; 2009; p. 601.
- United Nations. World Population Prospects: The 2008 Revision. Population Database, United Nations Population Division: New York, NY, USA, 2009. Available online: http://esa.un.org/unpp/index.asp (accessed on 15 November 2009).
- Koudougou, Z; Sidibé, I; Tamboura, I. Collection of entomological baseline data in the Mouhoun river basin (Burkina Faso): the use of GIS, remote sensing and GPS. Programme against African Trypanosomiasis Technical and Scientific Series 2009, 9, 53–59. [Google Scholar]
- Challier, A; Eyraud, M; Lafaye, A; Laveissière, C. Amélioration du rendement du piège biconique pour glossines (Diptera: Glossinidae) par l’emploi d’un cône inférieur bleu. Cahiers ORSTOM, sér Ent Méd Parasitol 1977, XV, 283–286. [Google Scholar]
- Rogers, D. Tsetse population dynamics and distribution: a new analytical approach. J. Anim. Ecol 1979, 48, 825–849. [Google Scholar]



© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Courtin, F.; Rayaissé, J.-B.; Tamboura, I.; Serdébéogo, O.; Koudougou, Z.; Solano, P.; Sidibé, I. Updating the Northern Tsetse Limit in Burkina Faso (1949–2009): Impact of Global Change. Int. J. Environ. Res. Public Health 2010, 7, 1708-1719. https://doi.org/10.3390/ijerph7041708
Courtin F, Rayaissé J-B, Tamboura I, Serdébéogo O, Koudougou Z, Solano P, Sidibé I. Updating the Northern Tsetse Limit in Burkina Faso (1949–2009): Impact of Global Change. International Journal of Environmental Research and Public Health. 2010; 7(4):1708-1719. https://doi.org/10.3390/ijerph7041708
Chicago/Turabian StyleCourtin, Fabrice, Jean-Baptiste Rayaissé, Issa Tamboura, Oumar Serdébéogo, Zowindé Koudougou, Philippe Solano, and Issa Sidibé. 2010. "Updating the Northern Tsetse Limit in Burkina Faso (1949–2009): Impact of Global Change" International Journal of Environmental Research and Public Health 7, no. 4: 1708-1719. https://doi.org/10.3390/ijerph7041708
APA StyleCourtin, F., Rayaissé, J. -B., Tamboura, I., Serdébéogo, O., Koudougou, Z., Solano, P., & Sidibé, I. (2010). Updating the Northern Tsetse Limit in Burkina Faso (1949–2009): Impact of Global Change. International Journal of Environmental Research and Public Health, 7(4), 1708-1719. https://doi.org/10.3390/ijerph7041708