Basic Apoptotic Mechanisms of Lead Toxicity in Human Leukemia (Hl-60) Cells



Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals and Media
2.2. Cell/Tissue Culture
2.3. Cell Treatment and Measurement of Cell Viability
2.4. Annexin V FITC/PI Binding Assay by Flow Cytometry
2.5. Active Caspase-3 Assay by Flow Cytometry
2.6. Statistical Analysis
3. Results and Discussion
3.1. Cell Viability
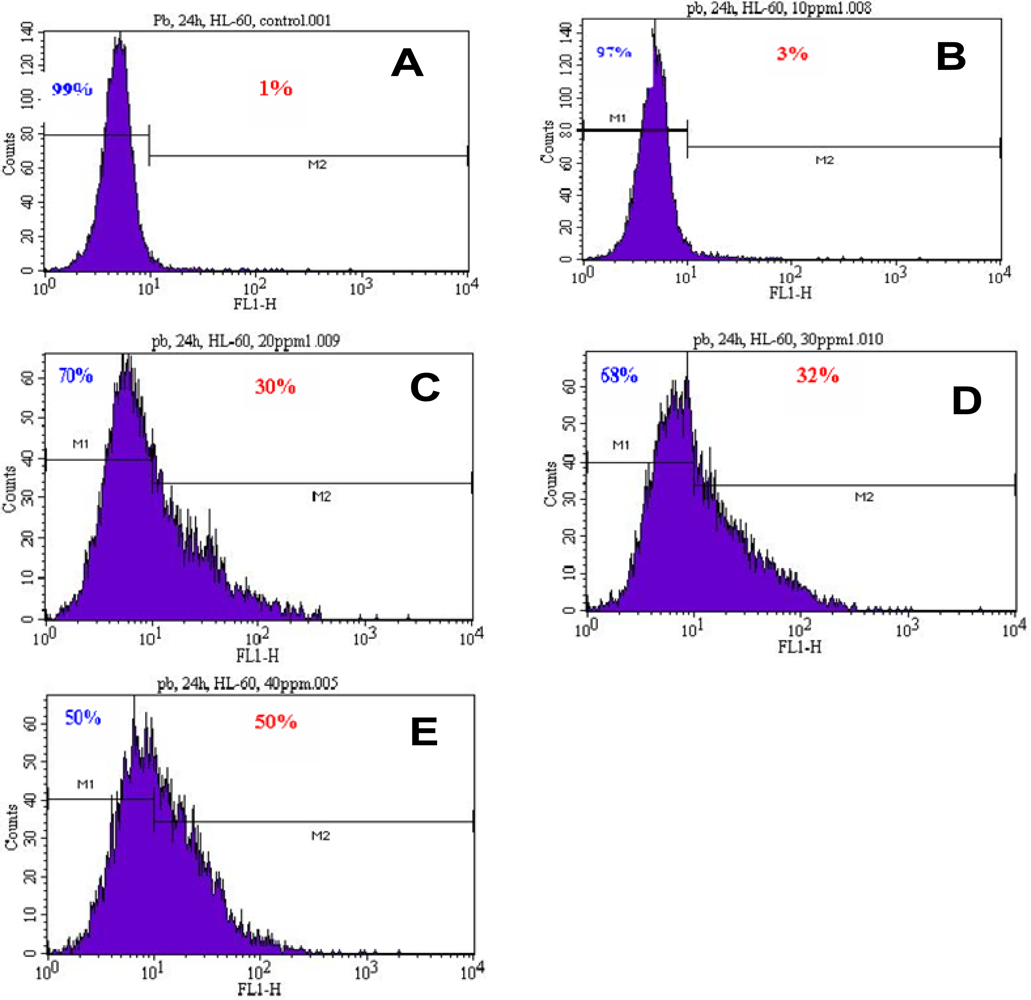
3.2. Induction of Phosphatidylserine Externalization
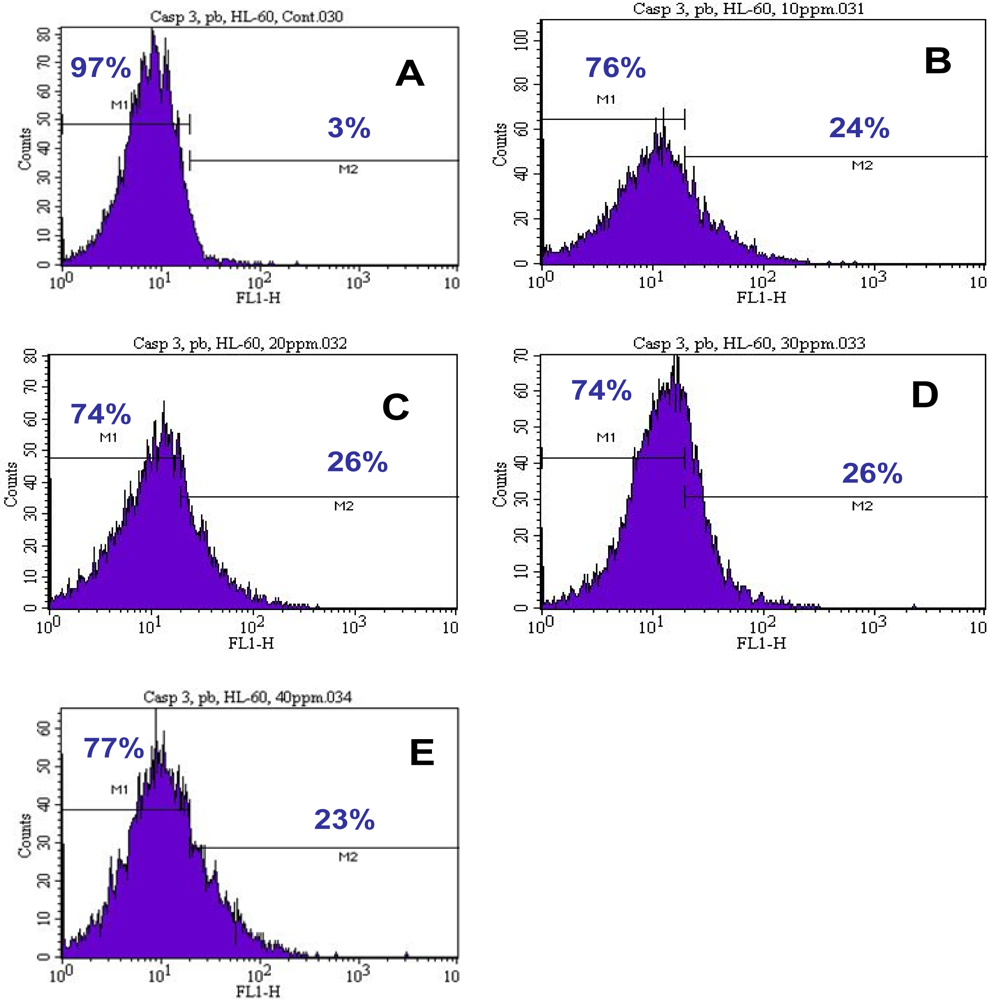
3.3. Activation of Caspase-3
4. Conclusions
Acknowledgments
References
- Rosen, JF. Adverse effects of lead at low exposure levels: trends in the treatment of childhood lead poisoning. Toxicology 1995, 97, 11–17. [Google Scholar]
- Apostoli, P; Boffetta, P. Why a conference on lead toxicity? Introductory remarks to the proceedings of the international conference on lead exposure. Amer. J. Infect. Control 2000, 38, 229–230. [Google Scholar]
- Bressler, J; Kim, KA; Chakraborti, T; Goldstein, G. Mechanism of lead neurotoxicity. Neurochem. Res 1999, 24, 595–600. [Google Scholar]
- Yedjou, C; Steverson, M; Tchounwou, P. Lead nitrate-induced oxidative stress in human liver carcinoma (HepG2) cells. Metal Ions Biol. Med 2006, 9, 293–297. [Google Scholar]
- Yedjou, CG; Tchounwou, CK; Haile, S; Edwards, F; Tchounwou, PB. NAC protection against DNA damage: N-acetyl-cysteine (NAC) protects against DNA damage associated with lead toxicity in HepG2 cells. Ethnicity and Disease 2010, 20, 101–103. [Google Scholar]
- Sharifi, AM; Baniasadi, S; Jorjani, M; Rahimi, F; Bakhshayesh, M. Investigation of acute lead poisoning on apoptosis in rat hippocampus in vivo. Neurosci. Lett 2002, 329, 45–48. [Google Scholar]
- Tavakoli-Nezhad, M; Barron, AJ; Pitts, DK. Postnatal inorganic lead exposure decreases the number of spontaneously active midbrain dopamine neurons in the rat. Neurotoxicology 2001, 22, 259–269. [Google Scholar]
- Adhikari, N; Sinha, N; Narayan, R; Saxena, DK. Lead-induced cell death in testes of young rats. J. Appl. Toxicol 2001, 21, 275–277. [Google Scholar]
- Iavicoli, I; Carelli, G; Sgambato, A. Lead inhibits growth and induces apoptosis in normal rat fibroblasts. Altern. Lab. Anim 2001, 29, 461–469. [Google Scholar]
- Shabani, A; Rabbani, A. Lead nitrate induced apoptosis in alveolar macrophages from rat lung. Toxicology 2000, 149, 109–114. [Google Scholar]
- He, L; Perkins, GA; Poblenz, AT; Harris, JB; Hung, M; Ellisman, MH; Fox, DA. Bcl-xL overexpression blocks bax-mediated mitochondrial contact site formation and apoptosis in rod photoreceptors of lead-exposed mice. Proc. Natl. Acad. Sci 2003, 100, 1022–1027. [Google Scholar]
- He, L; Poblenz, AT; Medrano, CJ; Fox, DA. Lead and calcium produce rod photoreceptor cell apoptosis by opening the mitochondrial permeability transition pore. J. Biol. Chem 2000, 275, 12175–12184. [Google Scholar]
- Tchounwou, PB; Yedjou, CG; Foxx, D; Ishaque, A; Shen, E. Lead induced cytotoxicity and transcriptional activation of stress genes in human liver carcinoma cells. Mol. Cell. Biochem 2004, 255, 161–170. [Google Scholar]
- Tully, DB; Collins, BJ; Overstreet, JD; Smith, CS; Dinse, GE; Mumtaz, MM; Chapin, RE. Effects of arsenic, cadmium, chromium and lead on gene expression regulated by a battery of 13 different promoters in recombinant HepG2 cells. Toxicol. Appl. Pharmacol 2000, 168, 79–90. [Google Scholar]
- Tchounwou, PB; Yedjou, CG; Dorsey, WC. Arsenic trioxide induced transcriptional activation and expression of stress genes in human liver carcinoma cells (HepG2). Cell. Mol. Biol 2003, 49, 1071–1079. [Google Scholar]
- Sengupta, M; Bishayi, B. Effect of lead and arsenic on murine macrophage response. Drug Chem. Toxicol 2002, 25, 459–472. [Google Scholar]
- Centers for Disease control (CDC). Fatal pediatric lead poisoning. New Hampshire, 2000. Morb. Mort. Weekly Rep 2001, 50, 457–459. [Google Scholar]
- Salvesen, GS; Abrams, JM. Caspase activation-stepping on the gas or releasing the brakes? Lessons from humans and flies. Oncogene 2004, 23, 2774–2784. [Google Scholar]
- Woo, M; Hakem, R; Soengas, MS; Duncan, GS; Shahinian, A; Kagi, D; Hakem, A; McCurrach, M; Khoo, W; Kaufman, SA; Senaldi, G; Howard, T; Lowe, SW; Mak, TW. Essential contribution of caspase 3/CPP32 to apoptosis and its associated nuclear changes. Genes. Develop 1998, 12, 806–819. [Google Scholar]
- Fox, DA; Campbell, ML; Blocker, YS. Functional alterations and apoptotic cell death in the retina following developmental or adult lead exposure. Neurotoxicology 1997, 18, 645–664. [Google Scholar]
- Oberto, A; Marks, N; Evans, HL; Guidotti, A. Lead (Pb+2) promotes apoptosis in newborn rat cerebellar neurons: pathological implications. J. Pharmacol. Exp. Ther 1996, 279, 435–442. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Concentrations | Annexin V Negative Cells or Viable Cells (Mean ± SD)% | Annexin V Positive Cells or Apoptotic Cells (Mean ± SD)% |
|---|---|---|
| 0μg/mL | 99.00 ± 0.00 | 1.00 ± 0.00 |
| 10μg/mL | 97.00 ± 0.28 | 3.00 ± 0.28 |
| 20μg/mL | 66.46 ± 5.15* | 33.54 ± 5.15* |
| 30μg/mL | 61.50 ± 8.56* | 38.50 ± 8.56* |
| 40μg/mL | 51.85 ± 3.04* | 48.15 ± 3.04* |
| Concentrations | Caspase-3 Negative Cells or Viable Cells (Mean ± SD)% | Caspase-3 Positive Cells or Apoptotic Cells (Mean ± SD)% |
|---|---|---|
| 0 μg/mL | 97.00 ± 0.00 | 3.00 ± 0.00 |
| 10 μg/mL | 69.25 ± 10.25* | 30.75 ± 10.25* |
| 20 μg/mL | 63.50 ± 14.84* | 36.5 0 ± 14.84* |
| 30 μg/mL | 67.75 ± 8.13* | 32.25 ± 8.13* |
| 40 μg/mL | 67.50 ± 13.57* | 32.50 ± 13.57* |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yedjou, C.G.; Milner, J.N.; Howard, C.B.; Tchounwou, P.B. Basic Apoptotic Mechanisms of Lead Toxicity in Human Leukemia (Hl-60) Cells. Int. J. Environ. Res. Public Health 2010, 7, 2008-2017. https://doi.org/10.3390/ijerph7052008
Yedjou CG, Milner JN, Howard CB, Tchounwou PB. Basic Apoptotic Mechanisms of Lead Toxicity in Human Leukemia (Hl-60) Cells. International Journal of Environmental Research and Public Health. 2010; 7(5):2008-2017. https://doi.org/10.3390/ijerph7052008
Chicago/Turabian StyleYedjou, Clement G., Jessica N. Milner, Carolyn B. Howard, and Paul B. Tchounwou. 2010. "Basic Apoptotic Mechanisms of Lead Toxicity in Human Leukemia (Hl-60) Cells" International Journal of Environmental Research and Public Health 7, no. 5: 2008-2017. https://doi.org/10.3390/ijerph7052008
APA StyleYedjou, C. G., Milner, J. N., Howard, C. B., & Tchounwou, P. B. (2010). Basic Apoptotic Mechanisms of Lead Toxicity in Human Leukemia (Hl-60) Cells. International Journal of Environmental Research and Public Health, 7(5), 2008-2017. https://doi.org/10.3390/ijerph7052008