RIOK2 Contributes to Cell Growth and Protein Synthesis in Human Oral Squamous Cell Carcinoma

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Tissue Samples
2.2. Antibodies
2.3. Tissue Microarrays and Fluorescence Immunohistochemistry
2.4. Cell Culture
2.5. siRNA Transfection
2.6. Western Blotting
2.7. ATP-Based Cell Growth Assay
2.8. Clonogenic Assay
2.9. Protein Synthesis Assay
2.10. Statistical Analysis
3. Results
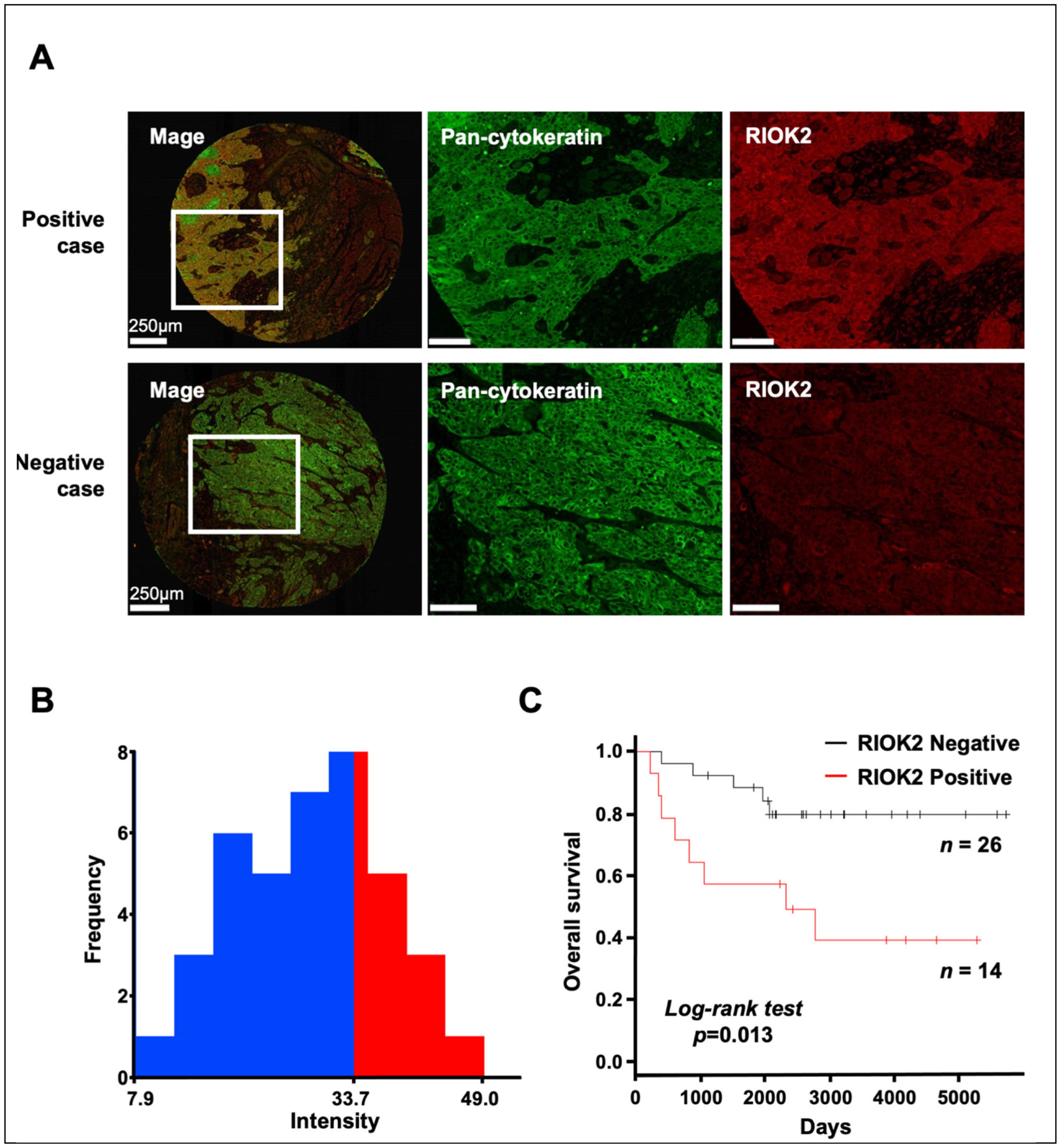
3.1. Prognostic Impact of RIOK2 Protein Expression in TSCC Patients
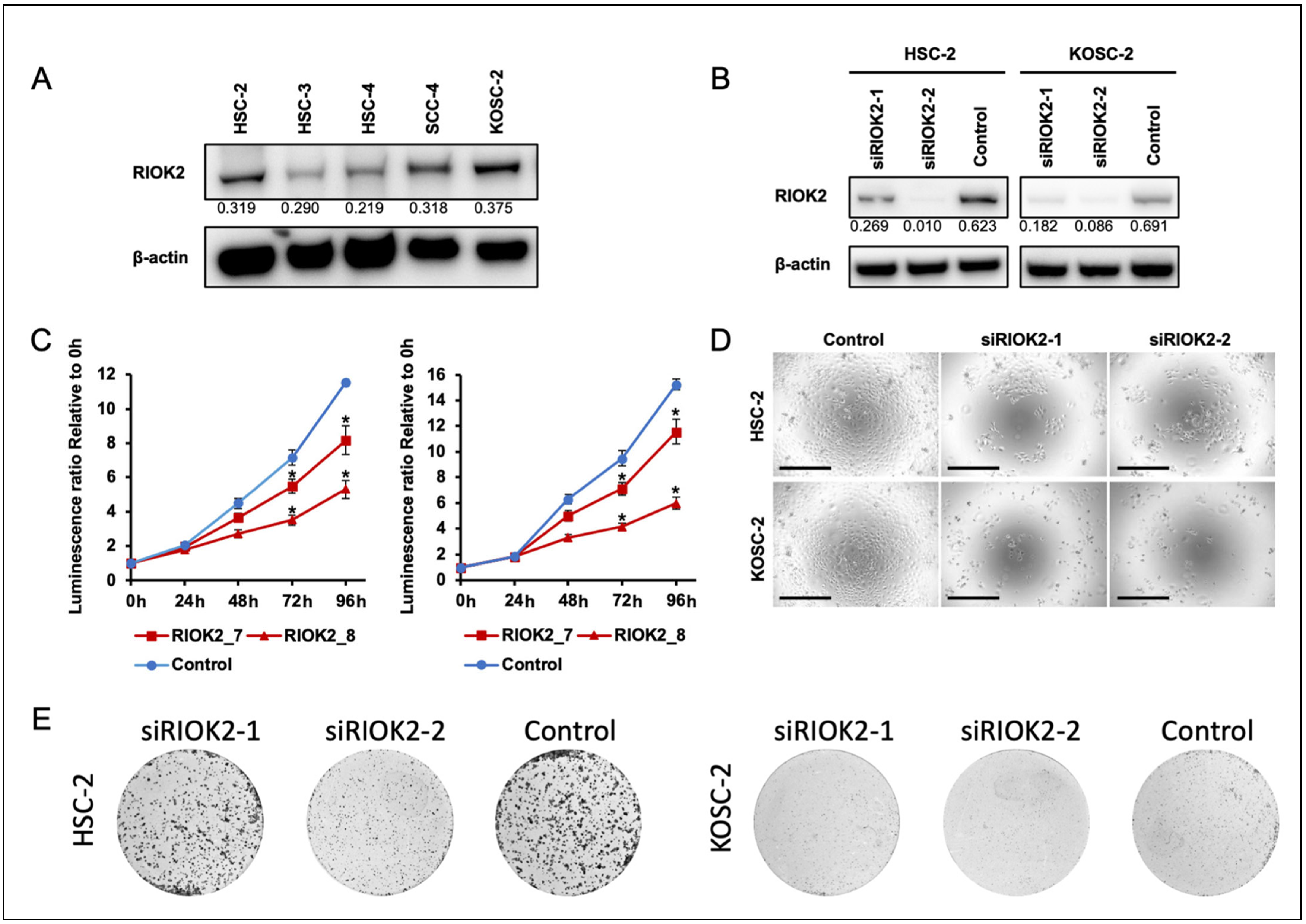
3.2. Effects of RIOK2 Expression on Oral Cancer Cell Growth
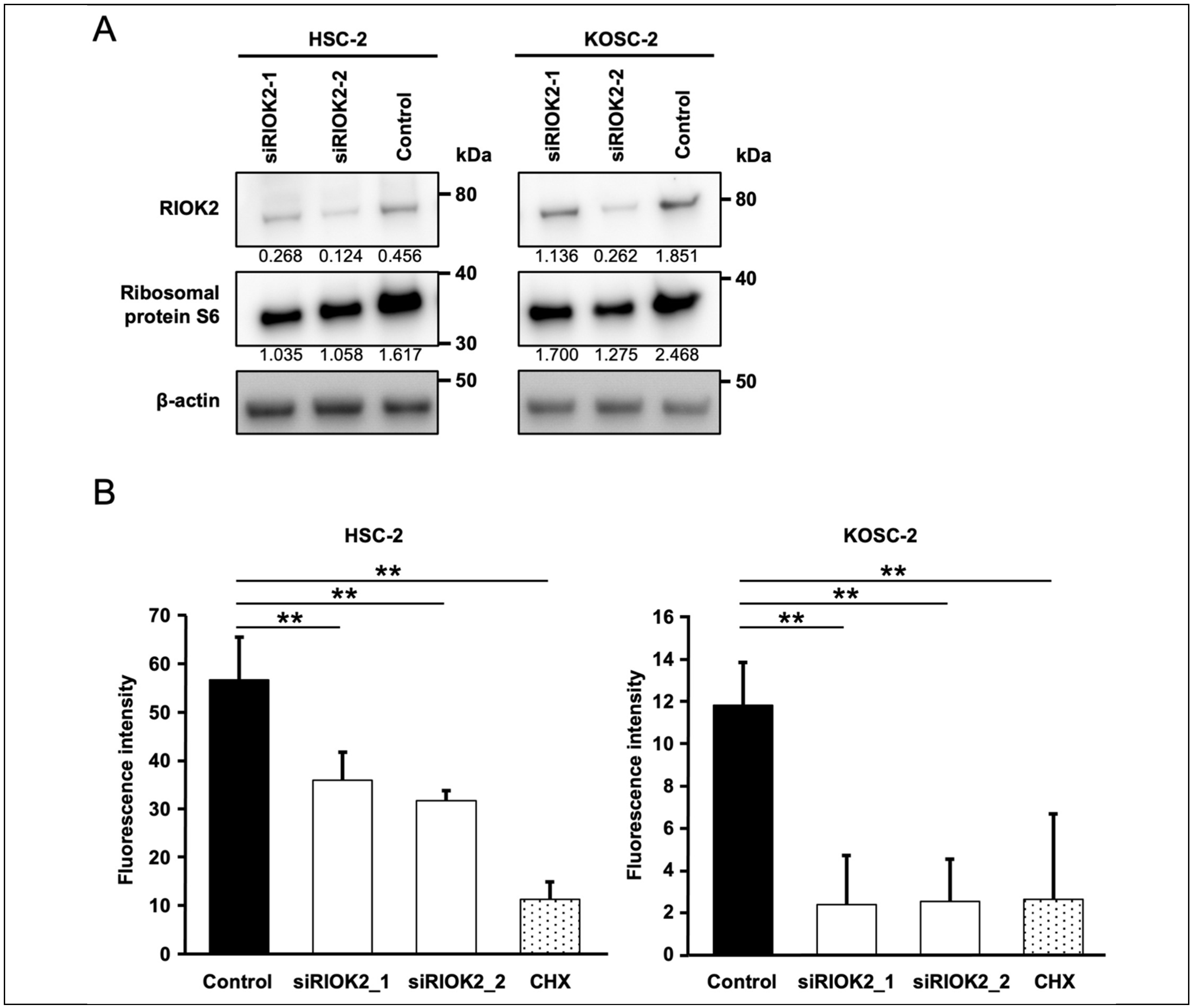
3.3. RIOK2 Regulated Protein Synthesis in Oral Cancer Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Primers 2020, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Itai, S.; Ohishi, T.; Kaneko, M.K.; Yamada, S.; Abe, S.; Nakamura, T.; Yanaka, M.; Chang, Y.W.; Ohba, S.I.; Nishioka, Y.; et al. Anti-podocalyxin antibody exerts antitumor effects via antibody-dependent cellular cytotoxicity in mouse xenograft models of oral squamous cell carcinoma. Oncotarget 2018, 9, 22480–22497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brocklehurst, P.R.; Baker, S.R.; Speight, P.M. Oral cancer screening: What have we learnt and what is there still to achieve? Future Oncol. 2010, 6, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.; Thomas, G.; Volarević, S. Ribosome biogenesis in cancer: New players and therapeutic avenues. Nat. Rev. Cancer 2018, 18, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Burger, K.; Mühl, B.; Harasim, T.; Rohrmoser, M.; Malamoussi, A.; Orban, M.; Kellner, M.; Gruber-Eber, A.; Kremmer, E.; Hölzel, M.; et al. Chemotherapeutic drugs inhibit ribosome biogenesis at various levels. J. Biol. Chem. 2010, 285, 12416–12425. [Google Scholar] [CrossRef] [Green Version]
- Brighenti, E.; Treré, D.; Derenzini, M. Targeted cancer therapy with ribosome biogenesis inhibitors: A real possibility? Oncotarget 2015, 6, 38617–38627. [Google Scholar] [CrossRef] [Green Version]
- Zemp, I.; Wild, T.; O’Donohue, M.F.; Wandrey, F.; Widmann, B.; Gleizes, P.E.; Kutay, U. Distinct cytoplasmic maturation steps of 40S ribosomal subunit precursors require hRio2. J. Cell Biol. 2009, 185, 1167–1180. [Google Scholar] [CrossRef] [Green Version]
- LaRonde-LeBlanc, N.; Wlodawer, A. The RIO kinases: An atypical protein kinase family required for ribosome biogenesis and cell cycle progression. Biochim. Biophys. Acta 2005, 1754, 14–24. [Google Scholar] [CrossRef]
- Ferreira-Cerca, S.; Sagar, V.; Schäfer, T.; Diop, M.; Wesseling, A.M.; Lu, H.; Chai, E.; Hurt, E.; LaRonde-LeBlanc, N. ATPase-dependent role of the atypical kinase Rio2 on the evolving pre-40S ribosomal subunit. Nat. Struct. Mol. Biol. 2012, 19, 1316–1323. [Google Scholar] [CrossRef] [Green Version]
- Ameismeier, M.; Cheng, J.; Berninghausen, O.; Beckmann, R. Visualizing late states of human 40S ribosomal subunit maturation. Nature 2018, 558, 249–253. [Google Scholar] [CrossRef]
- Asquith, C.R.M.; East, M.P.; Zuercher, W.J. RIOK2: Straddling the kinase/ATPase line. Nat. Rev. Drug Discov. 2019, 18, 574. [Google Scholar] [CrossRef]
- Liu, K.; Chen, H.L.; Wang, S.; Gu, M.M.; Chen, X.M.; Zhang, S.L.; Yu, K.J.; You, Q.S. High Expression of RIOK2 and NOB1 Predict Human Non-small Cell Lung Cancer Outcomes. Sci. Rep. 2016, 6, 28666. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Chen, H.; You, Q.; Ye, Q.; Wang, F.; Wang, S.; Zhang, S.; Yu, K.; Li, W.; Gu, M. miR-145 inhibits human non-small-cell lung cancer growth by dual-targeting RIOK2 and NOB1. Int. J. Oncol. 2018, 53, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Li, C.; Jin, L.; Xing, J.; Sha, Z.; Zhang, T.; Ji, D.; Yu, R.; Gao, S. RIOK2 is negatively regulated by miR-4744 and promotes glioma cell migration/invasion through epithelial-mesenchymal transition. J. Cell Mol. Med. 2020, 24, 4494–4509. [Google Scholar] [CrossRef] [Green Version]
- Messling, J.E.; Agger, K.; Andersen, K.L.; Kromer, K.; Kuepper, H.M.; Lund, A.H.; Helin, K. Targeting RIOK2 ATPase activity leads to decreased protein synthesis and cell death in acute myeloid leukemia. Blood 2022, 139, 245–255. [Google Scholar] [CrossRef]
- Honda, K.; Hishiki, T.; Yamamoto, S.; Yamamoto, T.; Miura, N.; Kubo, A.; Itoh, M.; Chen, W.Y.; Takano, M.; Yoshikawa, T.; et al. On-tissue polysulfide visualization by surface-enhanced Raman spectroscopy benefits patients with ovarian cancer to predict post-operative chemosensitivity. Redox Biol. 2021, 41, 101926. [Google Scholar] [CrossRef]
- Onidani, K.; Miura, N.; Sugiura, Y.; Abe, Y.; Watabe, Y.; Kakuya, T.; Mori, T.; Yoshimoto, S.; Adachi, J.; Kiyoi, T.; et al. Possible Therapeutic Strategy Involving the Purine Synthesis Pathway Regulated by ITK in Tongue Squamous Cell Carcinoma. Cancers 2021, 13, 3333. [Google Scholar] [CrossRef]
- Watabe, Y.; Mori, T.; Yoshimoto, S.; Nomura, T.; Shibahara, T.; Yamada, T.; Honda, K. Copy number increase of ACTN4 is a prognostic indicator in salivary gland carcinoma. Cancer Med. 2014, 3, 613–622. [Google Scholar] [CrossRef]
- Camp, R.L.; Dolled-Filhart, M.; Rimm, D.L. X-tile: A new bio-informatics tool for biomarker assessment and outcome-based cut-point optimization. Clin. Cancer Res. 2004, 10, 7252–7259. [Google Scholar] [CrossRef] [Green Version]
- Miura, N.; Kamita, M.; Kakuya, T.; Fujiwara, Y.; Tsuta, K.; Shiraishi, H.; Takeshita, F.; Ochiya, T.; Shoji, H.; Huang, W.; et al. Efficacy of adjuvant chemotherapy for non-small cell lung cancer assessed by metastatic potential associated with ACTN4. Oncotarget 2016, 7, 33165–33178. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awasthi, S.; Verma, M.; Mahesh, A.; MI, K.K.; Govindaraju, G.; Rajavelu, A.; Chavali, P.L.; Chavali, S.; Dhayalan, A. DDX49 is an RNA helicase that affects translation by regulating mRNA export and the levels of pre-ribosomal RNA. Nucleic Acids Res. 2018, 46, 6304–6317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, Y.W.; You, K.S.; Park, J.S.; Lee, S.G.; Seong, Y.S. Ribosomal Protein S6: A Potential Therapeutic Target against Cancer? Int. J. Mol. Sci. 2021, 23, 48. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, Y.; Stoleru, D.; Salic, A. Imaging protein synthesis in cells and tissues with an alkyne analog of puromycin. Proc. Natl. Acad. Sci. USA 2012, 109, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Luo, H.; Wang, L.; Li, H.; Liang, Y.; Xia, J.; Wang, Z.; Cheng, B.; Huang, L.; Liao, G.; et al. Combined inhibition of RNA polymerase I and mTORC1/2 synergize to combat oral squamous cell carcinoma. Biomed. Pharmacother. 2021, 133, 110906. [Google Scholar] [CrossRef]
- Read, R.D.; Fenton, T.R.; Gomez, G.G.; Wykosky, J.; Vandenberg, S.R.; Babic, I.; Iwanami, A.; Yang, H.; Cavenee, W.K.; Mischel, P.S.; et al. A kinome-wide RNAi screen in Drosophila Glia reveals that the RIO kinases mediate cell proliferation and survival through TORC2-Akt signaling in glioblastoma. PLoS Genet. 2013, 9, e1003253. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Hu, X.; Yan, J.; Wang, Y.; Lu, F.; Chang, J. RIOK2 Inhibitor NSC139021 Exerts Anti-Tumor Effects on Glioblastoma via Inducing Skp2-Mediated Cell Cycle Arrest and Apoptosis. Biomedicines 2021, 9, 1244. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Xavier, C.P.; Sukumar, G.; Tan, S.H.; Ravindranath, L.; Seraj, N.; Kumar, V.; Sreenath, T.; McLeod, D.G.; Petrovics, G.; et al. Identification of a Small Molecule That Selectively Inhibits ERG-Positive Cancer Cell Growth. Cancer Res. 2018, 78, 3659–3671. [Google Scholar] [CrossRef] [Green Version]
- Ruvinsky, I.; Meyuhas, O. Ribosomal protein S6 phosphorylation: From protein synthesis to cell size. Trends Biochem. Sci. 2006, 31, 342–348. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| RIOK2 IHC | |||||
|---|---|---|---|---|---|
| Number of Cases (%) | Negative | Positive | p-Value a | ||
| Total | 40 (100) | 26 | 14 | ||
| Age (years) | |||||
| <62 | 19 (47.5) | 15 | 4 | 0.1050 | |
| ≥62 | 21 (52.5) | 11 | 10 | ||
| Sex | |||||
| Male | 15 (37.5) | 7 | 8 | 0.08940 | |
| Female | 25 (62.5) | 19 | 6 | ||
| Stage b | |||||
| I | 21 (52.5) | 14 | 7 | 0.1860 | |
| II | 19 (47.5) | 12 | 7 | ||
| Histologic differentiation | |||||
| Well | 26 (65) | 20 | 6 | 0.04280 * | |
| Poor/Moderate | 14 (35) | 6 | 8 | ||
| Mode of invasion c | |||||
| 1,2 | 23 (57.5) | 17 | 6 | 0.1980 | |
| 3,4 | 17 (42.5) | 9 | 8 | ||
| Perineural invasion | |||||
| Negative | 39 (97.5) | 25 | 14 | 1.000 | |
| Positive | 1 (2.5) | 1 | 0 | ||
| Lymphovascular invasion | |||||
| Negative | 31 (77.5) | 22 | 9 | 0.2340 | |
| Positive | 9 (22.5) | 4 | 5 | ||
| Covariate | Univariate Analysis (n = 40) | Multivariate Analysis (n = 40) | |||||
|---|---|---|---|---|---|---|---|
| HR | 95%CI | p-Value | HR | 95%CI | p-Value | ||
| Age (year) | |||||||
| <62 | Reference | ||||||
| ≥62 | 3.514 | 0.9648–12.80 | 0.05671 | ||||
| Sex | |||||||
| Male | Reference | ||||||
| Female | 0.9959 | 0.3248–3.054 | 0.9943 | ||||
| Stage a | |||||||
| I | Reference | ||||||
| II | 1.445 | 0.4850–4.306 | 0.5086 | ||||
| Differentiation | |||||||
| Well | Reference | ||||||
| Poor/ Moderate | 2.527 | 0.8459–7.547 | 0.09686 | ||||
| Mode of invasion b | |||||||
| 1,2 | Reference | ||||||
| 3,4 | 2.620 | 0.8539–8.036 | 0.09686 | ||||
| Lymphovascular invasion | |||||||
| Negative | Reference | Reference | |||||
| Positive | 3.130 | 1.017–9.632 | 0.04660 * | 2.871 | 0.9218–8.941 | 0.06884 | |
| RIOK2 IHC | |||||||
| Negative | Reference | Reference | |||||
| Positive | 3.752 | 1.225–11.49 | 0.02063 * | 3.539 | 1.149–10.91 | 0.02772 * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsuzaki, Y.; Naito, Y.; Miura, N.; Mori, T.; Watabe, Y.; Yoshimoto, S.; Shibahara, T.; Takano, M.; Honda, K. RIOK2 Contributes to Cell Growth and Protein Synthesis in Human Oral Squamous Cell Carcinoma. Curr. Oncol. 2023, 30, 381-391. https://doi.org/10.3390/curroncol30010031
Matsuzaki Y, Naito Y, Miura N, Mori T, Watabe Y, Yoshimoto S, Shibahara T, Takano M, Honda K. RIOK2 Contributes to Cell Growth and Protein Synthesis in Human Oral Squamous Cell Carcinoma. Current Oncology. 2023; 30(1):381-391. https://doi.org/10.3390/curroncol30010031
Chicago/Turabian StyleMatsuzaki, Yusuke, Yutaka Naito, Nami Miura, Taisuke Mori, Yukio Watabe, Seiichi Yoshimoto, Takahiko Shibahara, Masayuki Takano, and Kazufumi Honda. 2023. "RIOK2 Contributes to Cell Growth and Protein Synthesis in Human Oral Squamous Cell Carcinoma" Current Oncology 30, no. 1: 381-391. https://doi.org/10.3390/curroncol30010031
APA StyleMatsuzaki, Y., Naito, Y., Miura, N., Mori, T., Watabe, Y., Yoshimoto, S., Shibahara, T., Takano, M., & Honda, K. (2023). RIOK2 Contributes to Cell Growth and Protein Synthesis in Human Oral Squamous Cell Carcinoma. Current Oncology, 30(1), 381-391. https://doi.org/10.3390/curroncol30010031