
Biological Consequences of Marine Energy Development on Marine Animals




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
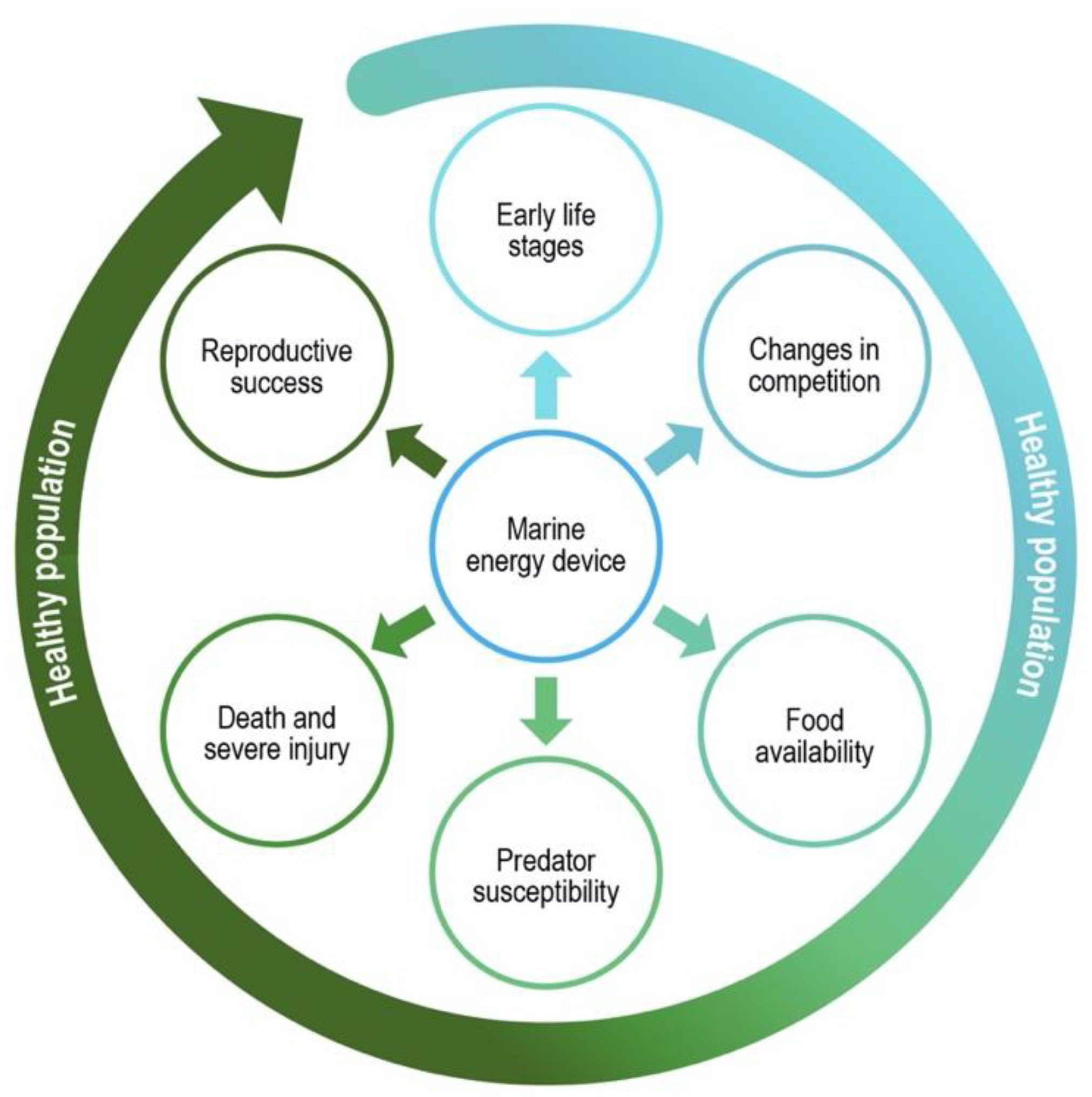
- Success of early life stages, through development of larvae, juveniles, or young marine animals.
- Changes in competitive capabilities, through the ability of marine animals to effectively compete for mates, food, or space resources.
- Growth and survival based on food availability, allowing individuals and populations to thrive.
- Susceptibility to predators, through the ability of marine animals to avoid or escape predators.
- Injury or death from an encounter that may remove specific individuals from populations of marine animals.
- Reproductive success, through the ability of marine animals to mate and rear young to support their populations.
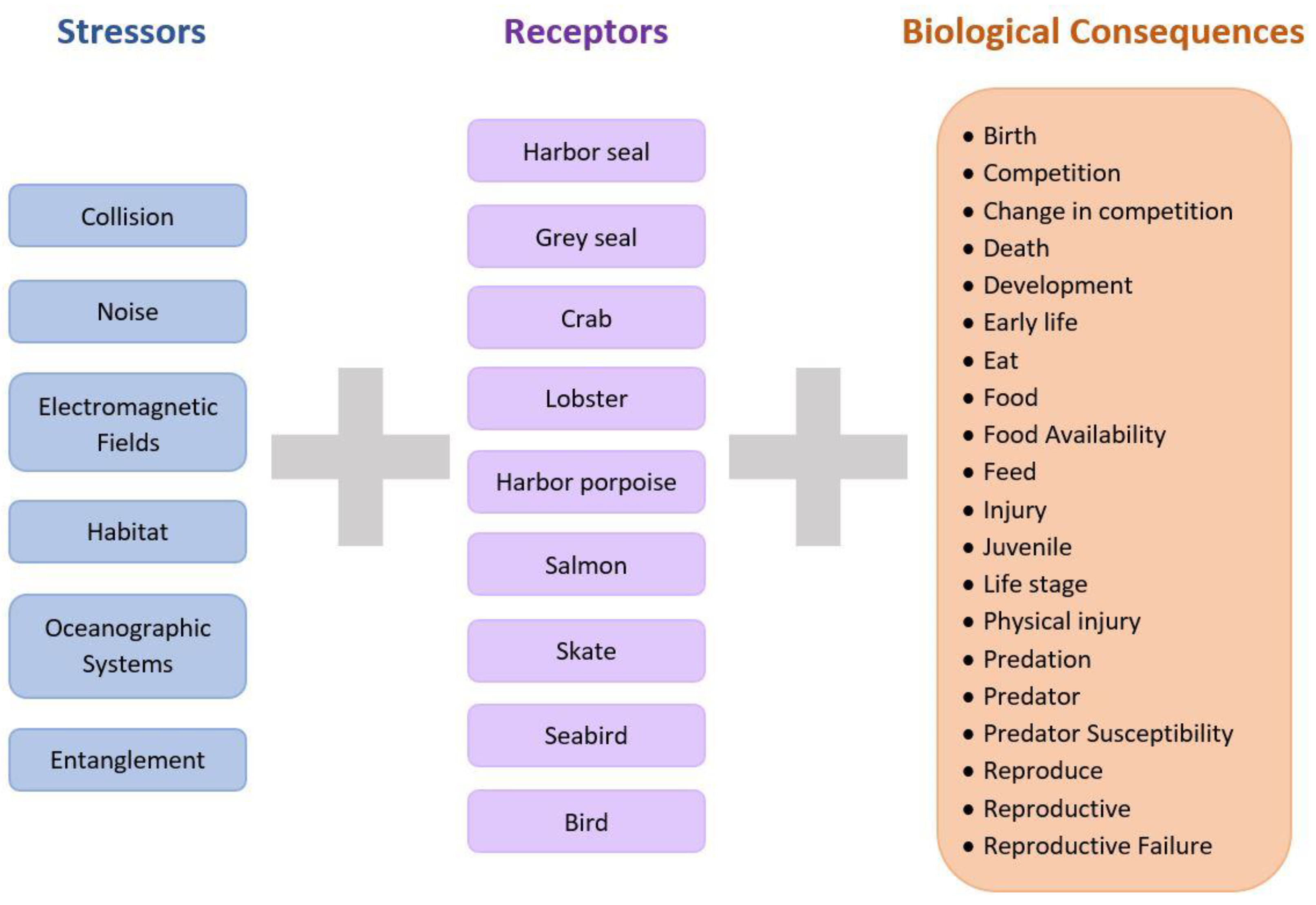
2. Methods
3. Marine Energy Effects on Seals
4. Marine Energy Effects on Crabs and Lobsters
5. Marine Energy Effects on Other Receptors
5.1. Harbor Porpoise
5.2. Salmon
5.3. Skates
5.4. Diving Seabirds
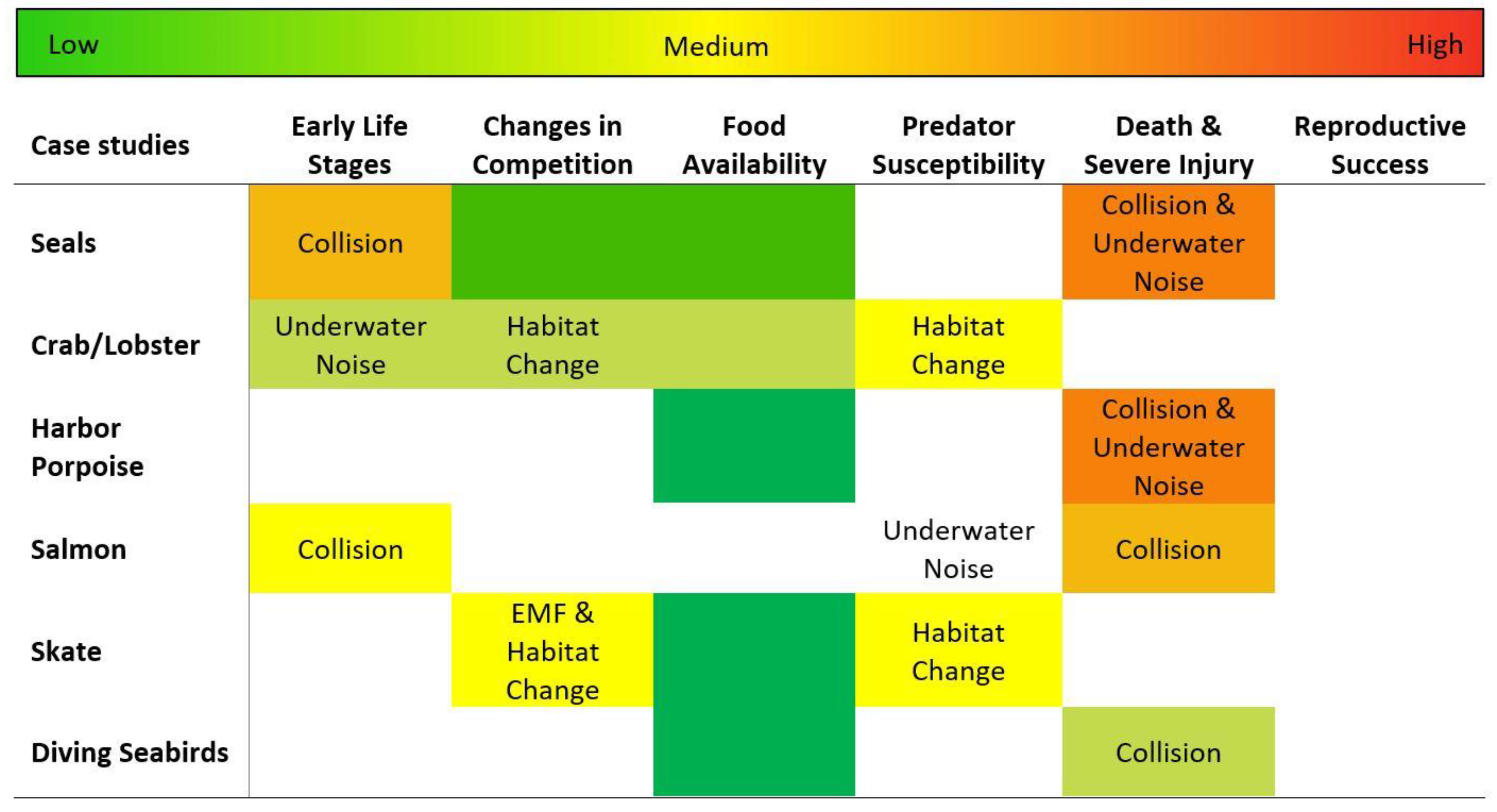
6. Risk to Species from Marine Energy Development
- High risk = likely to interact with severe consequences
- Medium-high risk = high likelihood of interaction, outcomes could be severe
- Medium risk = reasonable likelihood of interaction, outcomes could be detrimental
- Medium-low risk = low likelihood of interaction, outcomes not likely to be severe
- Low risk = unlikely to interact
- Insufficient information to make an estimate.
7. Discussion
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- IRENA Renewable Energy Statistics 2019; The International Renewable Energy Agency: Abu Dhabi, United Arab Emirates, 2019; pp. 1–398. ISBN 978-92-9260-137-9.
- Ocean Energy Systems IEA-OES Annual Report: An Overview of Ocean Energy Activities in 2020; The Executive Committee of Ocean Energy Systems: Lisbon, Portugal, 2021; pp. 1–188.
- Copping, A.E.; Hemery, L.G. OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 1–327. [Google Scholar]
- Prittimaa, L.; Cruz, E. Ocean Energy and the Environment: Research and Strategic Actions. Report to the European Commission; European Technology and Innovation Platform for Ocean Energy (ETIP): Brussel, Belgium, 2020; p. 36. [Google Scholar]
- Boehlert, G.; Gill, A. Environmental and Ecological Effects of Ocean Renewable Energy Development—A Current Synthesis. Oceanography 2010, 23, 68–81. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, D.; Palmer, L.; Macaulay, J.; Sparling, C.; Hastie, G. Harbour porpoises exhibit localized evasion of a tidal turbine. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 2459–2468. [Google Scholar] [CrossRef]
- Harwood, J.; King, S. The Sensitivity of UK Marine Mammal Populations to Marine Renewables Developments; Report No. SMRUL-NER-2012-027; SMRU Consulting: St Andrews, UK, 2014; pp. 1–53. [Google Scholar]
- Malinka, C.; Gillespie, D.; Macaulay, J.; Joy, R.; Sparling, C. First in situ passive acoustic monitoring for marine mammals during operation of a tidal turbine in Ramsey Sound, Wales. Mar. Ecol. Prog. Ser. 2018, 590, 247–266. [Google Scholar] [CrossRef] [Green Version]
- Williamson, B.J.; Blondel, P.; Williamson, L.D.; Scott, B.E. Application of a multibeam echosounder to document changes in animal movement and behaviour around a tidal turbine structure. ICES J. Mar. Sci. 2021, 78, 1253–1266. [Google Scholar] [CrossRef]
- Copping, A.E.; Hemery, L.G.; Viehman, H.; Seitz, A.C.; Staines, G.J.; Hasselman, D.J. Are fish in danger? A review of environmental effects of marine renewable energy on fishes. Biol. Conserv. 2021, 262, 109297. [Google Scholar] [CrossRef]
- Hastie, G.D.; Russell, D.J.F.; Benjamins, S.; Moss, S.; Wilson, B.; Thompson, D. Dynamic habitat corridors for marine predators; intensive use of a coastal channel by harbour seals is modulated by tidal currents. Behav. Ecol. Sociobiol. 2016, 70, 2161–2174. [Google Scholar] [CrossRef] [Green Version]
- Lieber, L.; Nimmo-Smith, W.A.M.; Waggitt, J.J.; Kregting, L. Fine-scale hydrodynamic metrics underlying predator occupancy patterns in tidal stream environments. Ecol. Indic. 2018, 94, 397–408. [Google Scholar] [CrossRef] [Green Version]
- SMRU Ltd. Utilisation of Space by Grey and Harbour Seals in the Pentland Firth and Orkney Waters; Scottish Natural Heritage: Oban, UK, 2011; p. 62. [Google Scholar]
- Benjamins, S.; Harnois, V.; Smith, H.; Johanning, L.; Greenhill, L.; Carter, C.; Wilson, B. Understanding the Potential for Marine Megafauna Entanglement Risk from Marine Renewable Energy Developments; Scottish Natural Heritage: Perth, UK, 2014; p. 95. [Google Scholar]
- Minesto Deep Green Holyhead Deep Project Phase I (0.5 MW)—Environmental Statement (Report No. L-100194-S14-EIAS-001); Minesto: Holyhead, UK, 2016; pp. 1–487.
- Garavelli, L. Encounters of Marine Animals with Marine Renewable Energy Device Mooring Systems and Subsea Cables. In OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development around the World; Copping, A.E., Hemery, L.G., Eds.; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 146–153. [Google Scholar] [CrossRef]
- Muelbert, M.M.C.; Bowen, W.D. Duration of lactation and postweaning changes in mass and body composition of harbour seal, Phoca vitulina, pups. Can. J. Zool. 1993, 71, 1405–1414. [Google Scholar] [CrossRef]
- Thompson, D. Annex I: Movements and Diving Behaviour of Juvenile Grey Seals in Areas of High Tidal Energy; RPS Group: Chepstow, UK, 2012; p. 55. [Google Scholar]
- Stansbury, A.L.; Götz, T.; Deecke, V.B.; Janik, V.M. Grey seals use anthropogenic signals from acoustic tags to locate fish: Evidence from a simulated foraging task. Proc. R. Soc. B Boil. Sci. 2015, 282, 20141595. [Google Scholar] [CrossRef] [Green Version]
- Fraser, S.; Williamson, B.J.; Nikora, V.; Scott, B.E. Fish distributions in a tidal channel indicate the behavioural impact of a marine renewable energy installation. Energy Rep. 2018, 4, 65–69. [Google Scholar] [CrossRef]
- Hemery, L. Changes in Benthic and Pelagic Habitats Caused by Marine Renewable Energy Devices. In OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World; Copping, A.E., Hemery, L.G., Eds.; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 105–125. [Google Scholar] [CrossRef]
- Arnould, J.P.Y.; Monk, J.; Ierodiaconou, D.; Hindell, M.A.; Semmens, J.; Hoskins, A.J.; Costa, D.P.; Abernathy, K.; Marshall, G.J. Use of Anthropogenic Sea Floor Structures by Australian Fur Seals: Potential Positive Ecological Impacts of Marine Industrial Development? PLoS ONE 2015, 10, e0130581. [Google Scholar] [CrossRef] [Green Version]
- Williamson, B.; Fraser, S.; Williamson, L.; Nikora, V.; Scott, B. Predictable changes in fish school characteristics due to a tidal turbine support structure. Renew. Energy 2019, 141, 1092–1102. [Google Scholar] [CrossRef]
- Hastie, G.D.; Lepper, P.; McKnight, J.C.; Milne, R.; Russell, D.J.F.; Thompson, D. Acoustic risk balancing by marine mammals: Anthropogenic noise can influence the foraging decisions by seals. J. Appl. Ecol. 2021, 58, 1854–1863. [Google Scholar] [CrossRef]
- Smith, K.; Simpson, N. Enabling Future Arrays in Tidal: Y1 Environmental Monitoring Report. EnFiat. 2018. Available online: https://tethys.pnnl.gov/sites/default/files/publications/Y1_Environmental_Monitoring_Report.pdf (accessed on 22 July 2021).
- Hastie, G.D.; Russell, D.J.; Lepper, P.; Elliott, J.; Wilson, B.; Benjamins, S.; Thompson, D. Harbour seals avoid tidal turbine noise: Implications for collision risk. J. Appl. Ecol. 2017, 55, 684–693. [Google Scholar] [CrossRef]
- Polagye, B.; Bassett, C. Risk to Marine Animals from Underwater Noise Generated by Marine Renewable Energy Devices. In OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World; Copping, A.E., Hemery, L.G., Eds.; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 67–86. [Google Scholar] [CrossRef]
- Band, B.; Sparling, C.; Thompson, D.; Onoufriou, J.; San Martin, E.; West, N. Refining Estimates of Collision Risk for Harbour Seals and Tidal Turbines; Report for Scottish Government; ABP Marine Environmental Research Ltd. (ABPmer); Marine Scotland Science: Edinburgh, UK, 2016; pp. 1–133. [Google Scholar]
- Copping, A.; Grear, M.; Jepsen, R.; Chartrand, C.; Gorton, A. Understanding the potential risk to marine mammals from collision with tidal turbines. Int. J. Mar. Energy 2017, 19, 110–123. [Google Scholar] [CrossRef]
- Thompson, D.; Onoufriou, J.; Brownlow, A.; Morris, C. Data Based Estimates of Collision Risk: An Example Based on Harbour Seal Tracking Data around a Proposed Tidal Turbine Array in the Pentland Firth; Natural Scottish Heritage: Perth, UK, 2016; pp. 1–41. [Google Scholar]
- Thompson, D.; Brownlow, A.; Onoufriou, J.; Moss, S. Collision Risk and Impact Study: Field Tests of Turbine Blade-Seal Carcass Collisions; Report to Scottish Government MR 7.2.3; Marine Scotland Science: Edinburgh, UK, 2015; pp. 1–16. [Google Scholar]
- Onoufriou, J.; Brownlow, A.; Moss, S.; Hastie, G.; Thompson, D. Empirical determination of severe trauma in seals from collisions with tidal turbine blades. J. Appl. Ecol. 2019, 56, 1712–1724. [Google Scholar] [CrossRef]
- Boudreau, S.A.; Worm, B. Ecological role of large benthic decapods in marine ecosystems: A review. Mar. Ecol. Prog. Ser. 2012, 469, 195–213. [Google Scholar] [CrossRef] [Green Version]
- Pine, M. Underwater Anthropogenic Sound: Understanding the Potential Impacts on the Marine Environment and the Influence on Crab Larval Behaviour. Ph.D. Thesis, The University of Auckland, Auckland, New Zealand, 2013; pp. 1–152. [Google Scholar]
- Pine, M.K.; Jeffs, A.G.; Radford, C.A. Turbine Sound May Influence the Metamorphosis Behaviour of Estuarine Crab Megalopae. PLoS ONE 2012, 7, e51790. [Google Scholar] [CrossRef] [Green Version]
- Pine, M.K.; Jeffs, A.G.; Radford, C.A. Effects of Underwater Turbine Noise on Crab Larval Metamorphosis. In The Effects of Noise on Aquatic Life II; Popper, A.N., Hawkins, A., Eds.; Springer: New York, NY, USA, 2016; pp. 847–852. [Google Scholar]
- Taormina, B.; Di Poi, C.; Agnalt, A.-L.; Carlier, A.; Desroy, N.; Escobar-Lux, R.H.; D’eu, J.-F.; Freytet, F.; Durif, C.M.F. Impact of magnetic fields generated by AC/DC submarine power cables on the behavior of juvenile European lobster (Homarus gammarus). Aquat. Toxicol. 2020, 220, 105401. [Google Scholar] [CrossRef]
- Langhamer, O. The location of offshore wave power devices structures epifaunal assemblages. Int. J. Mar. Energy 2016, 16, 174–180. [Google Scholar] [CrossRef]
- Bicknell, A.W.J.; Sheehan, E.V.; Godley, B.J.; Doherty, P.D.; Witt, M.J. Assessing the impact of introduced infrastructure at sea with cameras: A case study for spatial scale, time and statistical power. Mar. Environ. Res. 2019, 147, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Langhamer, O.; Wilhelmsson, D. Colonisation of fish and crabs of wave energy foundations and the effects of manufactured holes—A field experiment. Mar. Environ. Res. 2009, 68, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Taormina, B.; Laurans, M.; Marzloff, M.P.; Dufournaud, N.; Lejart, M.; Desroy, N.; Leroy, D.; Martin, S.; Carlier, A. Renewable energy homes for marine life: Habitat potential of a tidal energy project for benthic megafauna. Mar. Environ. Res. 2020, 161, 105131. [Google Scholar] [CrossRef]
- Scott, K.; Harsanyi, P.; Lyndon, A.R. Understanding the effects of electromagnetic field emissions from Marine Renewable Energy Devices (MREDs) on the commercially important edible crab, Cancer pagurus (L.). Mar. Pollut. Bull. 2018, 131, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Causon, P.D.; Gill, A.B. Linking ecosystem services with epibenthic biodiversity change following installation of offshore wind farms. Environ. Sci. Policy 2018, 89, 340–347. [Google Scholar] [CrossRef]
- Wale, M.A.; Simpson, S.D.; Radford, A.N. Noise negatively affects foraging and antipredator behaviour in shore crabs. Anim. Behav. 2013, 86, 111–118. [Google Scholar] [CrossRef]
- Hutchison, Z.L.; Gill, A.B.; Sigray, P.; He, H.; King, J.W. Anthropogenic electromagnetic fields (EMF) influence the behaviour of bottom-dwelling marine species. Sci. Rep. 2020, 10, 4219. [Google Scholar] [CrossRef] [Green Version]
- Leitão, F.; Santos, M.N.; Erzini, K.; Monteiro, C.C. The effect of predation on artificial reef juvenile demersal fish species. Mar. Biol. 2008, 153, 1233–1244. [Google Scholar] [CrossRef]
- Woodruff, D.; Cullinan, V.; Copping, A.; Marshall, K. Effects of Electromagnetic Fields on Fish and Invertebrates—FY2012 Progress Report (Report No. PNNL-22154); Report for US Department of Energy (DOE); Pacific Northwest National Laboratory (PNNL): Richland, WA, USA, 2013; pp. 1–62. [Google Scholar]
- Bochert, R.; Zettler, M.L. Long-term exposure of several marine benthic animals to static magnetic fields. Bioelectromagnetics 2004, 25, 498–502. [Google Scholar] [CrossRef]
- Appel, M.; Elwood, R.W. Motivational trade-offs and potential pain experience in hermit crabs. Appl. Anim. Behav. Sci. 2009, 119, 120–124. [Google Scholar] [CrossRef]
- Keenan, G.; Sparling, C.; Williams, H.; Fortune, F. SeaGen Environmental Monitoring Programme: Final Report; Royal Haskoning Enhancing Society: Edinburgh, UK, 2011; p. 81. [Google Scholar]
- Haskoning, R. 2012 Environmental Monitoring Report Final Draft; Report for Marine Current Turbines (MCT); Ocean Renewable Power Company (ORPC): Portland, ME, USA, 2013; p. 653. [Google Scholar]
- National Wildlife Federation. Harbor Porpoise. Available online: https://www.nwf.org/Home/Educational-Resources/Wildlife-Guide/Mammals/Harbor-Porpoise (accessed on 13 October 2021).
- Mikkelsen, L.; Mouritsen, K.N.; Dahl, K.; Teilmann, J.; Tougaard, J. Re-established stony reef attracts harbour porpoises Phocoena phocoena. Mar. Ecol. Prog. Ser. 2013, 481, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Lossent, J.; Lejart, M.; Folegot, T.; Clorennec, D.; Di Iorio, L.; Gervaise, C. Underwater operational noise level emitted by a tidal current turbine and its potential impact on marine fauna. Mar. Pollut. Bull. 2018, 131, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Oregon State University. Final License Application for the PacWave South Project; Oregon State University: Corvallis, OR, USA, 2019; p. 1681. [Google Scholar]
- Robertson, F.; Wood, J.; Joslin, J.; Joy, R.; Polagye, B. Marine Mammal Behavioral Response to Tidal Turbine Sound; University of Washington: Seattle, WA, USA, 2018; pp. 1–65. [Google Scholar]
- Tollit, D.; Joy, R.; Wood, J.; Redden, A.; Booth, C.; Boucher, T.; Porskamp, P.; Oldreive, M. Baseline Presence of and Effects of Tidal Turbine Installation and Operations on Harbour Porpoise in Minas Passage, Bay of Fundy, Canada. J. Ocean Technol. 2019, 14, 22–48. [Google Scholar]
- Roberts, L.; Harding, H.R.; Voellmy, I.; Bruintjes, R.; Simpson, S.D.; Radford, A.N.; Breithaupt, T.; Elliott, M. Exposure of benthic invertebrates to sediment vibration: From laboratory experiments to outdoor simulated pile-driving. Proc. Meet. Acoust. 2016, 27, 010029. [Google Scholar] [CrossRef] [Green Version]
- Staines, G.; Zydlewski, G.B.; Viehman, H.A.; Kocik, R. Applying Two Active Acoustic Technologies to Document Presence of Large Marine Animal Targets at a Marine Renewable Energy Site. J. Mar. Sci. Eng. 2020, 8, 704. [Google Scholar] [CrossRef]
- Bassett, C. Ambient Noise in an Urbanized Tidal Channel. Ph.D. Thesis, University of Washington, Seattle, WA, USA, 2013. Available online: https://ui.adsabs.harvard.edu/abs/2013PhDT.......269B/abstract (accessed on 14 June 2021).
- Cruz, E.; Simas, T. Discussion of the Effects of the Underwater Noise Radiated by a Wave Energy Device-Portugal. In Proceedings of the 11th European Wave and Tidal Energy Conference, Nantes, France, 6–11 September 2015; pp. 1–5. [Google Scholar]
- Palmer, L.; Gillespie, D.; Macaulay, J.; Onoufriou, J.; Sparling, C.; Thompson, D.; Hastie, G. Marine Mammals and Tidal Energy: Annual Report to Scottish Government-MRE Theme; Sea Mammal Research Unit, University of St. Andrews: St Andrews, UK, 2019; pp. 1–32. [Google Scholar]
- Gill, A.B.; Desender, M. Risk to Animals from Electromagnetic Fields Emitted by Electric Cables and Marine Renewable Energy Devices. In OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World; Copping, A.E., Hemery, L.G., Eds.; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 86–103. [Google Scholar] [CrossRef]
- Castro-Santos, T.; Haro, A. Survival and Behavioral Effects of Exposure to a Hydrokinetic Turbine on Juvenile Atlantic Salmon and Adult American Shad. Estuaries Coasts 2015, 38, 203–214. [Google Scholar] [CrossRef]
- Bevelhimer, M.S.; Pracheil, B.M.; Fortner, A.M.; Saylor, R.; Deck, K.L. Mortality and injury assessment for three species of fish exposed to simulated turbine blade strike. Can. J. Fish. Aquat. Sci. 2019, 76, 2350–2363. [Google Scholar] [CrossRef]
- Xodus Group. Brims Tidal Array Collision Risk Modelling—Atlantic Salmon; Report No. A-100242-S02-TECH-001; Xodus Group: London, UK, 2016; pp. 1–11. [Google Scholar]
- Snyder, D.; Bailey, W.; Palmquist, K.; Cotts, B.; Olsen, K. Evaluation of Potential EMF Effects on Fish Species of Commercial or Recreational Fishing Importance in Southern New England; U.S. Department of the Interior, Bureau of Ocean Energy Management: Sterling, VA, USA, 2019; p. 62. [Google Scholar]
- Wyman, M.T.; Klimley, A.P.; Battleson, R.D.; Agosta, T.V.; Chapman, E.D.; Haverkamp, P.J.; Pagel, M.D.; Kavet, R. Behavioral responses by migrating juvenile salmonids to a subsea high-voltage DC power cable. Mar. Biol. 2018, 165, 134. [Google Scholar] [CrossRef]
- Kramer, S.H.; Hamilton, C.D.; Spencer, G.C.; Ogston, H.O. Evaluating the Potential for Marine and Hydrokinetic Devices to Act as Artificial Reefs or Fish Aggregating Devices. Based on Analysis of Surrogates in Tropical, Subtropical, and Temperate U.S. West Coast and Hawaiian Coastal Waters; U.S. Department of Energy Efficiency and Renewable Energy: Golden, CO, USA, 2015; p. 90. [Google Scholar] [CrossRef]
- Bevelhimer, M.S.; Deng, Z.D.; Scherelis, C. Characterizing large river sounds: Providing context for understanding the environmental effects of noise produced by hydrokinetic turbines. J. Acoust. Soc. Am. 2016, 139, 85–92. [Google Scholar] [CrossRef]
- Halvorsen, M.B.; Carlson, T.J.; Copping, A.E. Effects of Tidal Turbine Noise on Fish Hearing and Tissues—Draft Final Report—Environmental Effects of Marine and Hydrokinetic Energy; Pacific Northwest National Lab. (PNNL-20786): Richland, WA, USA, 2011; pp. 1–48. [Google Scholar]
- Guerra, M.; Thomson, J. Wake measurements from a hydrokinetic river turbine. Renew. Energy 2019, 139, 483–495. [Google Scholar] [CrossRef]
- Camperi, M.; Tricas, T.C.; Brown, B.R. From Morphology to Neural Information: The Electric Sense of the Skate. PLoS Comput. Biol. 2007, 3, e113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fundy Ocean Research Center for Energy (FORCE). Environmental Effects Monitoring Report 2011–2013; FORCE: Parrsboro, NS, Canada, 2014; pp. 1–44. [Google Scholar]
- Hutchison, Z.; Sigray, P.; He, H.; Gill, A.; Gibson, C. Electromagnetic Field (EMF) Impacts on Elasmobranch (Shark, Rays, and Skates) and American Lobster Movement and Migration from Direct Current Cables; U.S. Department of the Interior, Bureau of Ocean Energy Management: Sterling, VA, USA, 2018; pp. 1–254. [Google Scholar]
- Henkel, S. Assessment of Benthic Effects of Anchor Presence and Removal; Report for Oregon Wave Energy Trust (OWET); Northwest National Marine Renewable Energy Center (NNMREC): Corvallis, OR, USA, 2016; pp. 1–22. [Google Scholar]
- Kregting, L.; Elsaesser, B.; Kennedy, R.; Smyth, D.; O’Carroll, J.; Savidge, G. Do Changes in Current Flow as a Result of Arrays of Tidal Turbines Have an Effect on Benthic Communities? PLoS ONE 2016, 11, e0161279. [Google Scholar] [CrossRef] [Green Version]
- Kuhnz, L.; Barry, J.; Buck, B.; Whaling, P. Potential Impacts of the Monterey Accelerated Research System (MARS) Cable on the Seabed and Benthic Faunal Assemblages; Monterey Bay Aquarium Research Institute: Moss Landing, CA, USA, 2011; pp. 1–58. [Google Scholar]
- Packer, D.; Zetlin, C.; Vitaliano, J. Essential Fish and Habitat Source Document: Thorny Skate, Amblyraja Radiata, Life History and Habitat Characteristics; U.S. Department of Commerce National Oceanic and Atmospheric Administration: Woods Hole, MA, USA, 2003; pp. 1–50. [Google Scholar]
- Hastie, G.; Lepper, P.; McKnight, C.; Milne, R.; Russell, D.; Thompson, D. Risk Balancing by a Marine Mammal: Man-Made Noise Reduces the Foraging Success of Seals [Abstract]. In Proceedings of the World Marine Mammal Conference (WMMC2019), Barcelona, Spain, 9–12 December 2019; pp. 1–2. [Google Scholar]
- Sparling, C.E.; Seitz, A.C.; Masden, E.; Smith, K. Collision Risk for Animals around Turbines. In OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World; Copping, A.E., Hemery, L.G., Eds.; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 29–65. [Google Scholar] [CrossRef]
- RPS Group. A Review of the Potential Use of Sonar to Observe the Underwater Behaviour of Diving Birds near Tidal Energy Devices (Report No. Report 051); Report for Scottish Natural Heritage; RPS Group: Chepstow, UK, 2010; pp. 1–26. [Google Scholar]
- Lieber, L.; Nimmo-Smith, W.A.M.; Waggitt, J.J.; Kregting, L. Localised anthropogenic wake generates a predictable foraging hotspot for top predators. Commun. Biol. 2019, 2, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCluskie, A.; Langston, A.; Wilkinson, N. Birds and Wave & Tidal Stream Energy: An Ecological Review; The Royal Society for the Protection of Birds (RSPB): Sandy, UK, 2012; p. 124. [Google Scholar]
- Gibbs, M.T.; Browman, H.I. Risk assessment and risk management: A primer for marine scientists. ICES J. Mar. Sci. 2015, 72, 992–996. [Google Scholar] [CrossRef] [Green Version]
- Holt, E.; Miller, S. Bioindicators: Using Organisms to Measure Environmental Impacts. Nat. Educ. Knowl. 2011, 3, 8. [Google Scholar]
- Bender, A.; Langhamer, O.; Sundberg, J. Colonisation of wave power foundations by mobile mega- and macrofauna—A 12 year study. Mar. Environ. Res. 2020, 161, 105053. [Google Scholar] [CrossRef]
- Langhamer, O. Artificial Reef Effect in Relation to Offshore Renewable Energy Conversion: State of the Art. Sci. World J. 2012, 2012, 386713. [Google Scholar] [CrossRef]
- Wilhelmsson, D.; Langhamer, O. The Influence of Fisheries Exclusion and Addition of Hard Substrata on Fish and Crustaceans. In Humanity and the Sea: Marine Renewable Energy Technology and Environmental Interactions; Springer: Berlin/Heidelberg, Germany, 2014; pp. 49–60. [Google Scholar]
- Copping, A.E.; Grear, M.E. Applying a simple model for estimating the likelihood of collision of marine mammals with tidal turbines. Int. Mar. Energy J. 2018, 1, 27–33. [Google Scholar] [CrossRef]
- Whiting, J.M.; Chang, G. Changes in Oceanographic Systems Associated with Marine Renewable Energy Devices. In OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World; Copping, A.E., Hemery, L.G., Eds.; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 127–145. [Google Scholar] [CrossRef]
- Wang, T.; Yang, Z.; Copping, A. A Modeling Study of the Potential Water Quality Impacts from In-Stream Tidal Energy Extraction. Estuaries Coasts 2015, 38, 173–186. [Google Scholar] [CrossRef]
- Nagelkerken, I.; Goldenberg, S.U.; Ferreira, C.M.; Ullah, H.; Connell, S.D. Trophic pyramids reorganize when food web architecture fails to adjust to ocean change. Science 2020, 369, 829–832. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hemery, L.G.; Copping, A.E.; Overhus, D.M. Biological Consequences of Marine Energy Development on Marine Animals. Energies 2021, 14, 8460. https://doi.org/10.3390/en14248460
Hemery LG, Copping AE, Overhus DM. Biological Consequences of Marine Energy Development on Marine Animals. Energies. 2021; 14(24):8460. https://doi.org/10.3390/en14248460
Chicago/Turabian StyleHemery, Lenaïg G., Andrea E. Copping, and Dorian M. Overhus. 2021. "Biological Consequences of Marine Energy Development on Marine Animals" Energies 14, no. 24: 8460. https://doi.org/10.3390/en14248460
APA StyleHemery, L. G., Copping, A. E., & Overhus, D. M. (2021). Biological Consequences of Marine Energy Development on Marine Animals. Energies, 14(24), 8460. https://doi.org/10.3390/en14248460