The Components of Bone and What They Can Teach Us about Regeneration

 ,
,
Abstract
:1. Introduction
2. Dissecting Bone at the Tissue Level
2.1. Periosteum
2.2. Osseous Tissue
2.3. Endosteum
2.4. Bone Marrow
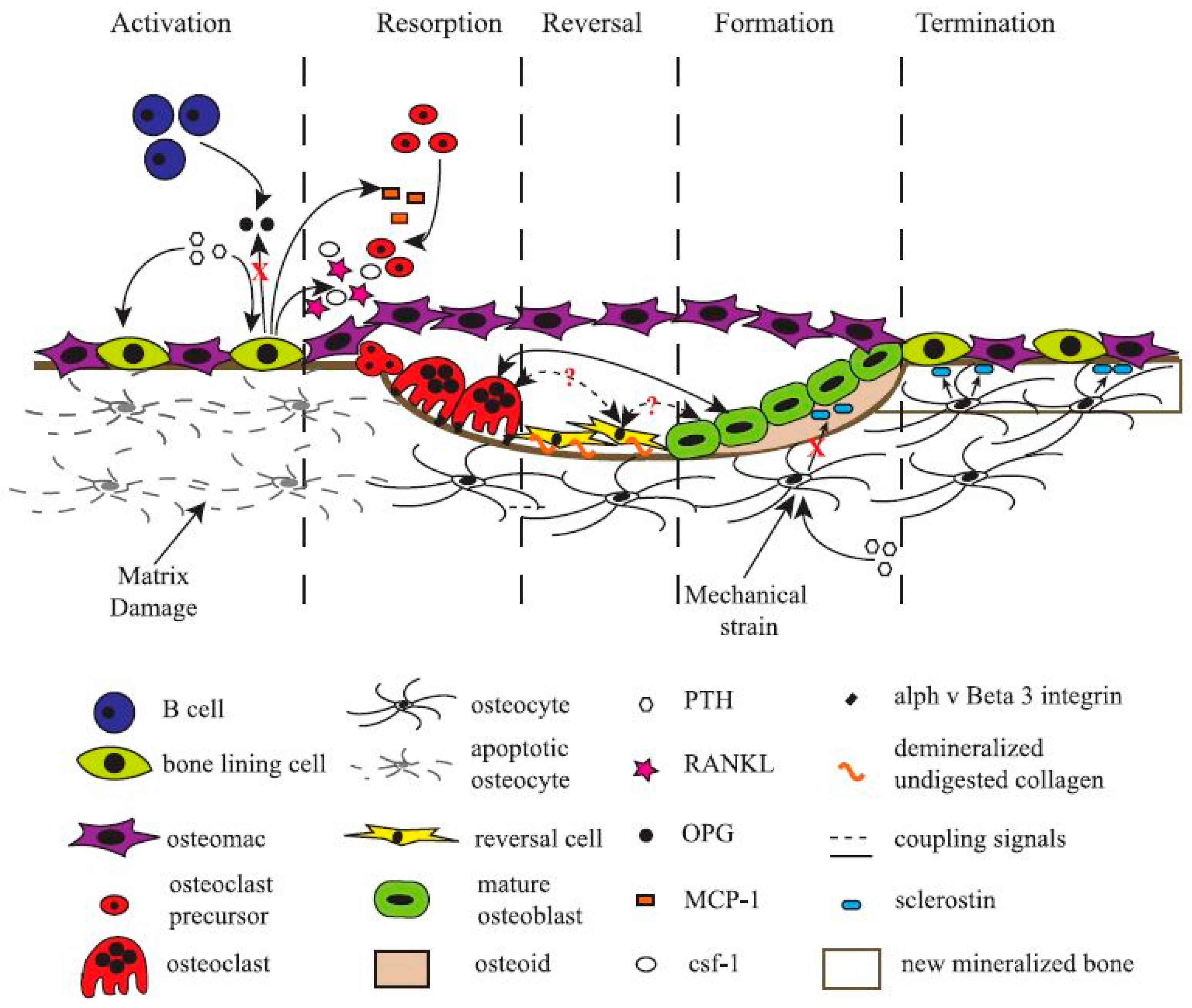
3. The Cells in Bone
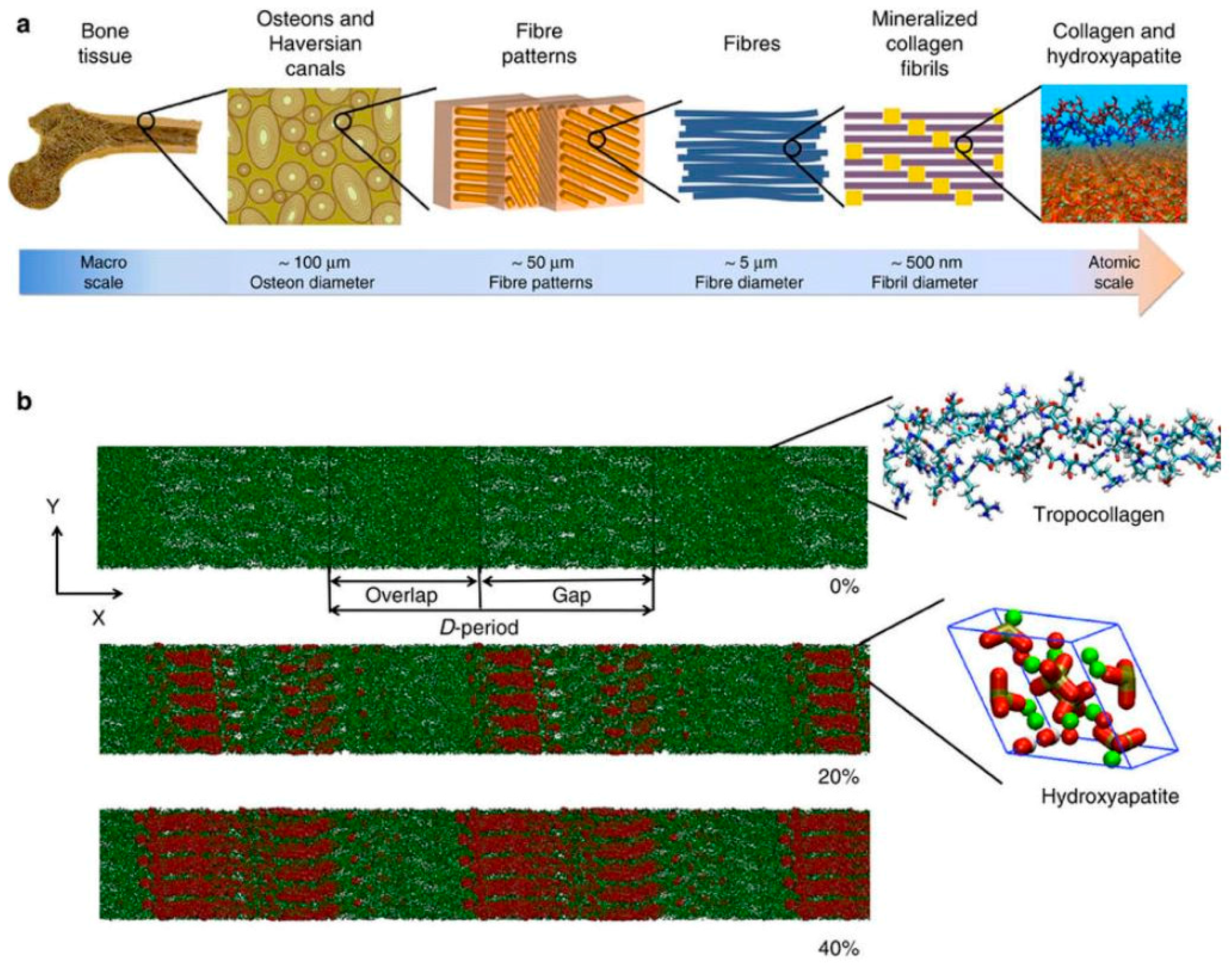
4. The Extracellular Matrix of Bone
5. The Future of Bone Tissue Engineering
Acknowledgments
Conflicts of Interest
References
- Clarke, B. Normal bone anatomy and physiology. Clin. J. Am. Soc. Nephrol. 2008, 3 (Suppl. 3), S131–S139. [Google Scholar] [CrossRef] [PubMed]
- Burt, L.A.; Greene, D.A.; Naughton, G.A. Bone health of young male gymnasts: A systematic review. Pediatr. Exerc. Sci. 2017, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Victoria, G.; Petrisor, B.; Drew, B.; Dick, D. Bone stimulation for fracture healing: What’s all the fuss? Indian J. Orthop. 2009, 43, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Global Burden of Disease Study 2013 Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 301 acute and chronic diseases and injuries in 188 countries, 1990–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2015, 386, 743–800. [Google Scholar] [CrossRef]
- Urist, M.R. Bone: Formation by autoinduction. Science 1965, 150, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Cranford, S.W.; de Boer, J.; van Blitterswijk, C.; Buehler, M.J. Materiomics: An-omics approach to biomaterials research. Adv. Mater. 2013, 25, 802–824. [Google Scholar] [CrossRef] [PubMed]
- Ollier, L. Recherches experimentales sur les greffes osseuses. impr. Jules Claye. J. Physiol. Homme Animaux 1860, 3, 88. [Google Scholar]
- Duhamel, H. Sur le developpement et la crue des os des animaux. Mem. Acad. R. Sci. 1742, 55, 354–357. [Google Scholar]
- Guillot, D. Almost invisible, often ignored: Periosteum, the living lace of bone. Medicographia 2012, 34, 221–227. [Google Scholar]
- OpenStar_College. “File:603-Anatomy of Long Bone”; “File:606-Spongy Bone”; “File:624-Diagram of Compact Bone”. Available online: https://commons.wikimedia.org/wiki/ (accessed on 22 December 2017).
- Doi, K.; Sakai, K. Vascularized periosteal bone graft from the supracondylar region of the femur. Microsurgery 1994, 15, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Vogelin, E.; Jones, N.F.; Huang, J.I.; Brekke, J.H.; Lieberman, J.R. Healing of a critical-sized defect in the rat femur with use of a vascularized periosteal flap, a biodegradable matrix, and bone morphogenetic protein. J. Bone Jt. Surg. Am. 2005, 87, 1323–1331. [Google Scholar] [CrossRef]
- Del Pinal, F.; Garcia-Bernal, F.J.; Regalado, J.; Ayala, H.; Cagigal, L.; Studer, A. Vascularised corticoperiosteal grafts from the medial femoral condyle for difficult non-unions of the upper limb. J. Hand Surg. Eur. Vol. 2007, 32, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Steinmann, S.P.; Bishop, A.T. Free vascularized corticoperiosteal bone graft for the treatment of persistent nonunion of the clavicle. J. Shoulder Elb. Surg. 2005, 14, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Shimizu, H.; Beppu, M.; Takagi, M. The role of beta-tricalcium phosphate in vascularized periosteum. J. Orthop. Sci. 2000, 5, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Owen, M. The origin of bone cells. Int. Rev. Cytol. 1970, 28, 213–238. [Google Scholar] [PubMed]
- Nakahara, H.; Dennis, J.E.; Bruder, S.P.; Haynesworth, S.E.; Lennon, D.P.; Caplan, A.I. In vitro differentiation of bone and hypertrophic cartilage from periosteal-derived cells. Exp. Cell Res. 1991, 195, 492–503. [Google Scholar] [CrossRef]
- Kisiel, A.H.; McDuffee, L.A.; Masaoud, E.; Bailey, T.R.; Esparza Gonzalez, B.P.; Nino-Fong, R. Isolation, characterization, and in vitro proliferation of canine mesenchymal stem cells derived from bone marrow, adipose tissue, muscle, and periosteum. Am. J. Vet. Res. 2012, 73, 1305–1317. [Google Scholar] [CrossRef] [PubMed]
- Colnot, C. Skeletal cell fate decisions within periosteum and bone marrow during bone regeneration. J. Bone Miner. Res. 2009, 24, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Bilkay, U.; Tokat, C.; Helvaci, E.; Ozek, C.; Zekioglu, O.; Onat, T.; Songur, E. Osteogenic capacities of tibial and cranial periosteum: A biochemical and histologic study. J. Craniofac. Surg. 2008, 19, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Uddstromer, L. The osteogenic capacity of tubular and membranous bone periosteum. A qualitative and quantitative experimental study in growing rabbits. Scand. J. Plast. Reconstr. Surg. 1978, 12, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Schonmeyr, B.; Clavin, N.; Avraham, T.; Longo, V.; Mehrara, B.J. Synthesis of a tissue-engineered periosteum with acellular dermal matrix and cultured mesenchymal stem cells. Tissue Eng. Part A 2009, 15, 1833–1841. [Google Scholar] [CrossRef] [PubMed]
- Beniker, D.; McQuillan, D.; Livesey, S.; Urban, R.M.; Turner, T.M.; Blum, B.; Hughes, K.; Haggard, W.O. The use of acellular dermal matrix as a scaffold for periosteum replacement. Orthopedics 2003, 26, S591–S596. [Google Scholar] [PubMed]
- Hattori, K.; Yoshikawa, T.; Takakura, Y.; Aoki, H.; Sonobe, M.; Tomita, N. Bio-artificial periosteum for severe open fracture—An experimental study of osteogenic cell/collagen sponge composite as a bio-artificial periosteum. Biomed. Mater. Eng. 2005, 15, 127–136. [Google Scholar] [PubMed]
- Zhang, K.G.; Zeng, B.F.; Zhang, C.Q. Periosteum construction in vitro by small intestinal submucosa combined with bone marrow mesenchymal stem cell. Chin. J. Surg. 2005, 43, 1594–1597. [Google Scholar] [PubMed]
- Baldwin, J.G.; Wagner, F.; Martine, L.C.; Holzapfel, B.M.; Theodoropoulos, C.; Bas, O.; Savi, F.M.; Werner, C.; De-Juan-Pardo, E.M.; Hutmacher, D.W. Periosteum tissue engineering in an orthotopic in vivo platform. Biomaterials 2017, 121, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Aarden, E.M.; Burger, E.H.; Nijweide, P.J. Function of osteocytes in bone. J. Cell. Biochem. 1994, 55, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, Y.; Sekiya, I.; Yagishita, K.; Ichinose, S.; Shinomiya, K.; Muneta, T. Suspended cells from trabecular bone by collagenase digestion become virtually identical to mesenchymal stem cells obtained from marrow aspirates. Blood 2004, 104, 2728–2735. [Google Scholar] [CrossRef] [PubMed]
- Tuli, R.; Seghatoleslami, M.R.; Tuli, S.; Wang, M.L.; Hozack, W.J.; Manner, P.A.; Danielson, K.G.; Tuan, R.S. A simple, high-yield method for obtaining multipotential mesenchymal progenitor cells from trabecular bone. Mol. Biotechnol. 2003, 23, 37–49. [Google Scholar] [CrossRef]
- Keaveny, T.M.; Hayes, W.C. Mechanical properties of cortical and trabecular bone. Bone 1993, 7, 285–344. [Google Scholar]
- Pape, H.C.; Evans, A.; Kobbe, P. Autologous bone graft: Properties and techniques. J. Orthop. Trauma 2010, 24 (Suppl. S1), S36–S40. [Google Scholar] [CrossRef] [PubMed]
- Albrektsson, T.; Johansson, C. Osteoinduction, osteoconduction and osseointegration. Eur. Spine J. 2001, 10 (Suppl. S2), S96–S101. [Google Scholar] [CrossRef] [PubMed]
- Burchardt, H. Biology of bone transplantation. Orthop. Clin. N. Am. 1987, 18, 187–196. [Google Scholar]
- Khan, S.N.; Cammisa, F.P., Jr.; Sandhu, H.S.; Diwan, A.D.; Girardi, F.P.; Lane, J.M. The biology of bone grafting. J. Am. Acad. Orthop. Surg. 2005, 13, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Minami, A.; Kasashima, T.; Iwasaki, N.; Kato, H.; Kaneda, K. Vascularised fibular grafts. An experience of 102 patients. J. Bone Jt. Surg. Br. 2000, 82, 1022–1025. [Google Scholar] [CrossRef]
- Roberts, T.T.; Rosenbaum, A.J. Bone grafts, bone substitutes and orthobiologics: The bridge between basic science and clinical advancements in fracture healing. Organogenesis 2012, 8, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Raggatt, L.J.; Partridge, N.C. Cellular and molecular mechanisms of bone remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [PubMed]
- Aldridge, J.M., 3rd; Urbaniak, J.R. Avascular necrosis of the femoral head: Role of vascularized bone grafts. Orthop.Clin. N. Am. 2007, 38, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Hauschka, P.V.; Mavrakos, A.E.; Iafrati, M.D.; Doleman, S.E.; Klagsbrun, M. Growth factors in bone matrix. Isolation of multiple types by affinity chromatography on heparin-Sepharose. J. Biol. Chem. 1986, 261, 12665–12674. [Google Scholar] [PubMed]
- Blum, B.; Moseley, J.; Miller, L.; Richelsoph, K.; Haggard, W. Measurement of bone morphogenetic proteins and other growth factors in demineralized bone matrix. Orthopedics 2004, 27, S161–S165. [Google Scholar] [PubMed]
- Wildemann, B.; Kadow-Romacker, A.; Haas, N.P.; Schmidmaier, G. Quantification of various growth factors in different demineralized bone matrix preparations. J. Biomed. Mater. Res. A 2007, 81, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Drosos, G.I.; Touzopoulos, P.; Ververidis, A.; Tilkeridis, K.; Kazakos, K. Use of demineralized bone matrix in the extremities. World J. Orthop. 2015, 6, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Zadegan, S.A.; Abedi, A.; Jazayeri, S.B.; Vaccaro, A.R.; Rahimi-Movaghar, V. Demineralized bone matrix in anterior cervical discectomy and fusion: A systematic review. Eur. Spine J. 2017, 26, 958–974. [Google Scholar] [CrossRef] [PubMed]
- Gerard, J.; Tortora, B.H.D. The skeletal system: Bone tissue. In Principles of Anatomy and Physiology, 13rd ed.; Wiley: Hoboken, NJ, USA, 2012; pp. 182–207. [Google Scholar]
- McParland, B.J. Nuclear Medicine Radiation Dosimetry; Springer: Dordrecht, The Netherlands, 2010. [Google Scholar]
- Luk, S.C.; Nopajaroonsri, C.; Simon, G.T. The ultrastructure of endosteum: A topographic study in young adult rabbits. J. Ultrastruct. Res. 1974, 46, 165–183. [Google Scholar] [CrossRef]
- Schepers, K.; Pietras, E.M.; Reynaud, D.; Flach, J.; Binnewies, M.; Garg, T.; Wagers, A.J.; Hsiao, E.C.; Passegue, E. Myeloproliferative neoplasia remodels the endosteal bone marrow niche into a self-reinforcing leukemic niche. Cell Stem Cell 2013, 13, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 505, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Bianco, P.; Robey, P.G. Skeletal stem cells. Development 2015, 142, 1023–1027. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I.; Correa, D. The MSC: An injury drugstore. Cell Stem Cell 2011, 9, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Tavassoli, M.; Crosby, W.H. Transplantation of marrow to extramedullary sites. Science 1968, 161, 54–56. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Piatetzky, S., II; Petrakova, K.V. Osteogenesis in transplants of bone marrow cells. J. Embryol. Exp. Morphol. 1966, 16, 381–390. [Google Scholar] [PubMed]
- Kuznetsov, S.A.; Krebsbach, P.H.; Satomura, K.; Kerr, J.; Riminucci, M.; Benayahu, D.; Robey, P.G. Single-colony derived strains of human marrow stromal fibroblasts form bone after transplantation in vivo. J. Bone Miner. Res. 1997, 12, 1335–1347. [Google Scholar] [CrossRef] [PubMed]
- Gianakos, A.; Ni, A.; Zambrana, L.; Kennedy, J.G.; Lane, J.M. Bone Marrow Aspirate Concentrate in Animal Long Bone Healing: An Analysis of Basic Science Evidence. J. Orthop. Trauma 2016, 30, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Barrena, E.; Rosset, P.; Lozano, D.; Stanovici, J.; Ermthaller, C.; Gerbhard, F. Bone fracture healing: Cell therapy in delayed unions and nonunions. Bone 2015, 70, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Le Nail, L.R.; Stanovici, J.; Fournier, J.; Splingard, M.; Domenech, J.; Rosset, P. Percutaneous grafting with bone marrow autologous concentrate for open tibia fractures: Analysis of forty three cases and literature review. Int. Orthop. 2014, 38, 1845–1853. [Google Scholar] [CrossRef] [PubMed]
- Bortolotti, F.; Ukovich, L.; Razban, V.; Martinelli, V.; Ruozi, G.; Pelos, B.; Dore, F.; Giacca, M.; Zacchigna, S. In vivo therapeutic potential of mesenchymal stromal cells depends on the source and the isolation procedure. Stem Cell Rep. 2015, 4, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Corral, D.A.; Amling, M.; Priemel, M.; Loyer, E.; Fuchs, S.; Ducy, P.; Baron, R.; Karsenty, G. Dissociation between bone resorption and bone formation in osteopenic transgenic mice. Proc. Natl. Acad. Sci. USA 1998, 95, 13835–13840. [Google Scholar] [CrossRef] [PubMed]
- Long, F. Building strong bones: Molecular regulation of the osteoblast lineage. Nat. Rev. Mol. Cell Biol. 2011, 13, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.N.; Hankenson, K.D. Mesenchymal Stem Cells in Bone Regeneration. Adv. Wound Care (New Rochelle) 2013, 2, 306–316. [Google Scholar] [CrossRef] [PubMed]
- da Silva Meirelles, L.; Caplan, A.I.; Nardi, N.B. In search of the in vivo identity of mesenchymal stem cells. Stem Cells 2008, 26, 2287–2299. [Google Scholar] [CrossRef] [PubMed]
- Murray, I.R.; Peault, B. Q&A: Mesenchymal stem cells—Where do they come from and is it important? BMC Biol. 2015, 13, 99. [Google Scholar] [CrossRef]
- Uccelli, A.; Moretta, L.; Pistoia, V. Mesenchymal stem cells in health and disease. Nat. Rev. Immunol. 2008, 8, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells-current trends and future prospective. Biosci. Rep. 2015, 35. [Google Scholar] [CrossRef] [PubMed]
- Lalu, M.M.; McIntyre, L.; Pugliese, C.; Fergusson, D.; Winston, B.W.; Marshall, J.C.; Granton, J.; Stewart, D.J.; Canadian Critical Care Trials, G. Safety of cell therapy with mesenchymal stromal cells (SafeCell): A systematic review and meta-analysis of clinical trials. PLoS ONE 2012, 7, e47559. [Google Scholar] [CrossRef] [PubMed]
- Goshima, J.; Goldberg, V.M.; Caplan, A.I. The osteogenic potential of culture-expanded rat marrow mesenchymal cells assayed in vivo in calcium phosphate ceramic blocks. Clin. Orthop. Relat. Res. 1991, 298–311. [Google Scholar] [CrossRef]
- Arinzeh, T.L.; Peter, S.J.; Archambault, M.P.; van den Bos, C.; Gordon, S.; Kraus, K.; Smith, A.; Kadiyala, S. Allogeneic mesenchymal stem cells regenerate bone in a critical-sized canine segmental defect. J. Bone Jt. Surg. Am. 2003, 85-A, 1927–1935. [Google Scholar] [CrossRef]
- Kruyt, M.C.; Wilson, C.E.; de Bruijn, J.D.; van Blitterswijk, C.A.; Oner, C.F.; Verbout, A.J.; Dhert, W.J. The effect of cell-based bone tissue engineering in a goat transverse process model. Biomaterials 2006, 27, 5099–5106. [Google Scholar] [CrossRef] [PubMed]
- Siddappa, R.; Martens, A.; Doorn, J.; Leusink, A.; Olivo, C.; Licht, R.; van Rijn, L.; Gaspar, C.; Fodde, R.; Janssen, F.; et al. cAMP/PKA pathway activation in human mesenchymal stem cells in vitro results in robust bone formation in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 7281–7286. [Google Scholar] [CrossRef] [PubMed]
- Chatterjea, A.; Meijer, G.; van Blitterswijk, C.; de Boer, J. Clinical application of human mesenchymal stromal cells for bone tissue engineering. Stem Cells Int. 2010, 2010, 215625. [Google Scholar] [CrossRef] [PubMed]
- Rosset, P.; Deschaseaux, F.; Layrolle, P. Cell therapy for bone repair. Orthop. Traumatol. Surg. Res. 2014, 100, S107–S112. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Gotherstrom, C.; Ringden, O.; Hassan, M.; McMahon, R.; Horwitz, E.; Anneren, G.; Axelsson, O.; Nunn, J.; Ewald, U.; et al. Fetal mesenchymal stem-cell engraftment in bone after in utero transplantation in a patient with severe osteogenesis imperfecta. Transplantation 2005, 79, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, E.M.; Gordon, P.L.; Koo, W.K.; Marx, J.C.; Neel, M.D.; McNall, R.Y.; Muul, L.; Hofmann, T. Isolated allogeneic bone marrow-derived mesenchymal cells engraft and stimulate growth in children with osteogenesis imperfecta: Implications for cell therapy of bone. Proc. Natl. Acad. Sci. USA 2002, 99, 8932–8937. [Google Scholar] [CrossRef] [PubMed]
- Miron, R.J.; Bosshardt, D.D. OsteoMacs: Key players around bone biomaterials. Biomaterials 2016, 82, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Batoon, L.; Millard, S.M.; Raggatt, L.J.; Pettit, A.R. Osteomacs and Bone Regeneration. Curr. Osteoporos. Rep. 2017, 15, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.C.; Raggatt, L.J.; Alexander, K.A.; Pettit, A.R. Unraveling macrophage contributions to bone repair. Bonekey Rep. 2013, 2, 373. [Google Scholar] [CrossRef] [PubMed]
- Burnett, S.H.; Kershen, E.J.; Zhang, J.; Zeng, L.; Straley, S.C.; Kaplan, A.M.; Cohen, D.A. Conditional macrophage ablation in transgenic mice expressing a Fas-based suicide gene. J. Leukoc. Biol. 2004, 75, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Alexander, K.A.; Chang, M.K.; Maylin, E.R.; Kohler, T.; Muller, R.; Wu, A.C.; Van Rooijen, N.; Sweet, M.J.; Hume, D.A.; Raggatt, L.J.; et al. Osteal macrophages promote in vivo intramembranous bone healing in a mouse tibial injury model. J. Bone Miner. Res. 2011, 26, 1517–1532. [Google Scholar] [CrossRef] [PubMed]
- Davison, N.L.; Gamblin, A.L.; Layrolle, P.; Yuan, H.; de Bruijn, J.D.; Barrere-de Groot, F. Liposomal clodronate inhibition of osteoclastogenesis and osteoinduction by submicrostructured beta-tricalcium phosphate. Biomaterials 2014, 35, 5088–5097. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.K.; Raggatt, L.J.; Alexander, K.A.; Kuliwaba, J.S.; Fazzalari, N.L.; Schroder, K.; Maylin, E.R.; Ripoll, V.M.; Hume, D.A.; Pettit, A.R. Osteal tissue macrophages are intercalated throughout human and mouse bone lining tissues and regulate osteoblast function in vitro and in vivo. J. Immunol. 2008, 181, 1232–1244. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, H.; Moroni, L.; van Blitterswijk, C.; de Boer, J. Extracellular matrix and tissue engineering applications. J. Mater. Chem. 2009, 19, 5474–5484. [Google Scholar] [CrossRef]
- Escobedo-Lucea, C.; Ayuso-Sacido, A.; Xiong, C.; Prado-Lopez, S.; del Pino, M.S.; Melguizo, D.; Bellver-Estelles, C.; Gonzalez-Granero, S.; Valero, M.L.; Moreno, R.; et al. Development of a human extracellular matrix for applications related with stem cells and tissue engineering. Stem Cell Rev. Rep. 2012, 8, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Mouw, J.K.; Ou, G.; Weaver, V.M. Extracellular matrix assembly: A multiscale deconstruction. Nat. Rev. Mol. Cell Biol. 2014, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Hadjidakis, D.J.; Androulakis, II. Bone remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Eppley, B.L.; Pietrzak, W.S.; Blanton, M.W. Allograft and alloplastic bone substitutes: A review of science and technology for the craniomaxillofacial surgeon. J. Craniofac. Surg. 2005, 16, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Gruskin, E.; Doll, B.A.; Futrell, F.W.; Schmitz, J.P.; Hollinger, J.O. Demineralized bone matrix in bone repair: History and use. Adv. Drug Deliv. Rev. 2012, 64, 1063–1077. [Google Scholar] [CrossRef] [PubMed]
- Kontis, T.C.; Rivkin, A. The history of injectable facial fillers. Facial Plast. Surg. 2009, 25, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Viguet-Carrin, S.; Garnero, P.; Delmas, P.D. The role of collagen in bone strength. Osteop. Int. 2006, 17, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Campana, V.; Milano, G.; Pagano, E.; Barba, M.; Cicione, C.; Salonna, G.; Lattanzi, W.; Logroscino, G. Bone substitutes in orthopaedic surgery: From basic science to clinical practice. J. Mater. Sci. Mater. Med. 2014, 25, 2445–2461. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.M.; Gentile, P.; Chiono, V.; Ciardelli, G. Collagen for bone tissue regeneration. Acta Biomater. 2012, 8, 3191–3200. [Google Scholar] [CrossRef] [PubMed]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [PubMed]
- Gorgieva, S.; Kokol, V. Collagen- vs. gelatine-based biomaterials and their biocompatibility: Review and perspectives. In Biomaterials Applications for Nanomedicine; InTech: Rijeka, Croatia, 2011. [Google Scholar]
- Chattopadhyay, S.; Raines, R.T. Review collagen-based biomaterials for wound healing. Biopolymers 2014, 101, 821–833. [Google Scholar] [CrossRef] [PubMed]
- O’brien, F.J. Biomaterials & scaffolds for tissue engineering. Mater. Today 2011, 14, 88–95. [Google Scholar]
- Sweeney, T.M.; Opperman, L.A.; Persing, J.A.; Ogle, R.C. Repair of critical size rat calvarial defects using extracellular matrix protein gels. J. Neurosurg. 1995, 83, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Saadeh, P.B.; Khosla, R.K.; Mehrara, B.J.; Steinbrech, D.S.; McCormick, S.A.; DeVore, D.P.; Longaker, M.T. Repair of a critical size defect in the rat mandible using allogenic type I collagen. J. Craniofac. Surg. 2001, 12, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Cicciu, M. Real Opportunity for the Present and a Forward Step for the Future of Bone Tissue Engineering. J. Craniofac. Surg. 2017, 28, 592–593. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.K.; Gautieri, A.; Chang, S.W.; Buehler, M.J. Molecular mechanics of mineralized collagen fibrils in bone. Nat. Commun. 2013, 4, 1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glimcher, M.J. Bone: Nature of the calcium phosphate crystals and cellular, structural, and physical chemical mechanisms in their formation. Rev. Mineral. Geochem. 2006, 64, 223–282. [Google Scholar] [CrossRef]
- Dorozhkin, S. History of Calcium Phosphates in Regenerative Medicine. In Advances in Calcium Phosphate Biomaterials; Springer: Berlin, Germany, 2014; pp. 435–483. [Google Scholar]
- Wang, P.; Zhao, L.; Liu, J.; Weir, M.D.; Zhou, X.; Xu, H.H. Bone tissue engineering via nanostructured calcium phosphate biomaterials and stem cells. Bone Res. 2014, 2, 14017. [Google Scholar] [CrossRef] [PubMed]
- den Hollander, W.; Patka, P.; Klein, C.P.; Heidendal, G.A. Macroporous calcium phosphate ceramics for bone substitution: A tracer study on biodegradation with 45Ca tracer. Biomaterials 1991, 12, 569–573. [Google Scholar] [CrossRef]
- Barrere, F.; van Blitterswijk, C.A.; de Groot, K. Bone regeneration: Molecular and cellular interactions with calcium phosphate ceramics. Int. J. Nanomed. 2006, 1, 317–332. [Google Scholar]
- Yuan, H.; Kurashina, K.; de Bruijn, J.D.; Li, Y.; de Groot, K.; Zhang, X. A preliminary study on osteoinduction of two kinds of calcium phosphate ceramics. Biomaterials 1999, 20, 1799–1806. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, T.; Zhu, J.; Cai, P. Osteoinduction of Calcium Phosphate Ceramics in Four Kinds of Animals for 1 Year: Dog, Rabbit, Rat, and Mouse. Transplant. Proc. 2016, 48, 1309–1314. [Google Scholar] [CrossRef] [PubMed]
- Barradas, A.M.; Fernandes, H.A.; Groen, N.; Chai, Y.C.; Schrooten, J.; van de Peppel, J.; van Leeuwen, J.P.; van Blitterswijk, C.A.; de Boer, J. A calcium-induced signaling cascade leading to osteogenic differentiation of human bone marrow-derived mesenchymal stromal cells. Biomaterials 2012, 33, 3205–3215. [Google Scholar] [CrossRef] [PubMed]
- Wennerberg, A.; Albrektsson, T. Effects of titanium surface topography on bone integration: A systematic review. Clin. Oral Implant. Res. 2009, 20 (Suppl. S4), 172–184. [Google Scholar] [CrossRef] [PubMed]
- Unadkat, H.V.; Hulsman, M.; Cornelissen, K.; Papenburg, B.J.; Truckenmuller, R.K.; Carpenter, A.E.; Wessling, M.; Post, G.F.; Uetz, M.; Reinders, M.J.; et al. An algorithm-based topographical biomaterials library to instruct cell fate. Proc. Natl. Acad. Sci. USA 2011, 108, 16565–16570. [Google Scholar] [CrossRef] [PubMed]
- Groen, N.; Yuan, H.; Hebels, D.G.; Kocer, G.; Mbuyi, F.; LaPointe, V.; Truckenmuller, R.; van Blitterswijk, C.A.; Habibovic, P.; de Boer, J. Linking the Transcriptional Landscape of Bone Induction to Biomaterial Design Parameters. Adv. Mater. 2017, 29. [Google Scholar] [CrossRef] [PubMed]
- Engfeldt, B.; Hjerpe, A. Glycosaminoglycans and proteoglycans of human bone tissue at different stages of mineralization. Acta Pathol. Microbiol. Scand. A 1976, 84, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Vejlens, L. Glycosaminoglycans of human bone tissue. I. Pattern of compact bone in relation to age. Calcif. Tissue Res. 1971, 7, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Vejlens, L. Glycosaminoglycans of human bone tissue. II. Chondroitin sulphate pattern of femoral cortex, medulla and epiphyseal plate in osteopetrosis. Calcif. Tissue Res. 1972, 9, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Coulson-Thomas, Y.M.; Coulson-Thomas, V.J.; Norton, A.L.; Gesteira, T.F.; Cavalheiro, R.P.; Meneghetti, M.C.; Martins, J.R.; Dixon, R.A.; Nader, H.B. The identification of proteoglycans and glycosaminoglycans in archaeological human bones and teeth. PLoS ONE 2015, 10, e0131105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrimon, N.; Bernfield, M. Specificities of heparan sulphate proteoglycans in developmental processes. Nature 2000, 404, 725–728. [Google Scholar] [CrossRef] [PubMed]
- Bishop, J.R.; Schuksz, M.; Esko, J.D. Heparan sulphate proteoglycans fine-tune mammalian physiology. Nature 2007, 446, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Hileman, R.E.; Fromm, J.R.; Weiler, J.M.; Linhardt, R.J. Glycosaminoglycan-protein interactions: Definition of consensus sites in glycosaminoglycan binding proteins. Bioessays 1998, 20, 156–167. [Google Scholar] [CrossRef]
- Griffin, C.C.; Linhardt, R.J.; Van Gorp, C.L.; Toida, T.; Hileman, R.E.; Schubert, R.L., 2nd; Brown, S.E. Isolation and characterization of heparan sulfate from crude porcine intestinal mucosal peptidoglycan heparin. Carbohydr. Res. 1995, 276, 183–197. [Google Scholar] [CrossRef]
- Blanquaert, F.; Saffar, J.L.; Colombier, M.L.; Carpentier, G.; Barritault, D.; Caruelle, J.P. Heparan-like molecules induce the repair of skull defects. Bone 1995, 17, 499–506. [Google Scholar] [CrossRef]
- Albo, D.; Long, C.; Jhala, N.; Atkinson, B.; Granick, M.S.; Wang, T.; Meddahi, A.; Barritault, D.; Solomon, M.P. Modulation of cranial bone healing with a heparin-like dextran derivative. J. Craniofac. Surg. 1996, 7, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Barritault, D.; Gilbert-Sirieix, M.; Rice, K.L.; Sineriz, F.; Papy-Garcia, D.; Baudouin, C.; Desgranges, P.; Zakine, G.; Saffar, J.L.; van Neck, J. RGTA(R) or ReGeneraTing Agents mimic heparan sulfate in regenerative medicine: From concept to curing patients. Glycoconj. J. 2017, 34, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.A.; McDonald, M.M.; Nurcombe, V.; Little, D.G.; Cool, S.M. The use of heparan sulfate to augment fracture repair in a rat fracture model. J. Orthop. Res. 2006, 24, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Murali, S.; Rai, B.; Dombrowski, C.; Lee, J.L.; Lim, Z.X.; Bramono, D.S.; Ling, L.; Bell, T.; Hinkley, S.; Nathan, S.S.; et al. Affinity-selected heparan sulfate for bone repair. Biomaterials 2013, 34, 5594–5605. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Fyrner, T.; Chen, F.; Alvarez, Z.; Sleep, E.; Chun, D.S.; Weiner, J.A.; Cook, R.W.; Freshman, R.D.; Schallmo, M.S.; et al. Sulfated glycopeptide nanostructures for multipotent protein activation. Nat. Nanotechnol. 2017, 12, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Donati, D.; Zolezzi, C.; Tomba, P.; Vigano, A. Bone grafting: Historical and conceptual review, starting with an old manuscript by Vittorio Putti. Acta Orthop. 2007, 78, 19–25. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Tissue | Search Term | Number of Hits |
|---|---|---|
| Periosteum | Periosteum [Title] OR periosteal [Title] | 2559 |
| Osseous | Osseous [Title] | 6091 |
| Endosteum | Endosteum [Title] OR endosteal [Title] | 446 |
| Bone marrow | “bone marrow” [Title] | 63,165 |
| Bone marrow (related to bone research) | (“bone marrow” [Title]) AND ((“bone regeneration” [Title/Abstract]) OR (“bone healing” [Title/Abstract]) OR (“bone tissue engineering” [Title/Abstract]) OR (“fracture healing” [Title/Abstract]) OR (“nonunion” [Title/Abstract]) OR (“non-union” [Title/Abstract])) | 805 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le, B.Q.; Nurcombe, V.; Cool, S.M.; Van Blitterswijk, C.A.; De Boer, J.; LaPointe, V.L.S. The Components of Bone and What They Can Teach Us about Regeneration. Materials 2018, 11, 14. https://doi.org/10.3390/ma11010014
Le BQ, Nurcombe V, Cool SM, Van Blitterswijk CA, De Boer J, LaPointe VLS. The Components of Bone and What They Can Teach Us about Regeneration. Materials. 2018; 11(1):14. https://doi.org/10.3390/ma11010014
Chicago/Turabian StyleLe, Bach Quang, Victor Nurcombe, Simon McKenzie Cool, Clemens A. Van Blitterswijk, Jan De Boer, and Vanessa Lydia Simone LaPointe. 2018. "The Components of Bone and What They Can Teach Us about Regeneration" Materials 11, no. 1: 14. https://doi.org/10.3390/ma11010014
APA StyleLe, B. Q., Nurcombe, V., Cool, S. M., Van Blitterswijk, C. A., De Boer, J., & LaPointe, V. L. S. (2018). The Components of Bone and What They Can Teach Us about Regeneration. Materials, 11(1), 14. https://doi.org/10.3390/ma11010014