New Parameters to Quantitatively Express the Invasiveness of Bacterial Strains from Implant-Related Orthopaedic Infections into Osteoblast Cells

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
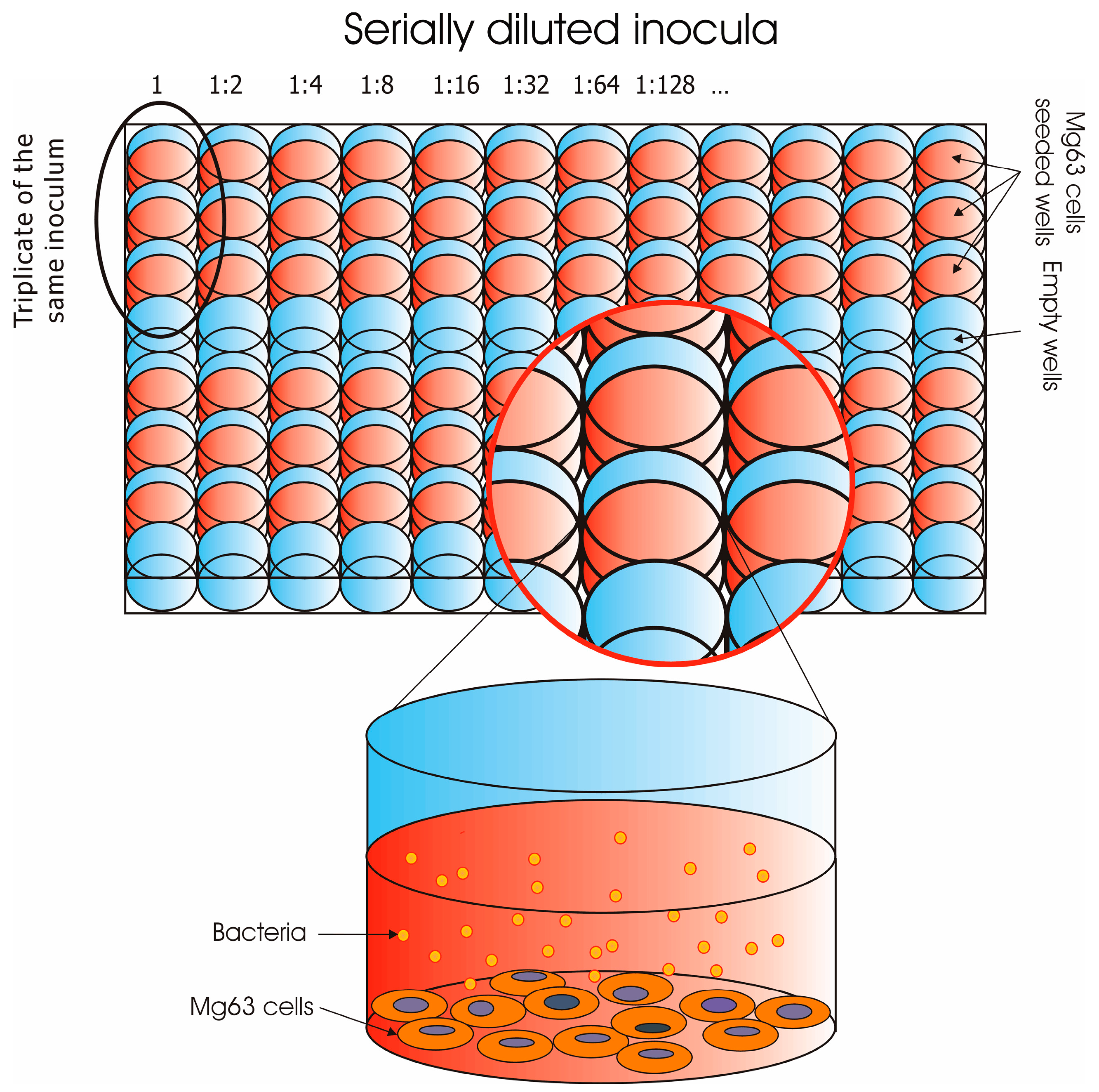
2.2. Preparation of the Bacterial Inoculum for Cell Invasion Assays
2.3. MG63 Cell Culture
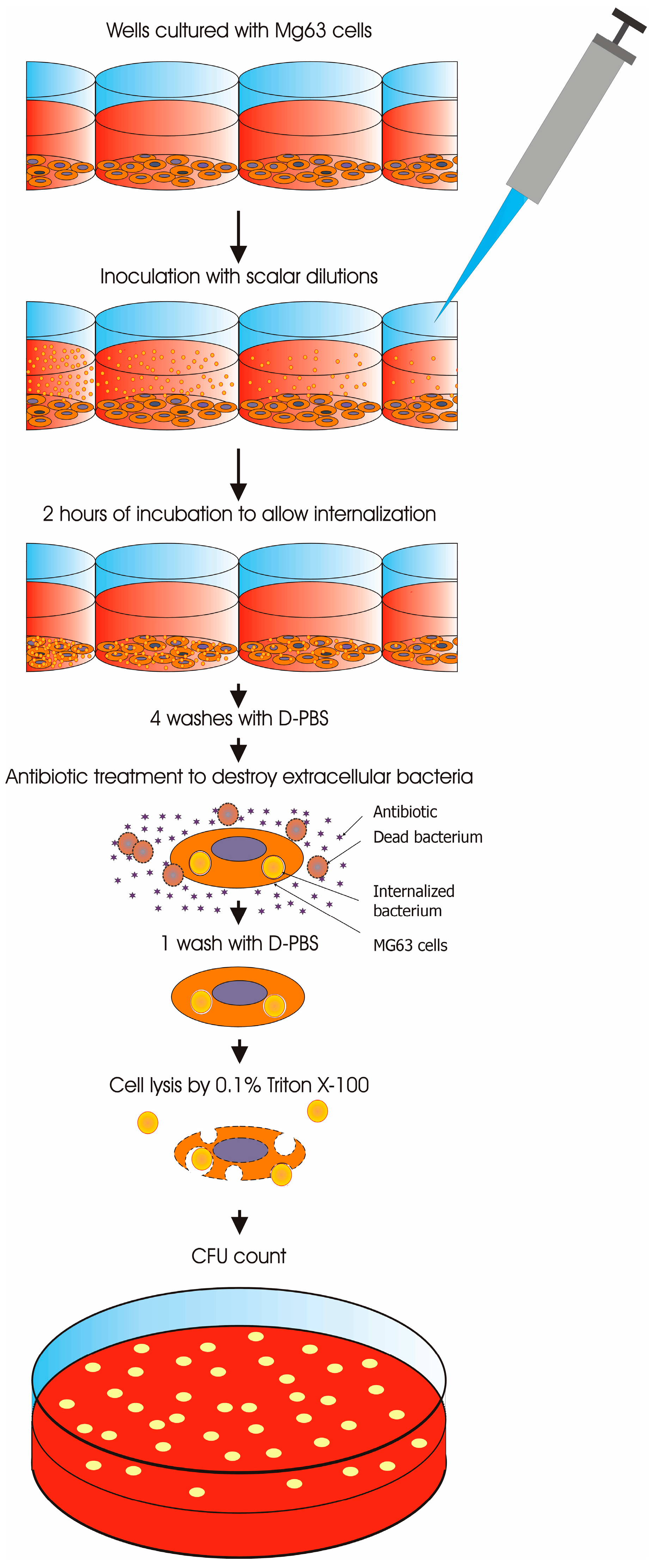
2.4. Invasion Assay of Osteoblasts in 96-Well Plates
2.5. Statistics
3. Results
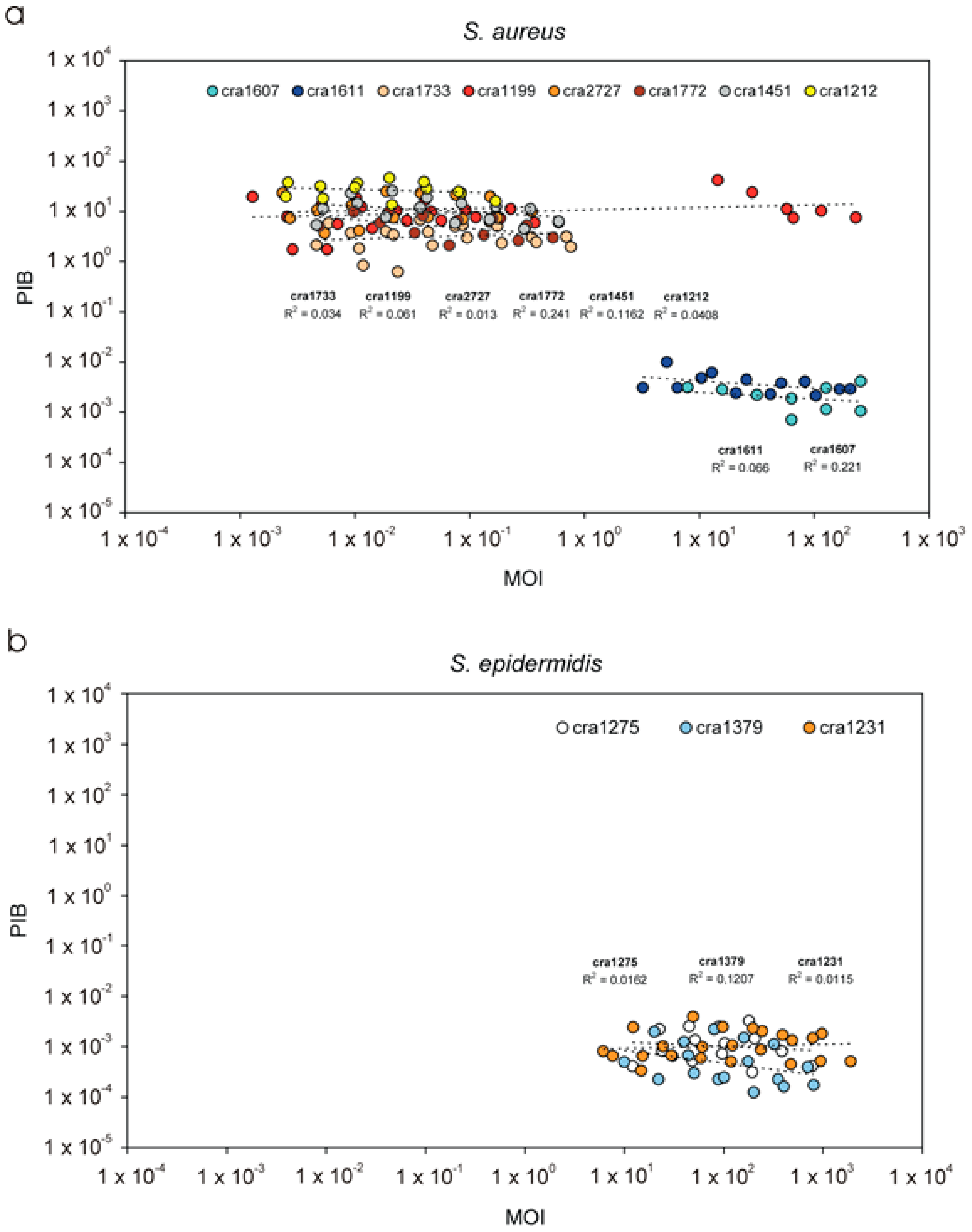
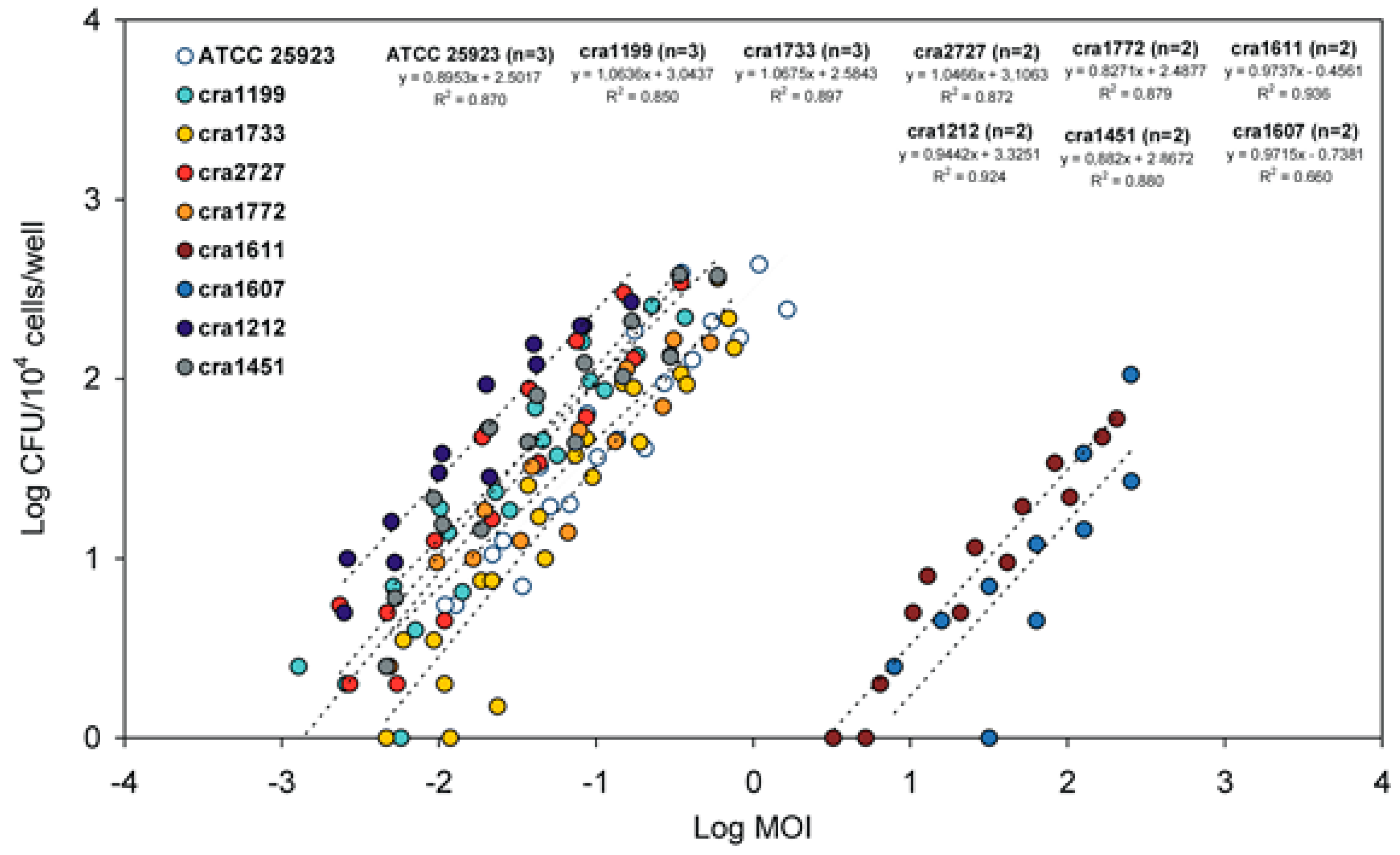
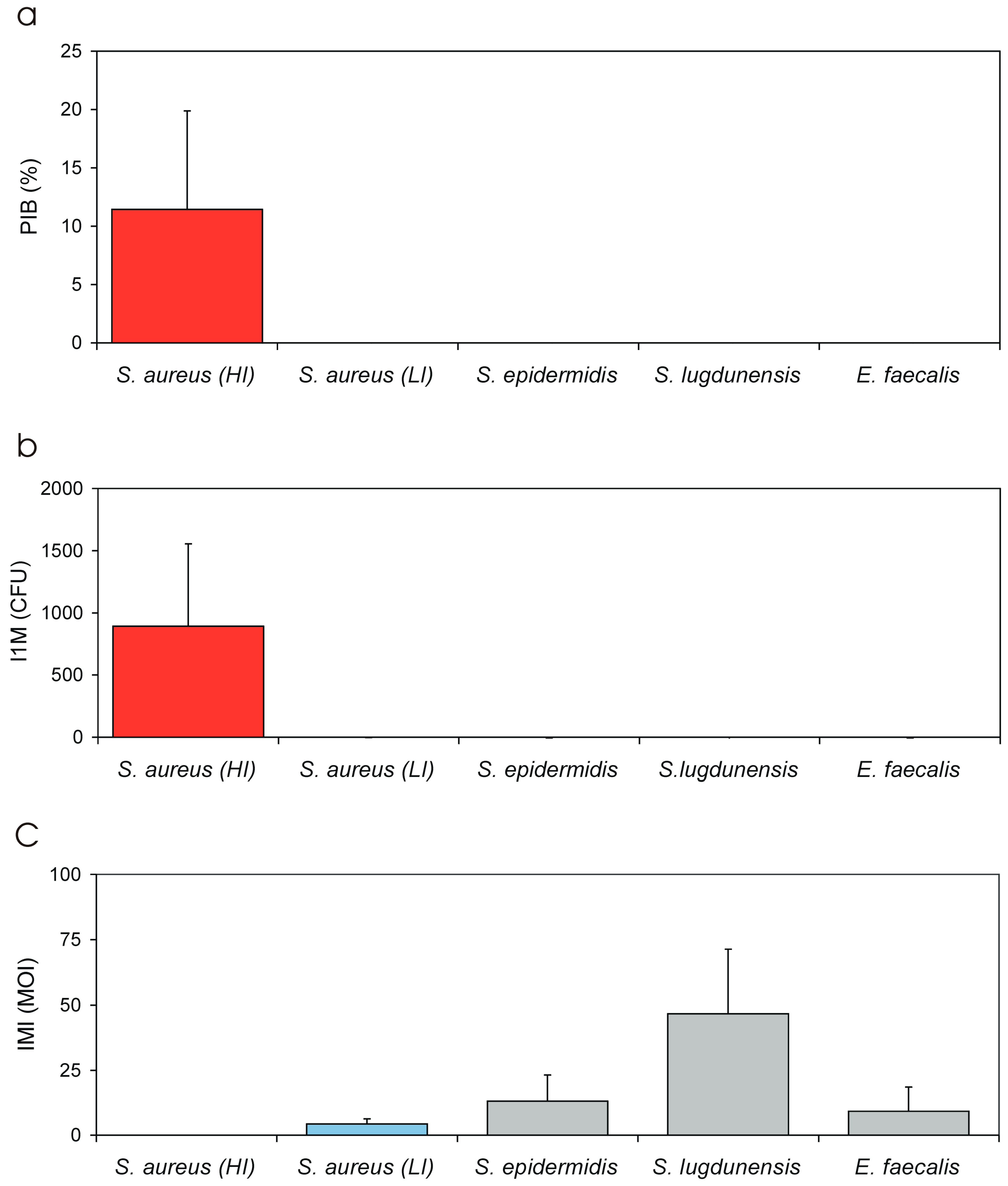
3.1. PIB Values vs. Inoculum Size (MOI)
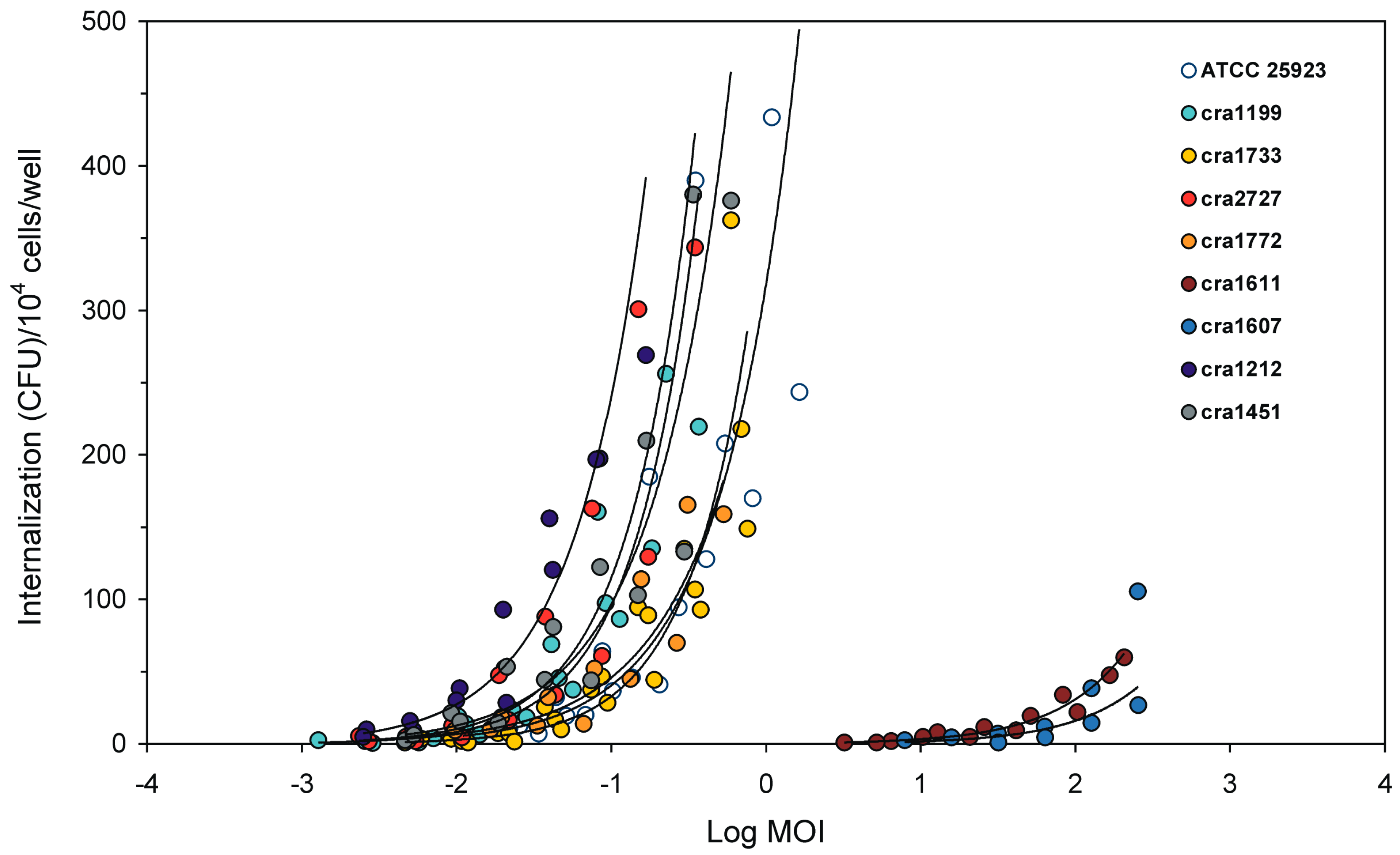
3.2. Internalised CFU vs. MOI Regression Curves
3.3. New Parameters for Describing the Internalization Efficiency
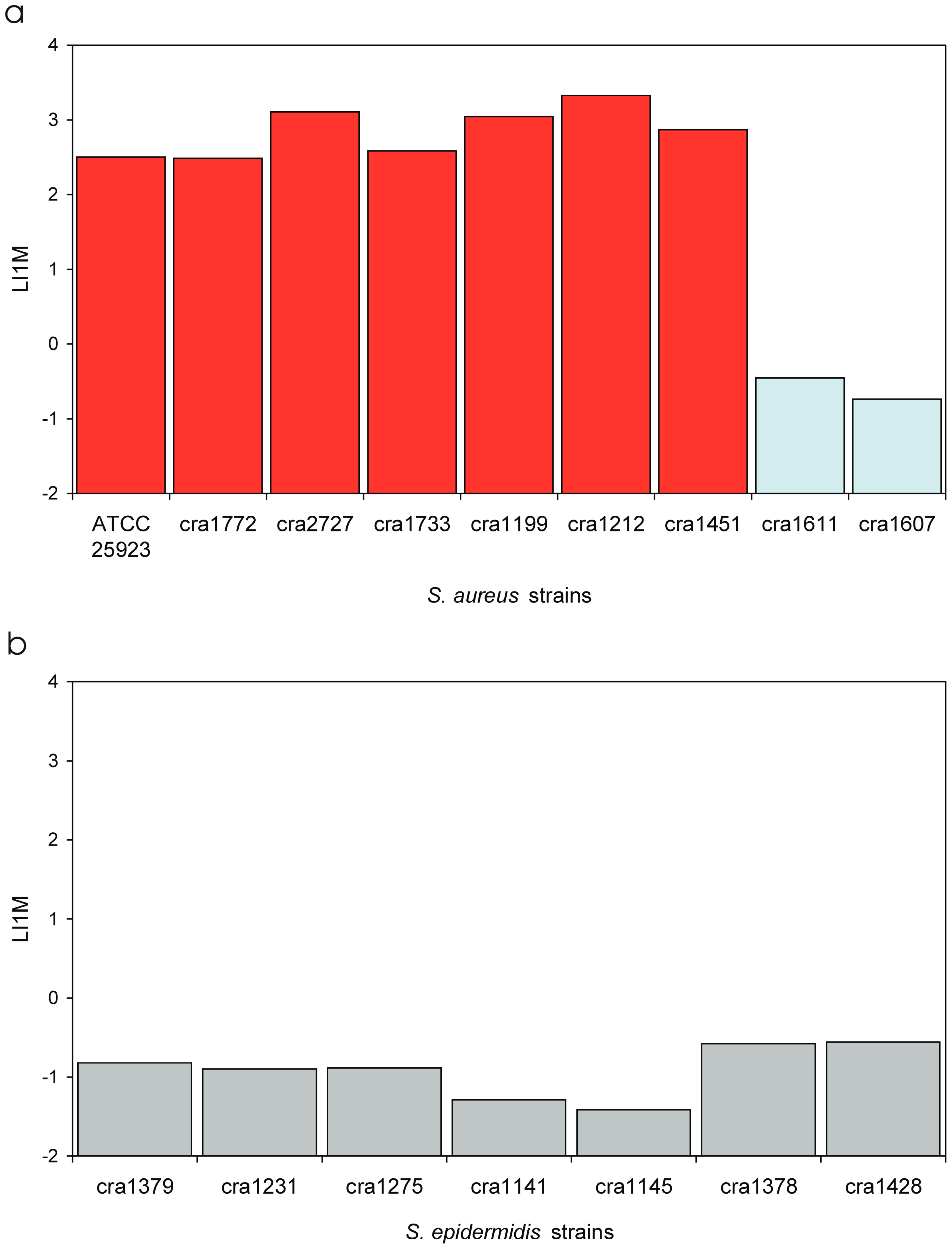
3.4. Invasiveness of S. aureus Strains
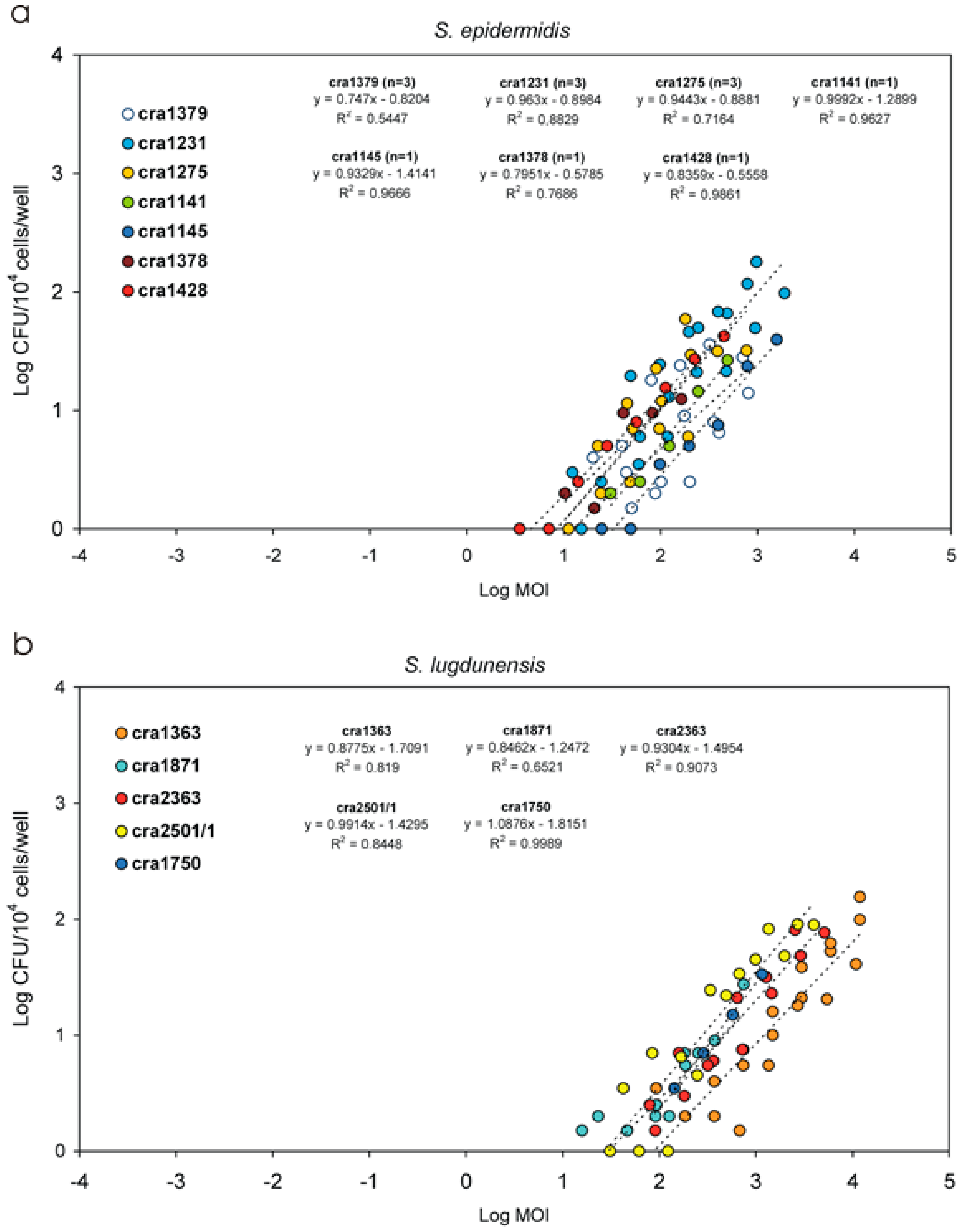
3.5. Invasiveness of S. epidermidis Strains
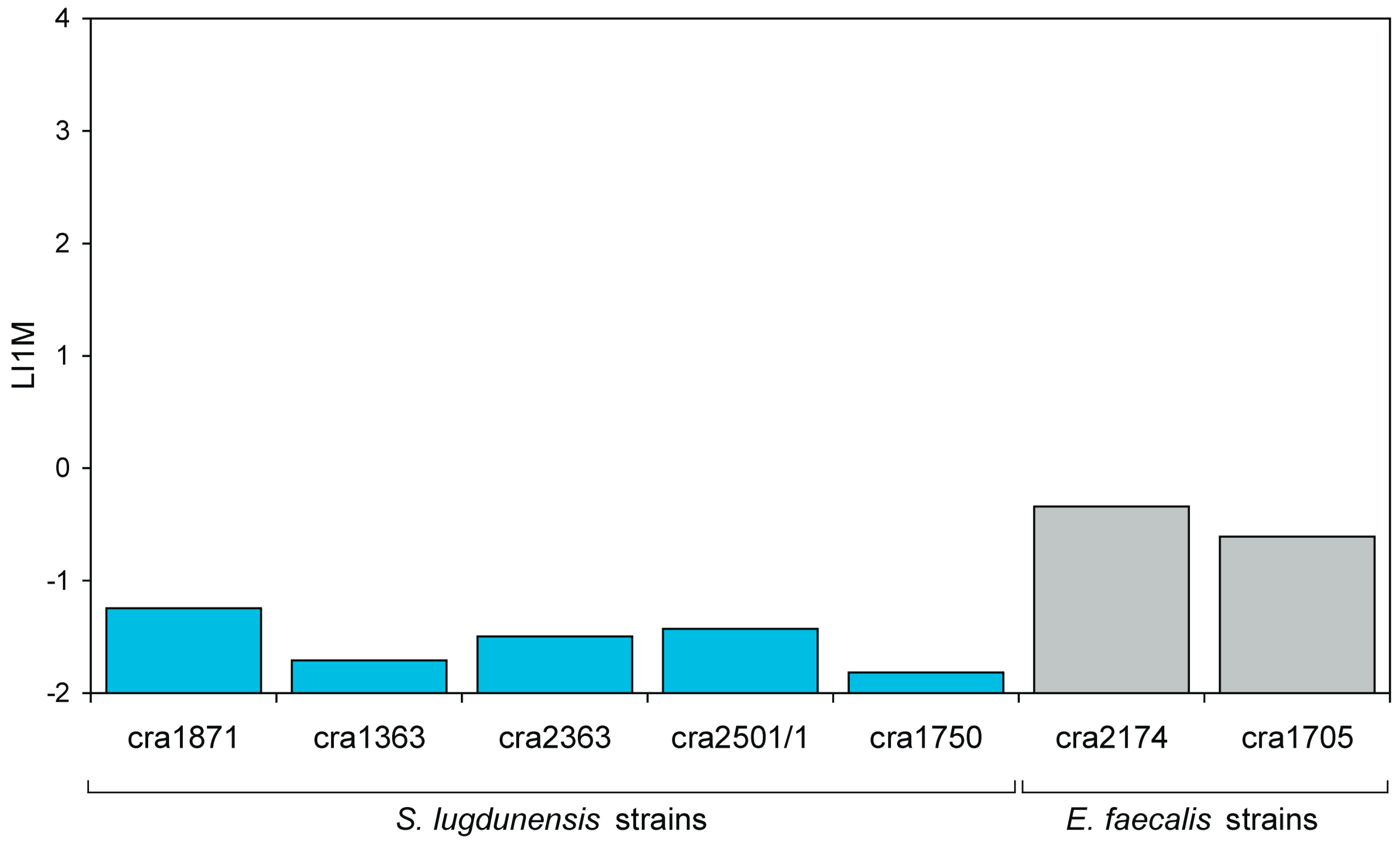
3.6. Invasiveness of S. lugdunensis Strains
3.7. Invasiveness of E. faecalis Strains
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bhavsar, A.P.; Guttman, J.A.; Finlay, B.B. Manipulation of host-cell pathways by bacterial pathogens. Nature 2007, 449, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Ellington, J.K.; Harris, M.; Hudson, M.C.; Vishin, S.; Webb, L.X.; Sherertz, R. Intracellular Staphylococcus aureus and antibiotic resistance: Implications for treatment of staphylococcal osteomyelitis. J. Orthop. Res. 2006, 24, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Rollin, G.; Tan, X.; Tros, F.; Dupuis, M.; Nassif, X.; Charbit, A.; Coureuil, M. Intracellular survival of Staphylococcus aureus in endothelial cells: A matter of growth or persistence. Front. Microbiol. 2017, 8, 1354. [Google Scholar] [CrossRef] [PubMed]
- Campoccia, D.; Testoni, F.; Ravaioli, S.; Cangini, I.; Maso, A.; Speziale, P.; Montanaro, L.; Visai, L.; Arciola, C.R. Orthopedic implant infections: Incompetence of Staphylococcus epidermidis, Staphylococcus lugdunensis, and Enterococcus faecalis to invade osteoblasts. J. Biomed. Mater. Res. A 2016, 104, 788–801. [Google Scholar] [CrossRef] [PubMed]
- Valour, F.; Trouillet-Assant, S.; Rasigade, J.P.; Lustig, S.; Chanard, E.; Meugnier, H.; Tigaud, S.; Vandenesch, F.; Etienne, J.; Ferry, T.; et al. Staphylococcus epidermidis in orthopedic device infections: The role of bacterial internalization in human osteoblasts and biofilm formation. PLoS ONE 2013, 8, e67240. [Google Scholar] [CrossRef] [PubMed]
- Maali, Y.; Martins-Simões, P.; Valour, F.; Bouvard, D.; Rasigade, J.P.; Bes, M.; Haenni, M.; Ferry, T.; Laurent, F.; Trouillet-Assant, S. Pathophysiological mechanisms of Staphylococcus non-aureus bone and joint infection: Interspecies homogeneity and specific behavior of S. pseudintermedius. Front. Microbiol. 2016, 7, 1063. [Google Scholar] [CrossRef] [PubMed]
- Khalil, H.; Williams, R.J.; Stenbeck, G.; Henderson, B.; Meghji, S.; Nair, S.P. Invasion of bone cells by Staphylococcus epidermidis. Microbes Infect. 2007, 9, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Steinbacher, T.; Peters, G.; Heilmann, C.; Becker, K. The adhesive properties of the Staphylococcus lugdunensis multifunctional autolysin AtlL and its role in biofilm formation and internalization. Int. J. Med. Microbiol. 2015, 305, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Millán, D.; Chiriboga, C.; Patarroyo, M.A.; Fontanilla, M.R. Enterococcus faecalis internalization in human umbilical vein endothelial cells (HUVEC). Microb. Pathog. 2013, 57, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.M.; Potts, J.R.; Josefsson, E.; Massey, R.C. Staphylococcus aureus host cell invasion and virulence in sepsis is facilitated by the multiple repeats within FnBPA. PLoS Pathog. 2010, 6, e1000964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valour, F.; Rasigade, J.P.; Trouillet-Assant, S.; Gagnaire, J.; Bouaziz, A.; Karsenty, J.; Lacour, C.; Bes, M.; Lustig, S.; Bénet, T.; et al. Lyon BJI Study Group. Delta-toxin production deficiency in Staphylococcus aureus: A diagnostic marker of bone and joint infection chronicity linked with osteoblast invasion and biofilm formation. Clin. Microbiol. Infect. 2015, 21, 568.e1–568.e11. [Google Scholar] [CrossRef] [PubMed]
- Campoccia, D.; Montanaro, L.; von Eiff, C.; Pirini, V.; Ravaioli, S.; Becker, K.; Arciola, C.R. Cluster analysis of ribotyping profiles of Staphylococcus epidermidis isolates recovered from foreign body-associated orthopedic infections. J. Biomed. Mater. Res. A 2009, 88, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Arciola, C.R.; Campoccia, D.; Baldassarri, L.; Pirini, V.; Huebner, J.; Montanaro, L. The role of Enterococcus faecalis in orthopaedic peri-implant infections demonstrated by automated ribotyping and cluster analysis. Biomaterials 2007, 28, 3987–3995. [Google Scholar] [CrossRef] [PubMed]
- Montanaro, L.; Ravaioli, S.; Ruppitsch, W.; Campoccia, D.; Pietrocola, G.; Visai, L.; Speziale, P.; Allerberger, F.; Arciola, C.R. Molecular characterization of a prevalent ribocluster of methicillin-sensitive Staphylococcus aureus from orthopedic implant infections. Correspondence with MLST CC30. Front. Cell. Infect. Microbiol. 2016, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Campoccia, D.; Montanaro, L.; Pirini, V.; Ravaioli, S.; Arciola, C.R. Prevalence of genes for aminoglycoside-modifying enzymes in Staphylococcus epidermidis isolates from orthopedic postsurgical and implant-related infections. J. Biomed. Mater. Res. A 2009, 88, 654–663. [Google Scholar] [CrossRef] [PubMed]
- Campoccia, D.; Montanaro, L.; Ravaioli, S.; Pirini, V.; Cangini, I.; Arciola, C.R. Exopolysaccharide production by Staphylococcus epidermidis and its relationship with biofilm extracellular DNA. Int. J. Artif. Organs 2011, 34, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.G.; Murray, S.; Pascoe, B.; Bray, J.; Meric, G.; Mageiros, L.; Wilkinson, T.S.; Jeeves, R.; Rohde, H.; Schwarz, S.; et al. Biofilm morphotypes and population structure among Staphylococcus epidermidis from commensal and clinical samples. PLoS ONE 2016, 11, e0151240. [Google Scholar]
- De Kort, G.; Bolton, A.; Martin, G.; Stephen, J.; van de Klundert, J.A. Invasion of rabbit ileal tissue by Enterobacter cloacae varies with the concentration of OmpX in the outer membrane. Infect. Immun. 1994, 62, 4722–4726. [Google Scholar] [PubMed]
- Donnarumma, G.; Paoletti, I.; Buommino, E.; Tufano, M.A.; Baroni, A. Alpha-MSH reduces the internalization of Staphylococcus aureus and down-regulates HSP 70, integrins and cytokine expression in human keratinocyte cell lines. Exp. Dermatol. 2004, 13, 748–754. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acronym | Description | Meaning |
|---|---|---|
| NIB | Number of internalised bacteria | Number of internalised bacteria (at a given MOI). Usually expressed in terms of CFU per well or CFU per number of eukaryotic cells, NIB is easily obtained, but its use presents limitations. Its value is influenced by the MOI in use. The assessment of internalization of bacteria belonging to different species is usually performed at different MOI, thus direct comparison of NIB among strains is hampered. Obstacles are met when strains of the same species exhibit remarkable differences in invasiveness, ideally needing different inoculum MOI. |
| PIB | Per cent of internalised bacteria | PIB value represents the per cent fraction of the inoculum that is taken up by the eukaryotic cells. Its value is directly proportional to the invasiveness of the test strain towards a specific eukaryotic cell type. Based on our current results, PIB is not significantly affected by MOI and is more appropriate than NIB for strains comparison. |
| Species | Strain | Ribogroup | MLST CC | spa Type | spa CC | Origin |
|---|---|---|---|---|---|---|
| S. aureus | ATCC 25923 | - | CC30 * | t021 | CC021/012 | Clinical |
| cra1733 | cra-119-S-8 | K | ||||
| cra2727 | H | |||||
| cra1772 | t298 | IF | ||||
| cra1199 | t012 | EF | ||||
| cra1451 | cra-138-S-2 | H | ||||
| cra1611 | IF | |||||
| cra1607 | H | |||||
| cra1212 | cra-53-S-7 | EF |
| Species | Strain | Ribogroup | Origin |
|---|---|---|---|
| S. epidermidis | cra1379 | cra-63-S-7 | FI |
| cra1231 | H | ||
| cra1275 | cra-63-S-4 | PSI | |
| cra1141 | cra-92-S-5 | H | |
| cra1145 | cra-122-S-2 | H | |
| cra1378 | cra-119-S-4 | H | |
| cra1428 | cra-80-S-1 | K | |
| S. lugdunensis | cra1871 | cra-62-S-1 | FI |
| cra1363 | FI | ||
| cra2363 | cra-64-S-8 | FI | |
| cra2501/1 | FE | ||
| cra1750 | cra-74-S-5 | No MD | |
| cra1871 | cra-62-S-1 | FI | |
| E. faecalis | cra2174 | cra-116-S-1 | H |
| cra1705 | cra-115-S-8 | FI |
| Acronym | Description | Meaning |
|---|---|---|
| IMI | Internalization minimal inoculum | IMI is a virtual value extrapolated from the equation of the regression curve achieved by plotting Log MOI vs. Log (CFU). IMI corresponds to the lowest MOI required for the internalization of a single bacterium. The lower the value the higher the invasiveness of the strain. IMI lowest value, in our system, would correspond to 0.0001:1 MOI, i.e., a suspension containing a single bacterium for 104 eukaryotic cells. Being inversely related to invasiveness, IMI could advantageously be used to express the resistance to phagocytosis. |
| I1M | Internalization at 1:1 MOI inoculum | I1M is a virtual value, extrapolated from the equation of the regression curve achieved by plotting Log MOI vs. Log (CFU). I1M corresponds to the number of bacteria internalised when hypothetically exposing each eukaryotic cell to a single bacterium (i.e., when using a 1:1 MOI). Its value is directly proportional to the degree of invasiveness of the strain. |
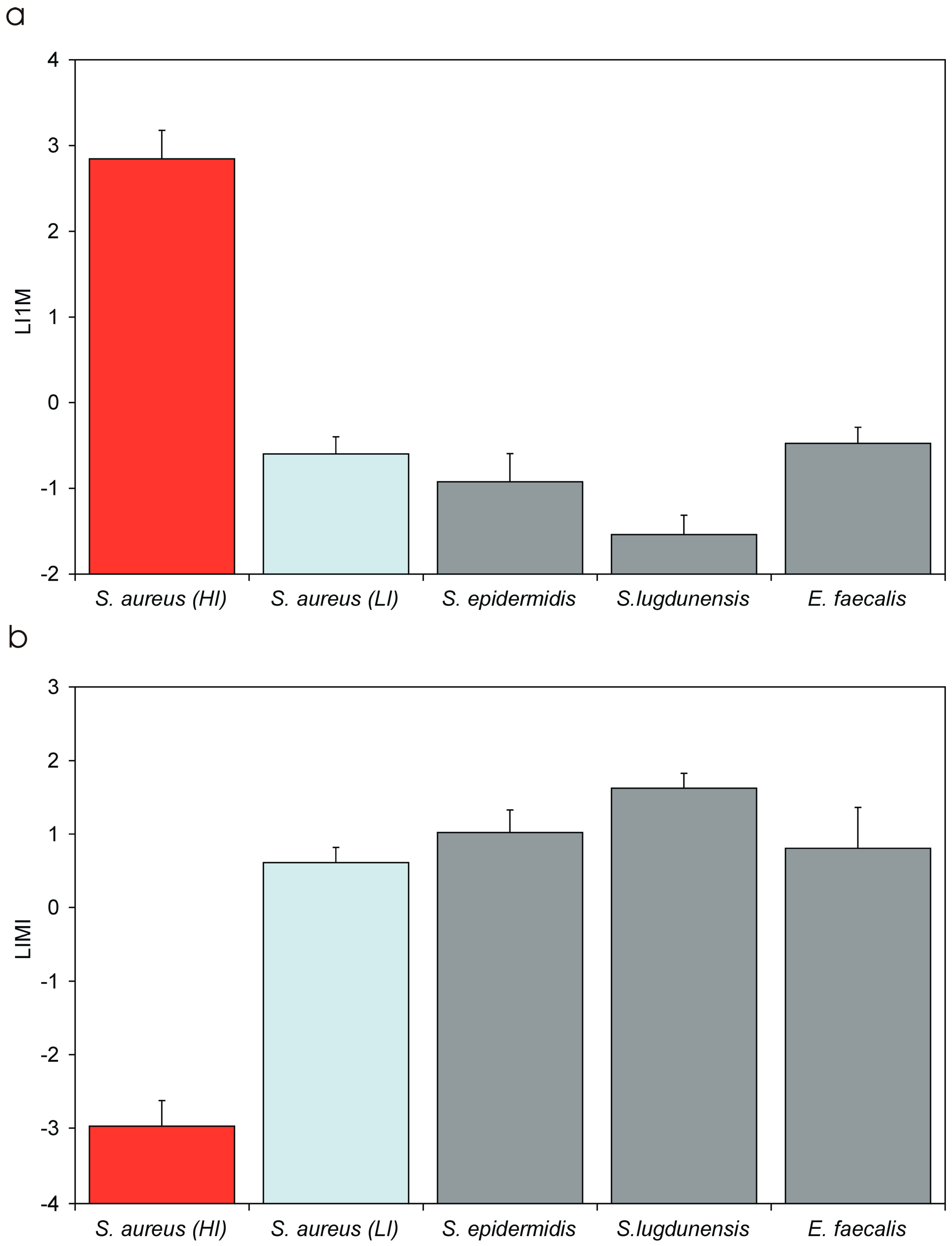
| LIMI | Log10 of the IMI value | LIMI is promptly obtained from the regression curve of Log MOI vs. Log (CFU). Its value, expected in the range from −3 to 4, is inversely proportional to the degree of invasiveness of the bacterial strain. LIMI exhibits a lower coefficient of variation with respect to IMI, PIB and I1M, and it more closely approaches a normal distribution. LIMI can be easily transformed to obtain the corresponding IMI value. |
| LI1M | Log10 of the I1M value | LI1M is promptly obtained from the regression curve of Log MOI vs. Log (CFU). Its value, expected in the range from −4 to 3, is directly proportional to the degree of invasiveness of the bacterial strain. LI1M exhibits a lower coefficient of variation with respect to IMI, PIB and I1M, and it more closely approaches a normal distribution. It can be easily transformed to obtain the corresponding I1M value. |
| Bacterium | PIB [Mean ± S.D. 1/(C.V.)] | IMI [Mean ± S.D./(C.V.)] | I1M [Mean ± S.D./(C.V.)] | LIMI [Mean ± S.D./(C.V.)] | LI1M [Mean ± S.D./(C.V.)] |
|---|---|---|---|---|---|
| S. aureus (HI + LI) | 8.89 ± 8.88 (99.9%) | 0.97 ± 2.04 (211.1%) | 693.67 ± 696.23 (100.4) | −2.18 ± 1.61 (74.1%) | 2.08 ± 1.15 (74.3%) |
| S. aureus (HI) | 11.43 ± 8.45 h (73.9%) | 0.0014 ± 0.0012 l (83.2%) | 891.79 ± 663.52 h (74.4%) | −2.97 ± 0.35 a,b,c,d (11.7%) | 2.85 ± 0.33 a,b,c,d (11.6%) |
| S. aureus (LI) | 0.0029 ± 0.0016 (43.9%) | 4.34 ± 1.99 (45.7%) | 0.27 ± 0.12 (44.4%) | 0.61 ± 0.21 a,f (33.5%) | −0.60 ± 0.20 a,f (33.4%) |
| S. epidermidis | 0.0010 ± 0.0005 (49.9%) | 13.2 ± 10.0 (76.1%) | 0.15 ± 0.09 (63.0%) | 1.02 ± 0.30 b,e (29.6%) | −0.92 ± 0.33 b,e (35.5%) |
| S. lugdunensis | 0.00027 ± 0.00012 l (46.2%) | 46.6 ± 24.7 h (53.0%) | 0.032 ± 0.016 l (50.8%) | 1.63 ± 0.20 c,e,f (12.4%) | −1.54 ± 0.23 c,e,f,g (14.7) |
| E. faecalis | 0.0011 ± 0.0011 (95%) | 9.2 ± 9.37 (101.5%) | 0.35 ± 0.15 (42.1%) | 0.81 ± 0.55 d (68.6%) | −0.47 ± 0.19 d,g (39.8%) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campoccia, D.; Montanaro, L.; Ravaioli, S.; Cangini, I.; Testoni, F.; Visai, L.; Arciola, C.R. New Parameters to Quantitatively Express the Invasiveness of Bacterial Strains from Implant-Related Orthopaedic Infections into Osteoblast Cells. Materials 2018, 11, 550. https://doi.org/10.3390/ma11040550
Campoccia D, Montanaro L, Ravaioli S, Cangini I, Testoni F, Visai L, Arciola CR. New Parameters to Quantitatively Express the Invasiveness of Bacterial Strains from Implant-Related Orthopaedic Infections into Osteoblast Cells. Materials. 2018; 11(4):550. https://doi.org/10.3390/ma11040550
Chicago/Turabian StyleCampoccia, Davide, Lucio Montanaro, Stefano Ravaioli, Ilaria Cangini, Francesca Testoni, Livia Visai, and Carla Renata Arciola. 2018. "New Parameters to Quantitatively Express the Invasiveness of Bacterial Strains from Implant-Related Orthopaedic Infections into Osteoblast Cells" Materials 11, no. 4: 550. https://doi.org/10.3390/ma11040550
APA StyleCampoccia, D., Montanaro, L., Ravaioli, S., Cangini, I., Testoni, F., Visai, L., & Arciola, C. R. (2018). New Parameters to Quantitatively Express the Invasiveness of Bacterial Strains from Implant-Related Orthopaedic Infections into Osteoblast Cells. Materials, 11(4), 550. https://doi.org/10.3390/ma11040550