The Impact Behaviour of Crab Carapaces in Relation to Morphology


 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
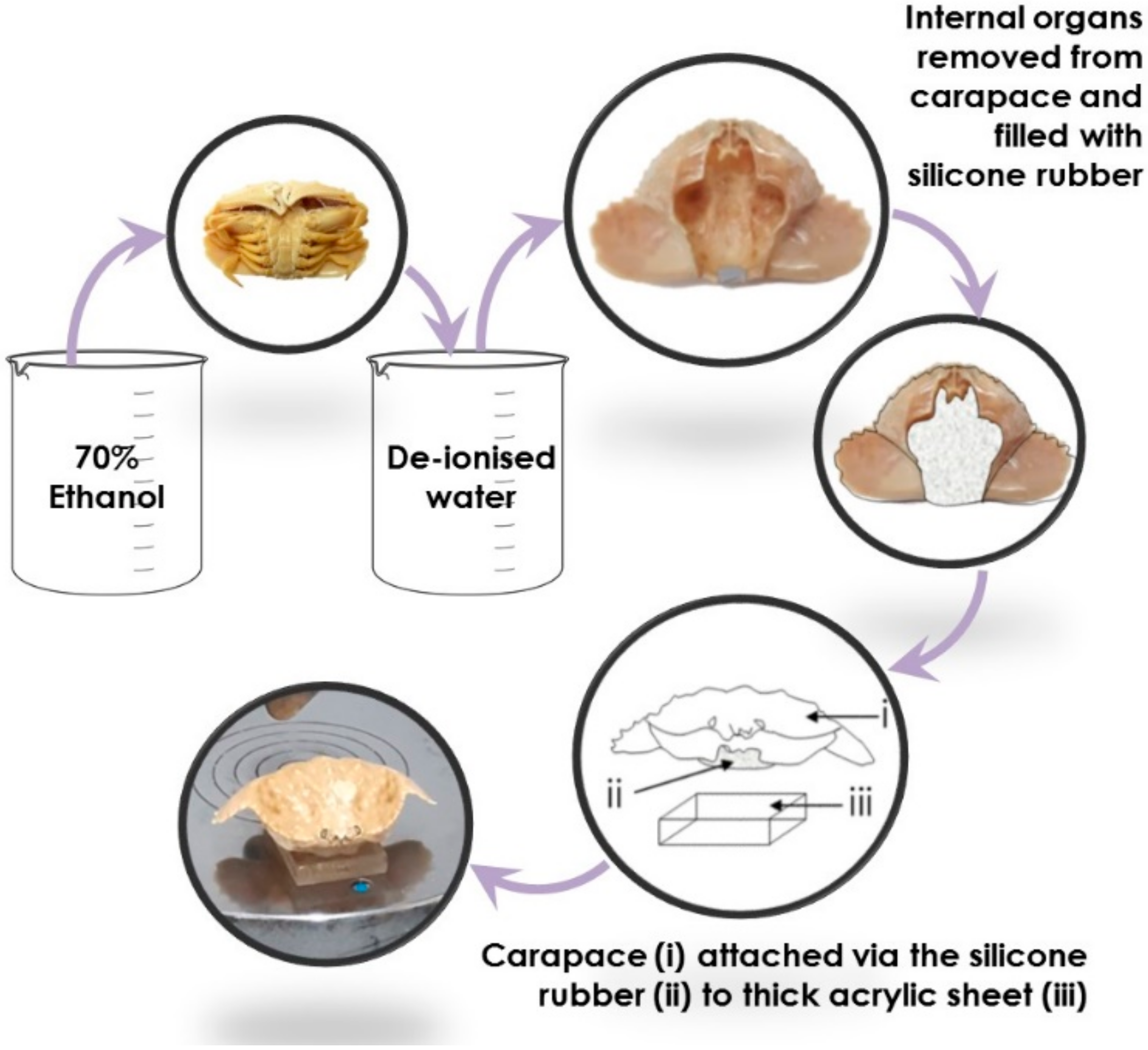
2.1. Collection and Identification of Crabs
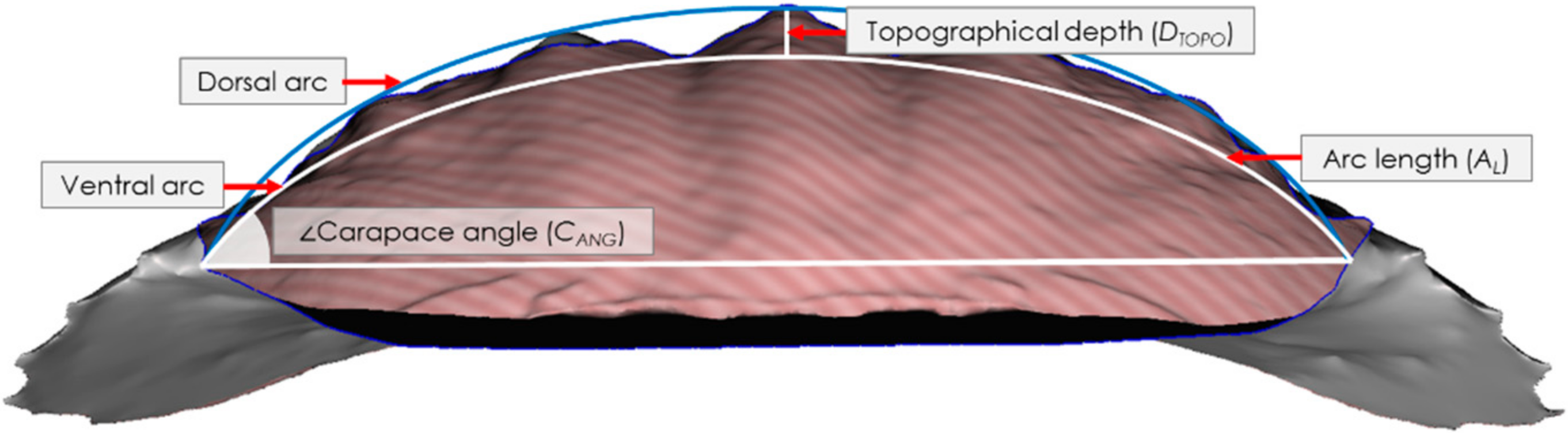
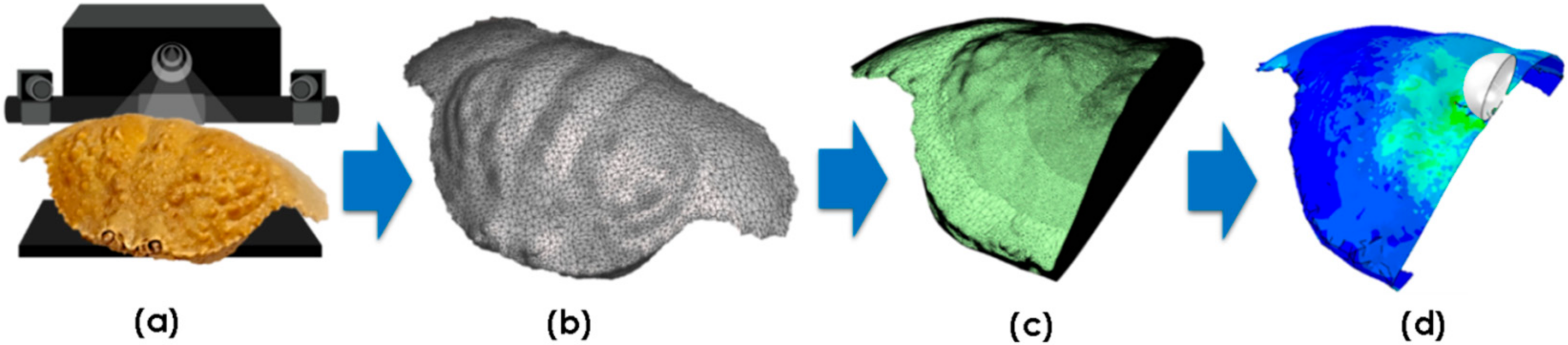
2.2. 3D Imaging and Morphometric Analyses
2.3. Impact Testing
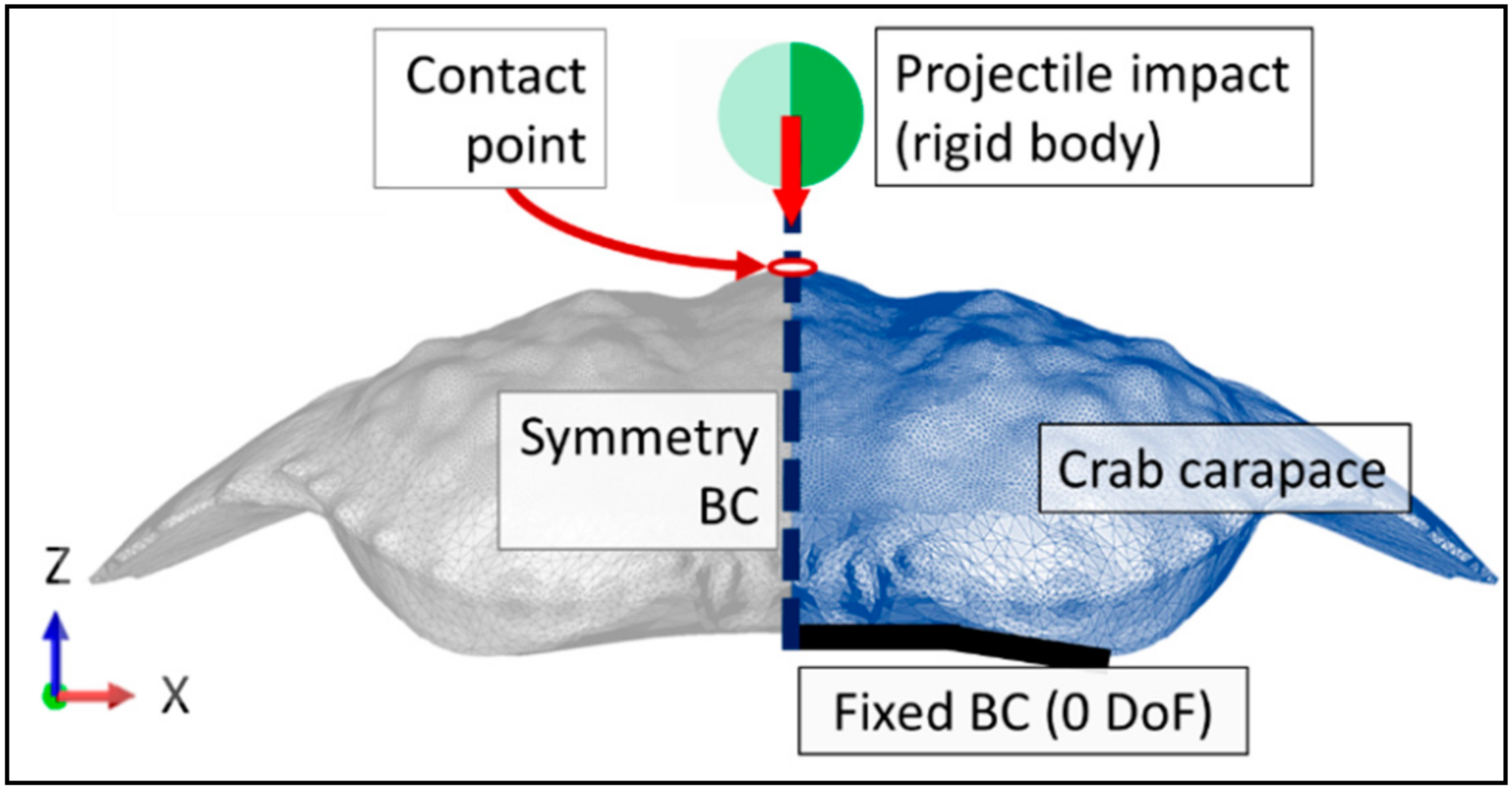
2.4. Impact Simulations
3. Results
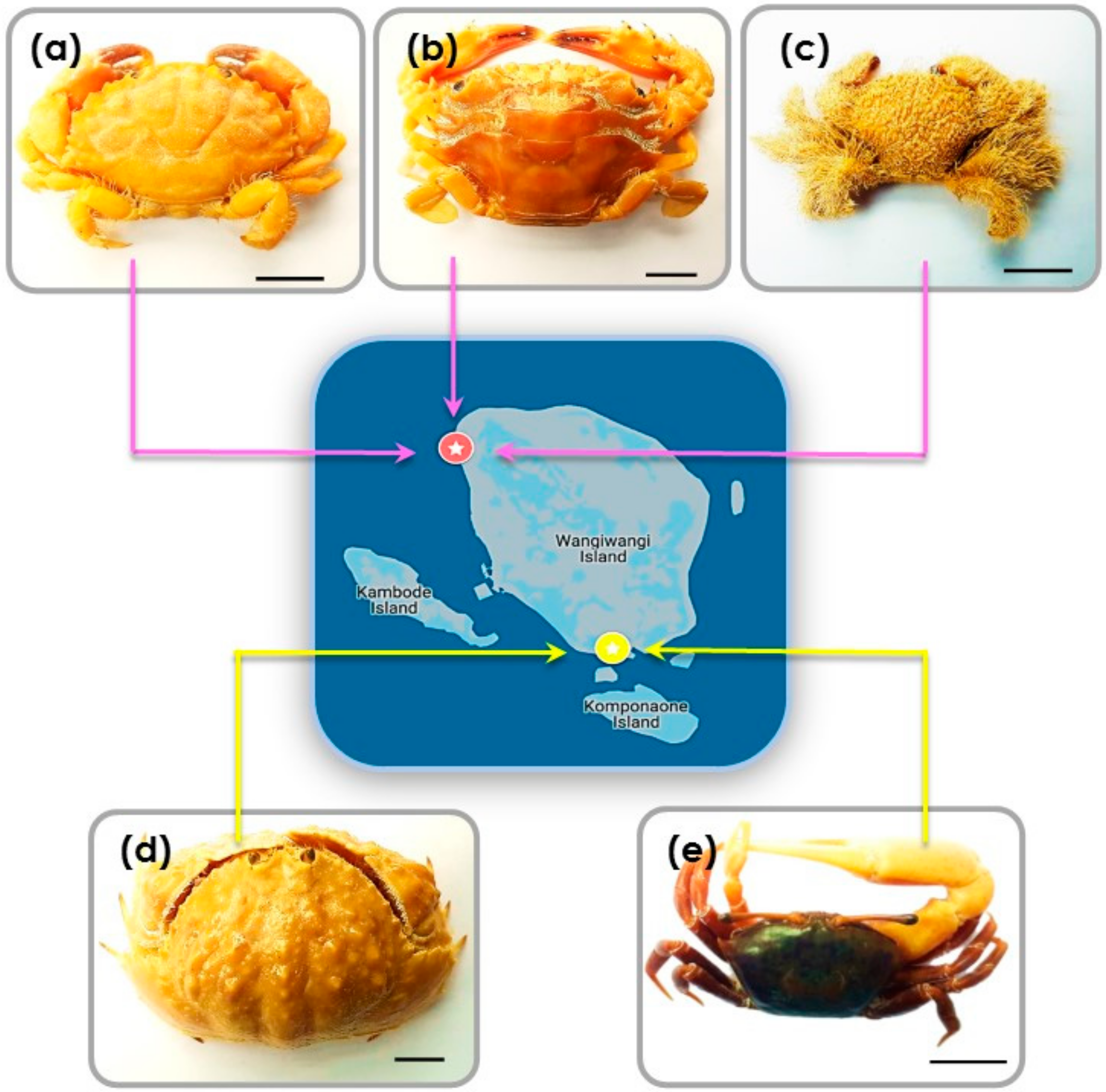
3.1. Crab Species and Habitat Descriptions
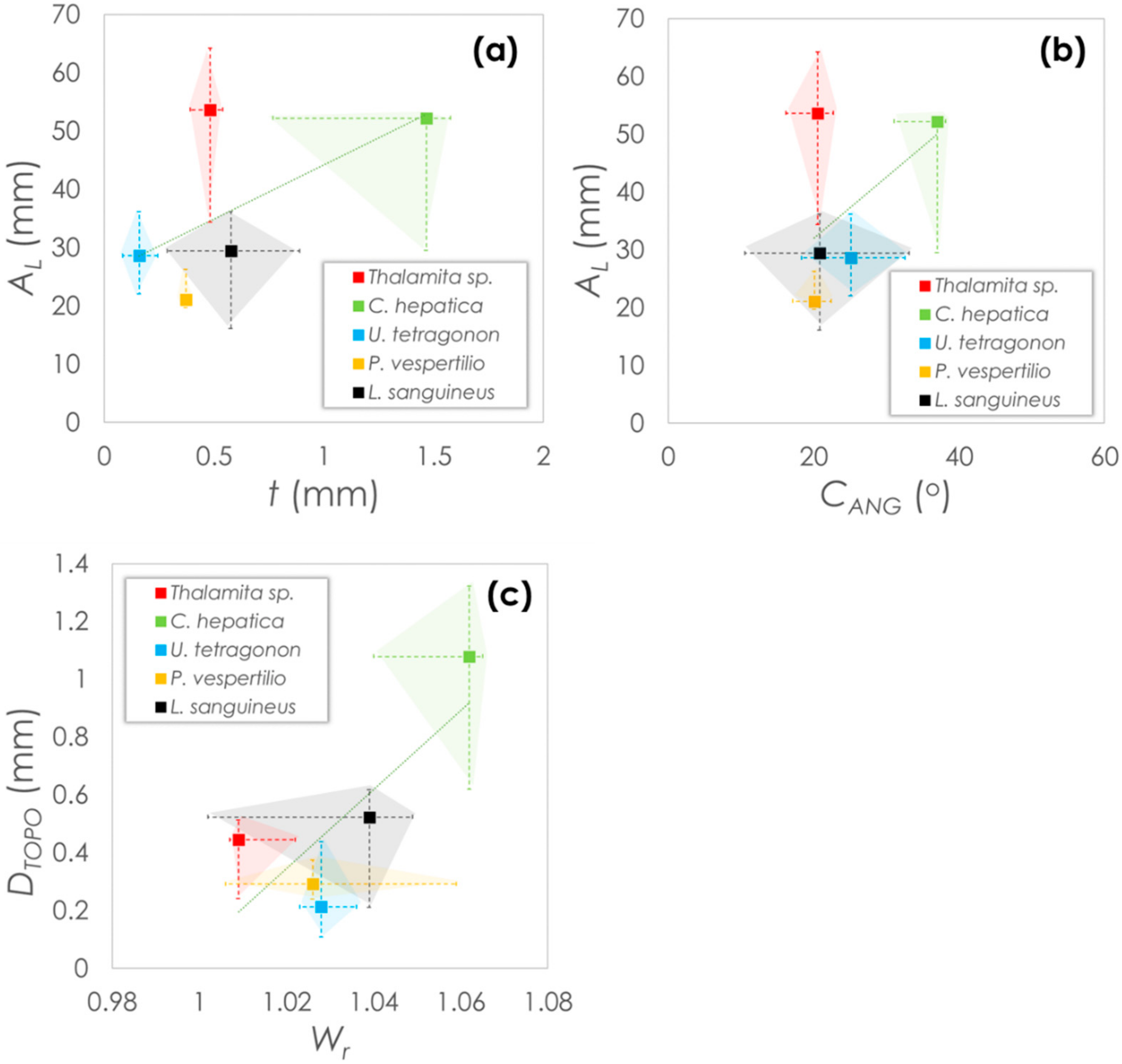
3.2. Morphometric Analysis of Crab Carapaces
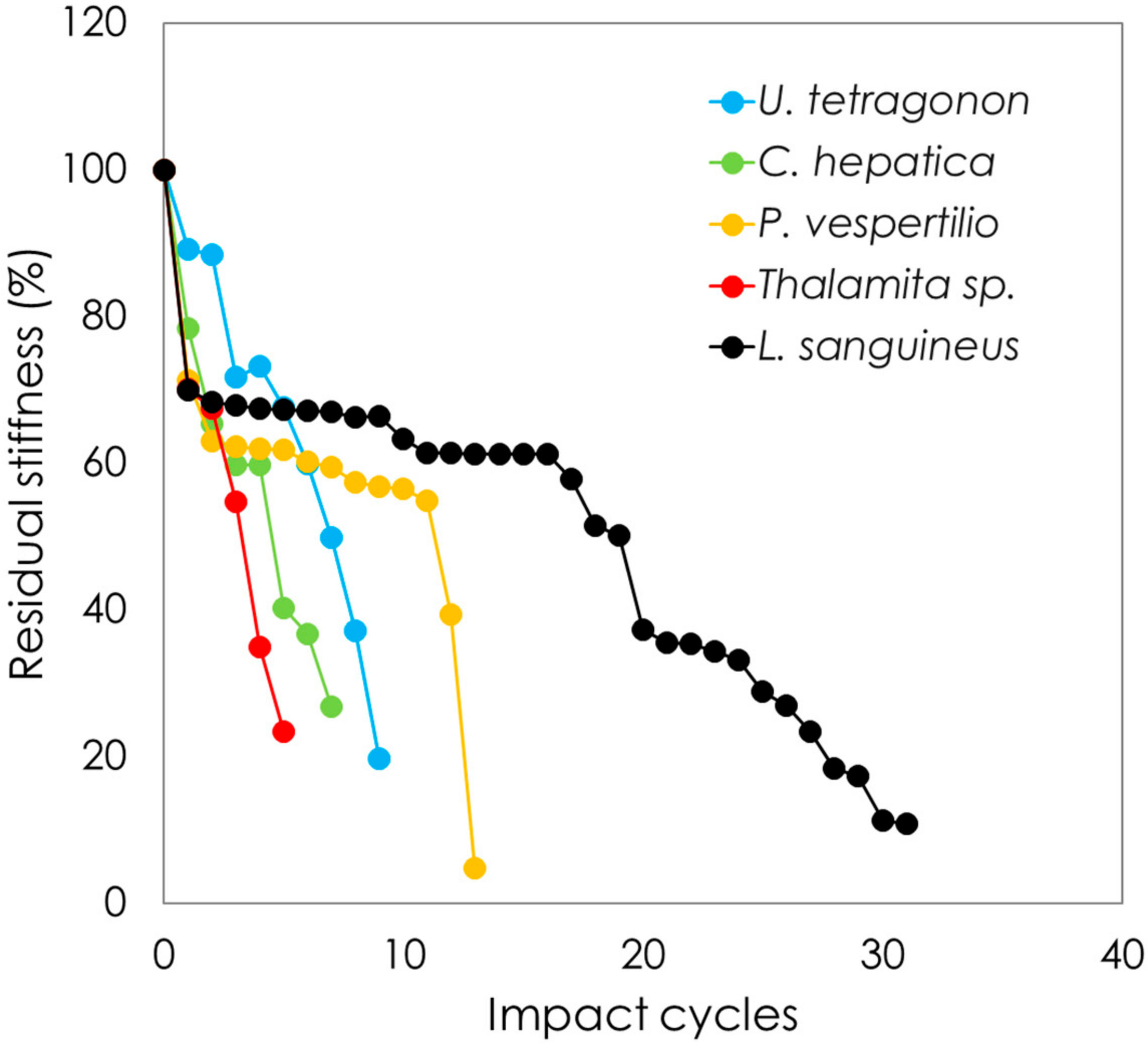
3.3. Cyclic Drop-Weight Impact Testing
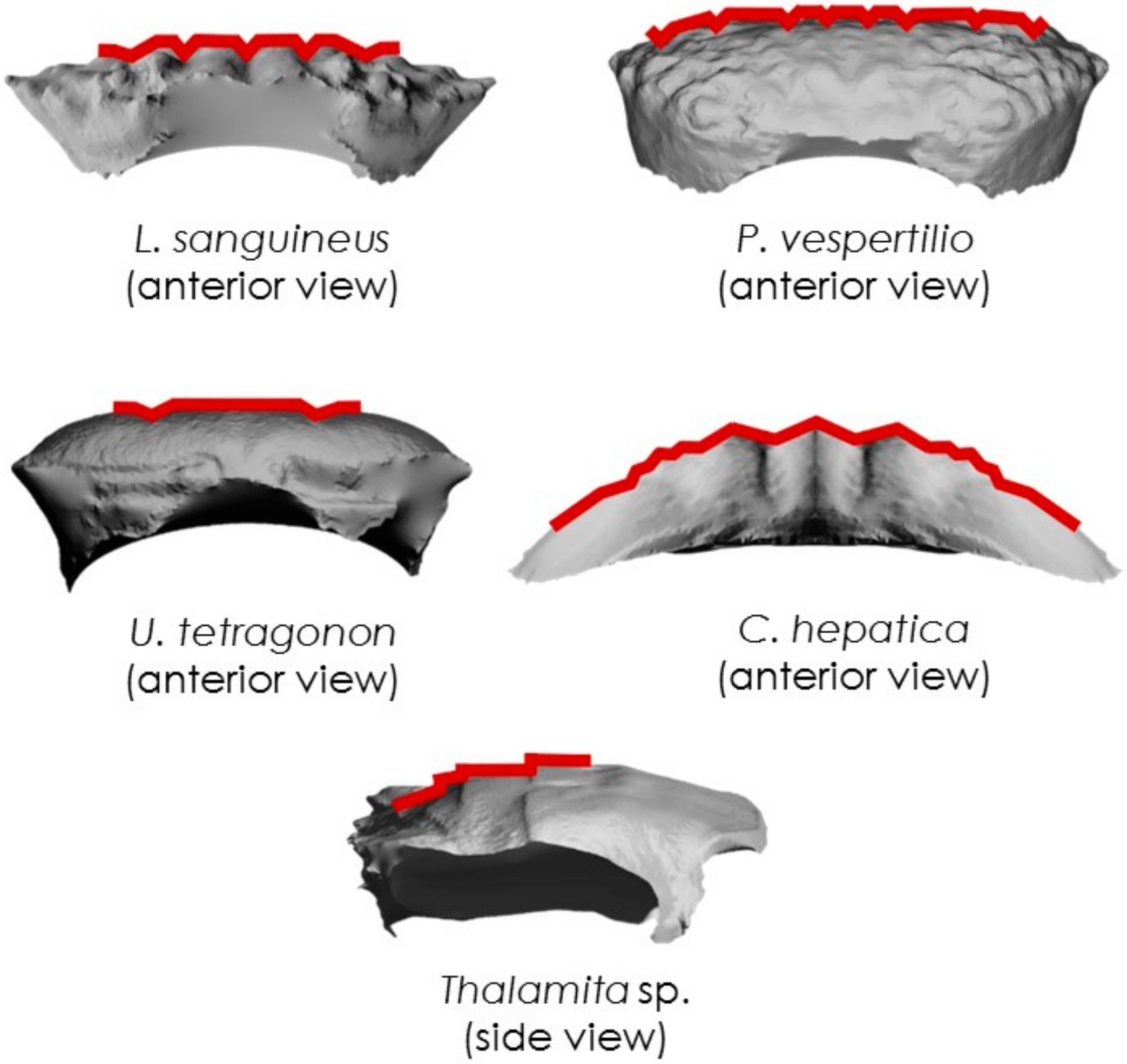
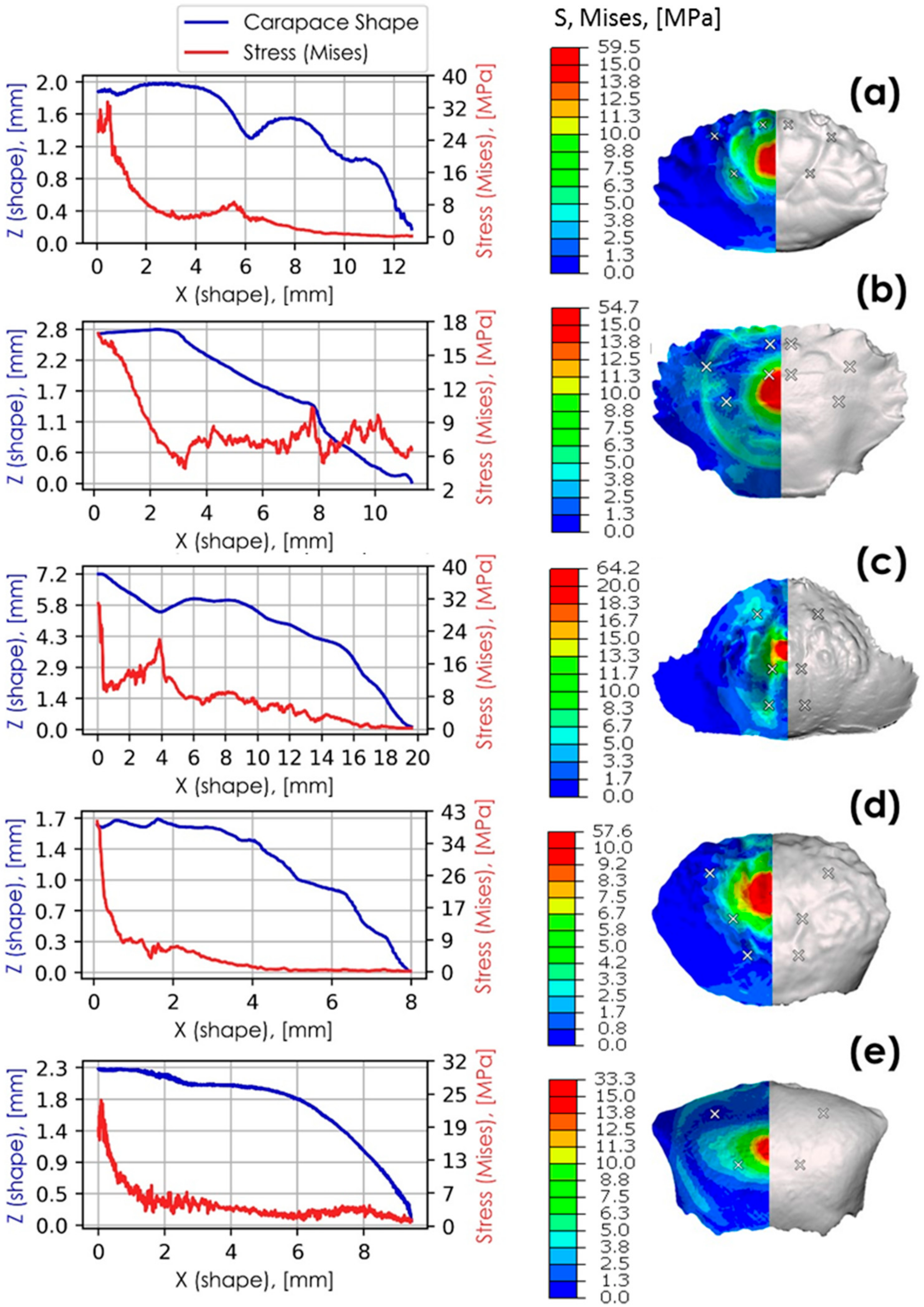
3.4. Finite Element Analyses
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tsang, L.M.; Schubart, C.D.; Ahyong, S.T.; Lai, J.C.Y.; Au, E.Y.C.; Chan, T.; Ng, P.K.L.; Chu, K.H. Evolutionary history of true crabs (Crustacea: Decapoda: Brachyura) and the origin of freshwater crabs. Mol. Biol. Evol. 2014, 31, 1173–1187. [Google Scholar] [CrossRef] [PubMed]
- Garth, J.S.; Abbott, D.P. Brachyura: The True Crabs; Stanford University Press: Stanford, CA, USA, 1980; pp. 592–596. [Google Scholar]
- Hild, S.; Marti, O.; Ziegler, A. Spatial distribution of calcite and amorphous calcium carbonate in the cuticle of the terrestrial crustaceans Porcellio scaber and Armadillidium vulgare. J. Struct. Biol. 2008, 163, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Seidl, B.; Huemer, K.; Neues, F.; Hild, S.; Epple, M.; Ziegler, A. Ultrastructure and mineral distribution in the tergite cuticle of the beach isopod Tylos europaeus Arcangeli, 1938. J. Struct. Biol. 2011, 174, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Hepburn, H.R.; Joffe, I.; Green, N.; Nelson, K.J. Mechanical properties of a crab shell. Comp. Biochem. Phys. A 1975, 50, 551–554. [Google Scholar] [CrossRef]
- Zhou, F.; Wu, Z.; Wang, M.; Chen, K. Structure and mechanical properties of pincers of lobster (Procambarus clarkii) and crab (Eriocheir Sinensis). J. Mech. Behav. Biomed. Mater. 2010, 3, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, X.; Li, J.; Qui, F. Microstructure and Mechanical Properties of the Dactylopodites of the Chinese Mitten Crab (Eriocheir sinensis). Appl. Sci. 2018, 8, 674. [Google Scholar] [CrossRef] [Green Version]
- Alam, P. Structures and Composition of the Crab Carapace: An Archetypal Material in Biomimetic Mechanical Design. In Marine Organisms as Model Systems in Biology and Medicine. Results and Problems in Cell Differentiation; Kloc, M., Kubiak, J., Eds.; Springer-Nature: Cham, Switzerland, 2018; Volume 65, pp. 569–581. [Google Scholar]
- Chen, B.; Yin, D.G.; Yuan, Q.; Fan, J.H. Correlation between Microstructure and Toughness of Crab Carapace. Mater. Sci. Forum 2011, 689, 395–399. [Google Scholar] [CrossRef]
- Chen, B.; Yin, D.G.; Chen, X.; Yuan, Q.; Zhang, Z. Nanostructural mechanism of toughness of crab carapace. J. Comput. Theor. Nanosci. 2013, 10, 1436–1440. [Google Scholar] [CrossRef]
- Chen, P.; Lin, A.Y.; McKittrick, J.; Meyers, M.A. Structure and mechanical properties of crab exoskeletons. Acta Biomater. 2008, 4, 587–596. [Google Scholar] [CrossRef]
- Dutil, J.D.; Rollet, C.; Bouchard, R.; Claxton, W.T. Shell strength and carapace size in non-adult and adult male snow crab (Chionoecetes opilo). J. Crustacean Biol. 2000, 20, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.R.A.; Hebrank, J.; Kier, W.M. Mechanical properties of the rigid and hydrostatic skeletons of molting blue crabs, Callinectes sapidus Rathbun. J. Exp. Biol. 2007, 210, 4272–4278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabritius, H. Correlation of structure, composition and local mechanical properties in the dorsal carapace of the edible crab Cancer pagurus. Z. Krist. Cryst. Mater. 2012, 227, 766–776. [Google Scholar] [CrossRef]
- Urzua, A.; Urbina, M.A. Ecophysiological adaptations to variable salinity environments in the crab Hemigrapsus crenulatus from the Southeastern Pacific coast: Sodium regulation, respiration and excretion. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 210, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.G.; Abello, P.; Kaiser, M.J.; Warman, C.G. Carapace colour, inter-moult duration and the behavioural and physiological ecology of the shore crab Carcinus maenas. Estuar. Coast. Shelf Sci. 1997, 44, 203–211. [Google Scholar] [CrossRef]
- Reaney, L.T.; Backwell, P.R.Y. Risk-taking behavior predicts aggression and mating success in a fiddler crab. Behav. Ecol. 2007, 18, 521–525. [Google Scholar] [CrossRef] [Green Version]
- Bertness, M.D.; Cunningham, C. Crab shell-crushing predation and gastropod architectural defense. J. Exp. Mar. Biol. Ecol. 1981, 50, 213–230. [Google Scholar] [CrossRef]
- Travedi, J.N.; Vachhrajani, K.D. First record of brachyuran crab Leptodius affinis (De Haan, 1835) (Crustacea: Decapoda: Xanthidae) from the western coast of India. Mar. Biodivers. Rec. 2015, 8, 1–5. [Google Scholar] [CrossRef]
- Harvey, H. Predation Risk and Molting Decisions in the Hawaiian Crab Leptodius Sanguineus: The Ups and Downs of An Armored Exoskeleton. Master’s Thesis, Simon Fraser University, Burnaby, BC, Canada, 6 January 1994. [Google Scholar]
- Emmerson, W.D. A Guide to and Checklist for the Decapoda of Namibia, South Africa and Mozambique; Cambridge Scholars Publishing: Cambridge, UK, 2016; Volume 3, p. 138. [Google Scholar]
- Fahimi, N.; Science, M.; Seyfabadi, J.; Sari, A. Size at sexual maturity, breeding season, and fecundity of the intertidal xanthid crab Leptodius exaratus (H. Milne Edwards, 1834) (Decapoda: Brachyura) in the Persian Gulf, Iran. J. Crustacean Biol. 2017, 37, 465–472. [Google Scholar] [CrossRef]
- Sung, J.; Lee, J.; Kim, S.; Karagozlu, M.Z.; Kim, C. Complete mitochondrial genome of Leptodius sanguineus (Decapoda, Xanthidae). Mitochondrial DNA B 2016, 1, 500–501. [Google Scholar] [CrossRef]
- Kumar, A.A.J.; Wesley, S.G. Spatio-temporal distribution of assemblages of Brachyuran crabs at Laamu Atoll, Maldives. Atoll Res. Bull. 2010, 588, 1–18. [Google Scholar] [CrossRef]
- Cannicci, S.; Dahdouh-Guebas, F.; Anyona, D.; Vannini, M. Natural diet and feeding habits of Thalamita crenata (Decapoda: Portunidae). J. Crustacean Biol. 1996, 16, 678–683. [Google Scholar] [CrossRef]
- Vezzosi, R.; Barbaresi, S.; Anyona, D. Marine behaviour and physiology activity patterns in Thalamita crenata (Portunidae, Decapoda): A shaping by the tidal cycles. Mar. Behav. Physiol. 2015, 24, 37–41. [Google Scholar] [CrossRef]
- Takeda, M.; Rahayu, D.L.; Aswandy, I. Prawns and crabs. In Field Guide to Lombok Island: Identification Guide to Marine Organisms in Seagrass Beds of Lombok Island, Indonesia; Matsuura, K., Sumadhiharga, O.K., Tsukamoto, K., Eds.; Ocean Research Institute University of Tokyo: Tokyo, Japan, 2000; p. 44. [Google Scholar]
- Sanka, I.; Suyono, E.A.; Rivero-Muller, A.; Alam, P. Carapace surface architecture facilitates camouflage of the decorator crab Tiarinia cornigera. Acta Biomater. 2016, 41, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Hogarth, P.J. The Biology of Mangroves and Seagrasses, 2nd ed.; Oxford University Press: New York, NY, USA, 2007; pp. 114–118. [Google Scholar]
- Mokhtari, M.; Ghaffar, M.A.; Usup, G.; Cob, Z.C. The ecology of fiddler crab Uca forcipata in mangrove forest. AIP Conf. Proc. 2013, 1571, 498–504. [Google Scholar] [CrossRef]
- Poupin, J.; Juncker, M. A Guide to Decapod Crustaceans of the South Pacific; SPC FAME Digital Library: Noumea, France, 2010; p. 320. [Google Scholar]
- Carpenter, K.E.; Niem, V.H. The Living Marine Resources of the Western Central Pacific Volume 2 Cephalopods, Crustaceans, Holothurians and Sharks (FAO Species Identification Guide for Fishery Purposes); FAO: Rome, Italy, 1998; pp. 687–1396. [Google Scholar]
- Hopkins, M.J.; Thurman, C.L. The geographic structure of morphological variation in eight species of fiddler crabs (Ocypodidae: Genus Uca) from the eastern United States and Mexico. Biol. J. Linn. Soc. 2010, 100, 248–270. [Google Scholar] [CrossRef] [Green Version]
- Suthamrit, W.; Thaewnon-Ngiw, B. Morphometry of mountain crabs (Crustacea: Decapoda: Brachyura: Potamidae) from Phetchabun Mountains Thailand. Interdiscip. Res. Rev. 2020, 15, 24–31. [Google Scholar]
- Silva, A.C.; Silva, I.C.; Hawkins, S.J.; Boaventura, D.M.; Thompson, R.C. Cheliped morphological variation of the intertidal crab Eriphia verrucosa across shores of differing exposure to wave action. J. Exp. Mar. Biol. Ecol. 2010, 391, 84–91. [Google Scholar] [CrossRef]
- Campbell, N.A.; Mahon, R.J. A multivariate study of variation in two species of rock crab of the genus Leptograpsus. Aust. J. Zool. 1974, 22, 418–425. [Google Scholar] [CrossRef]
- Kazemahvazi, S.; Tanner, D.; Zenkert, D. Corrugated all-composite sandwich structures. Part 2: Failure mechanisms and experimental programme. Compos. Sci. Technol. 2008, 69, 920–925. [Google Scholar] [CrossRef] [Green Version]
- Qin, Q.H.; Zhang, J.X.; Wang, T.J. Low velocity impact response of lightweight metal sandwich panel with corrugated core. Mater. Res. Innov. 2011, 15, s198–s200. [Google Scholar] [CrossRef]
- Li, X.W.; Ji, H.M.; Yang, W.; Zhang, G.P.; Chen, D.L. Mechanical properties of crossed-lamellar structures in biological shells: A review. J. Mech. Behav. Biomed. Mater. 2017, 74, 54–71. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | No of Elements | Element Type | Fine Edge Mesh Size (mm) | Coarse Edge Mesh Size (mm) |
|---|---|---|---|---|
| L. sanguineus | 60,618 | S3R | 0.12 | 0.27 |
| P. vespertilio | 53,849 | S3R | 0.08 | 0.2 |
| U. tetragonon | 55,132 | S3R | 0.07 | 0.2 |
| C. hepatica | 75,490 | S3R | 0.12 | 0.63 |
| T. danae | 59,741 | S3R | 0.12 | 0.27 |
| Species | Morphologies |
|---|---|
| L. sanguineus | The carapace is wider than it is long and is anteriorly convex. Markings no the dorsal face of the carapace surface are unclear. The antero-lateral margins have six teeth. The chelae fingers are spoon-like in shape, and the chelae fingers are black in colour. |
| T. danae & T. prymna | The carapace is transversely ovate with five antero-lateral teeth. The fronto-orbital margin is very wide. T. danae and T. prymna are of a very similar morphology. The main distinction is the basal antennal segment joints of T. prymna, which are without spines, unlike in T. danae. |
| P. vespertilio | The carapace is marginally little longer than it is wide. The antero-lateral margin of the carapace has three teeth. The carapace is covered in, as are the chelae and legs. The lower half of the chela (outside face) is covered by bead-like granules. |
| C. hepatica | Posterolateral parts of the carapace with clypeiform expansion covers the crab’s legs. Its margin has teeth, laterally the margin is dentate. The anterior edge of this clypeiform structure is gently denticulate. The right (larger) chela exhibits a specialised cutting tooth. |
| U. tetragonon | The frontal region of the carapace is narrower at its base than at its tip. Sexual dimorphism between male and female chelae is obvious since the outer face of male (larger) chela is granulated. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayekti, P.R.; Fahrunnida; Cerniauskas, G.; Robert, C.; Retnoaji, B.; Alam, P. The Impact Behaviour of Crab Carapaces in Relation to Morphology. Materials 2020, 13, 3994. https://doi.org/10.3390/ma13183994
Sayekti PR, Fahrunnida, Cerniauskas G, Robert C, Retnoaji B, Alam P. The Impact Behaviour of Crab Carapaces in Relation to Morphology. Materials. 2020; 13(18):3994. https://doi.org/10.3390/ma13183994
Chicago/Turabian StyleSayekti, Puspa Restu, Fahrunnida, Gabrielis Cerniauskas, Colin Robert, Bambang Retnoaji, and Parvez Alam. 2020. "The Impact Behaviour of Crab Carapaces in Relation to Morphology" Materials 13, no. 18: 3994. https://doi.org/10.3390/ma13183994
APA StyleSayekti, P. R., Fahrunnida, Cerniauskas, G., Robert, C., Retnoaji, B., & Alam, P. (2020). The Impact Behaviour of Crab Carapaces in Relation to Morphology. Materials, 13(18), 3994. https://doi.org/10.3390/ma13183994