Mapping Single Walled Carbon Nanotubes in Photosynthetic Algae by Single-Cell Confocal Raman Microscopy

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of SWCNTs
2.2. Algal Growth and Treatment
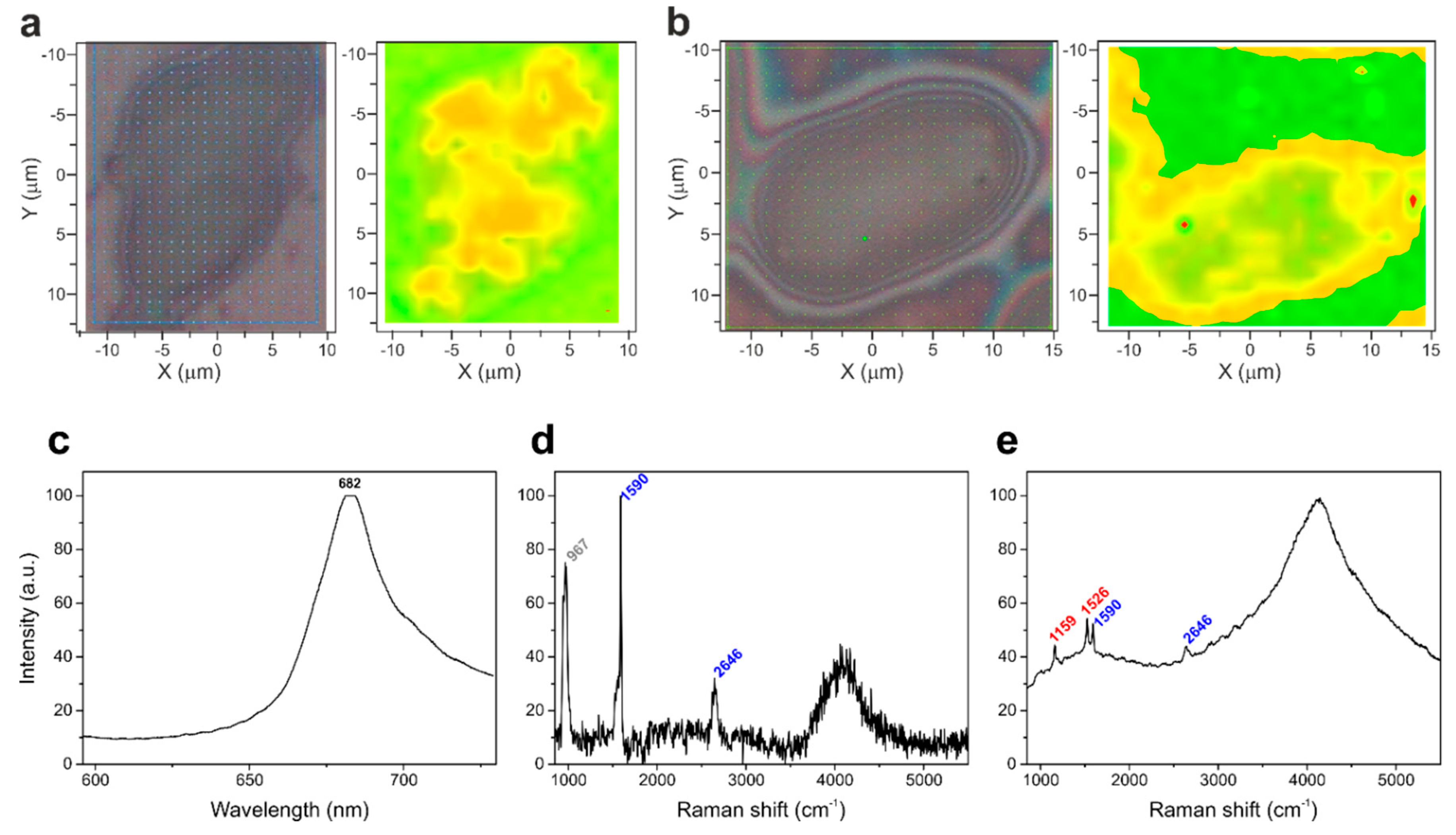
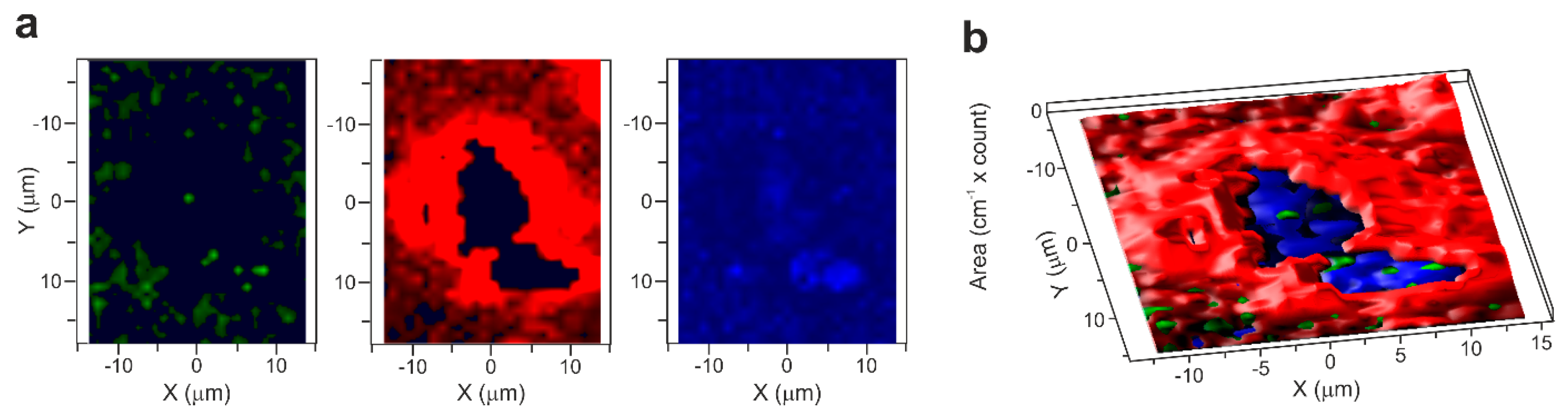
2.3. Raman Microscopy
2.4. Scanning Electron Microscopy
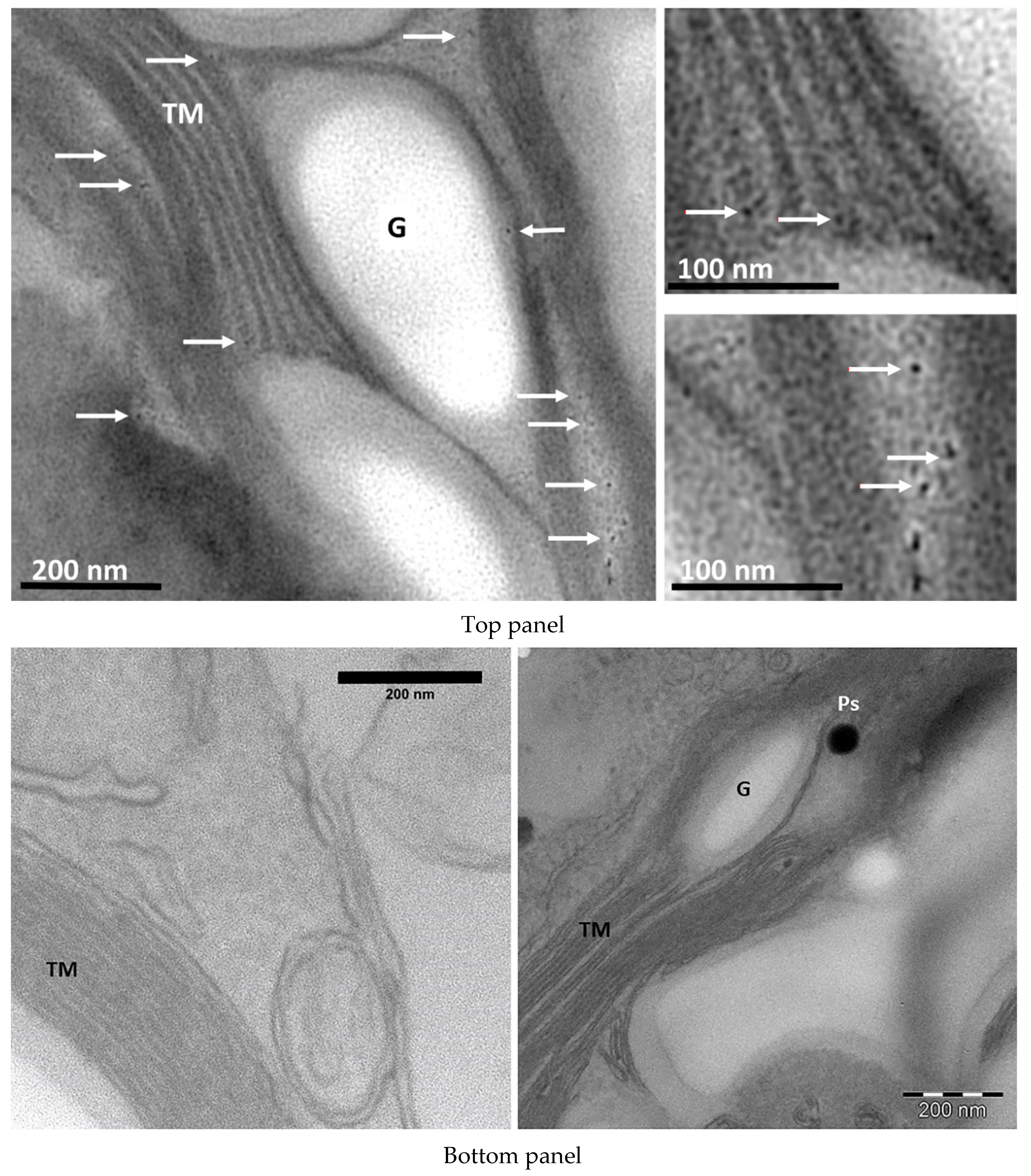
2.5. Transmission Electron Microscopy
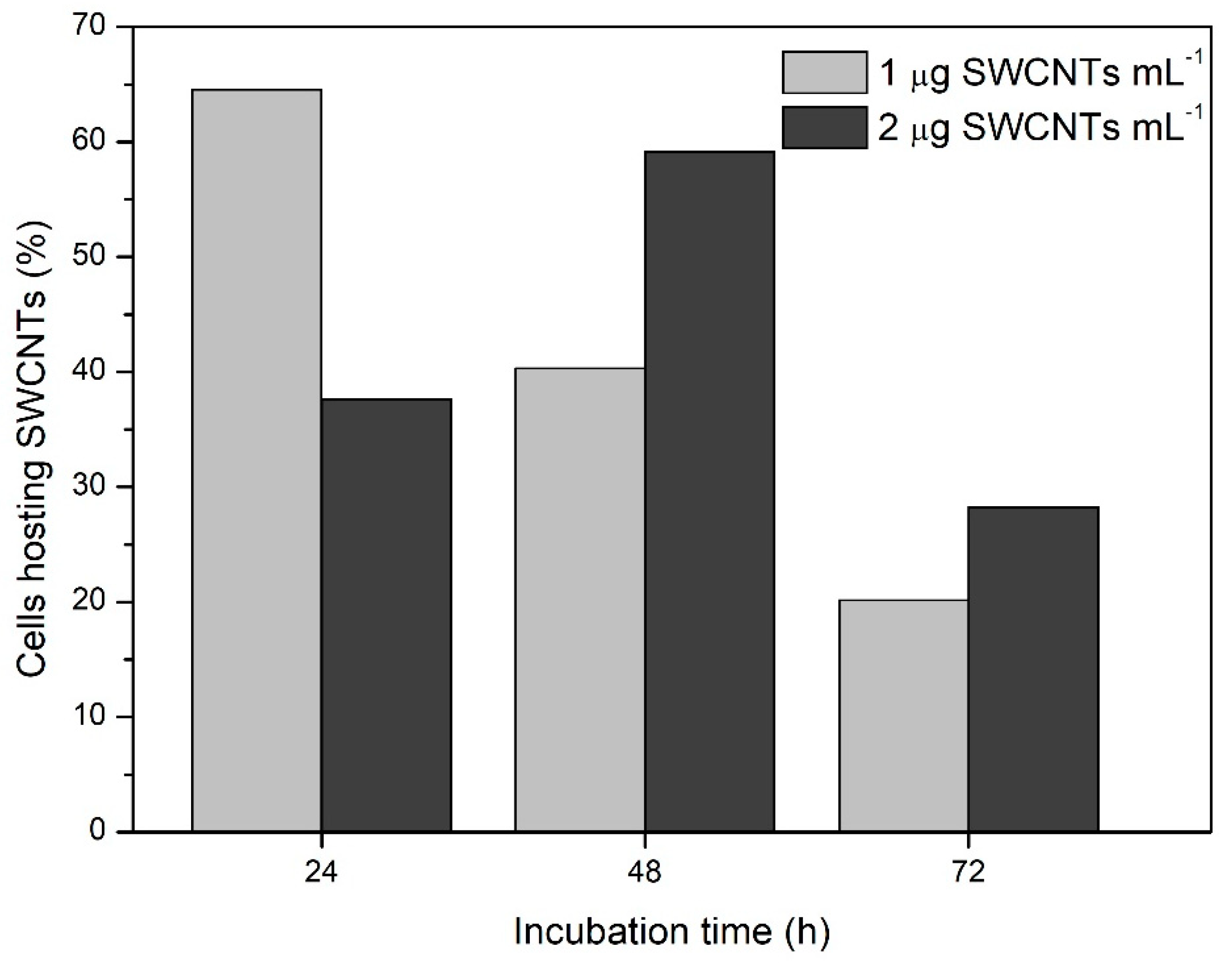
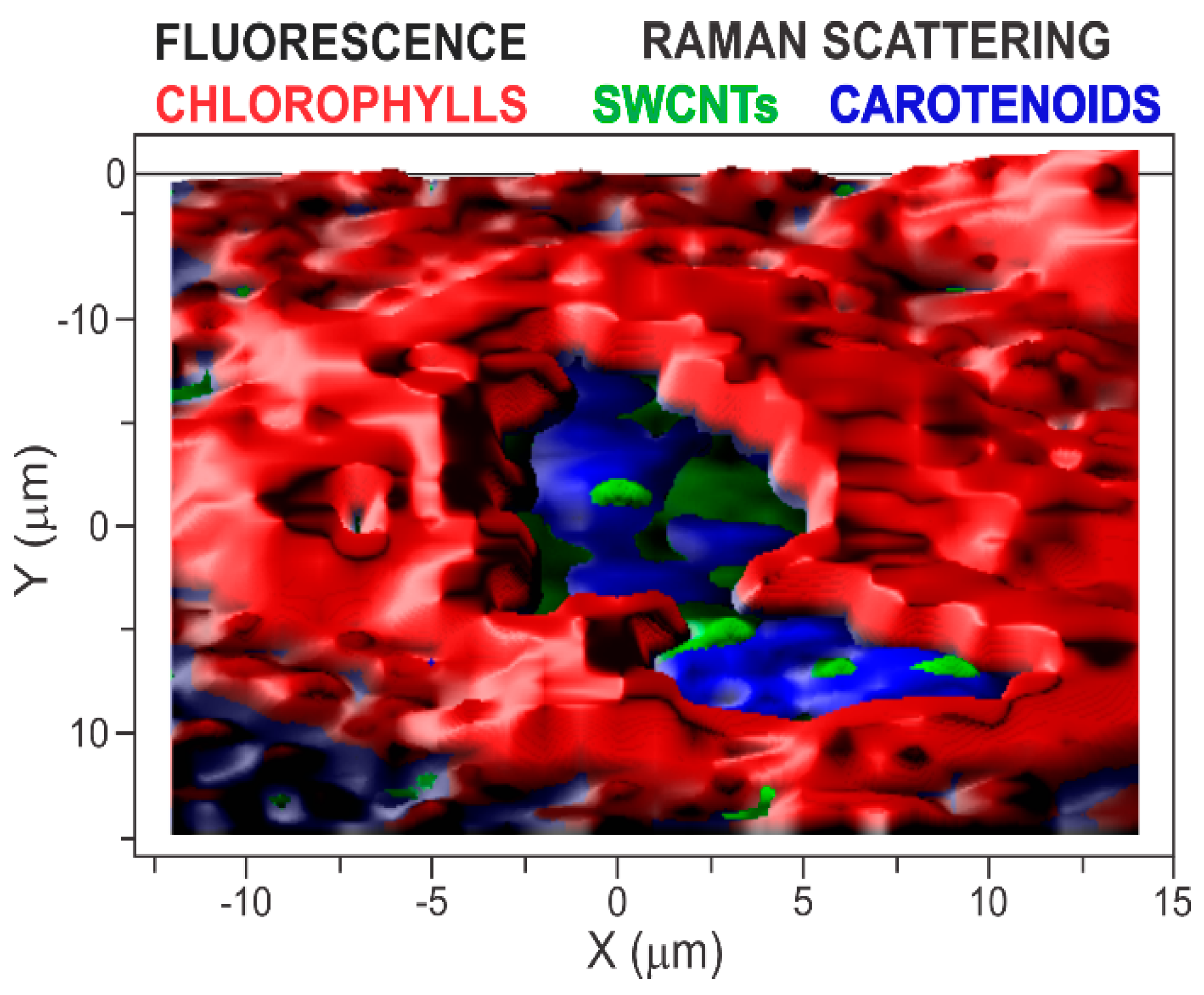
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Nair, R.; Varghese, S.H.; Nair, B.G.; Maekawa, T.; Yoshida, Y.; Kumar, D.S. Nanoparticulate material delivery to plants. Plant Sci. 2010, 179, 154–163. [Google Scholar] [CrossRef]
- Solanki, P.; Bhargava, A.; Chhipa, H.; Jain, N.; Panwar, J. Nano-fertilizers and Their Smart Delivery System. In Nanotechnologies in Food and Agriculture; Springer International Publishing: Cham, Switzerland, 2015; pp. 81–101. [Google Scholar] [CrossRef]
- Guatimosim, S.; Ladeira, M.; Rocha-Resende, C.; Resende, R.R. The Use of Single Wall Carbon Nanotubes as a Delivery System for siRNA; Springer: Cham, Switzerland, 2016; pp. 17–29. [Google Scholar] [CrossRef]
- Seabra, A.B.; Rai, M.; Durán, N.J. Nano carriers for nitric oxide delivery and its potential applications in plant physiological process: A mini review. Plant Biochem. Biotechnol. 2014, 23, 1–10. [Google Scholar] [CrossRef]
- Wang, P.; Lombi, E.; Zhao, F.-J.; Kopittke, P.M. Nanotechnology: A New Opportunity in Plant Sciences. Trends Plant Sci. 2016, 21, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, J.P.; Landry, M.P.; Faltermeier, S.M.; McNicholas, T.P.; Iverson, N.M.; Boghossian, A.A.; Reuel, N.F.; Hilmer, A.J.; Sen, F.; Brew, J.A.; et al. Plant nanobionics approach to augment photosynthesis and biochemical sensing. Nat. Mater. 2014, 13, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Boghossian, A.a.; Sen, F.; Gibbons, B.M.; Sen, S.; Faltermeier, S.M.; Giraldo, J.P.; Zhang, C.T.; Zhang, J.; Heller, D.a.; Strano, M.S. Application of nanoparticle antioxidants to enable hyperstable chloroplasts for solar energy harvesting. Adv. Energy Mater. 2013, 3, 881–893. [Google Scholar] [CrossRef]
- Serag, M.F.; Kaji, N.; Habuchi, S.; Bianco, A.; Baba, Y. Nanobiotechnology meets plant cell biology: Carbon nanotubes as organelle targeting nanocarriers. RSC Adv. 2013, 3, 4856. [Google Scholar] [CrossRef]
- Giraldo, J.P.; Landry, M.P.; Kwak, S.Y.; Jain, R.M.; Wong, M.H.; Iverson, N.M.; Ben-Naim, M.; Strano, M.S. A Ratiometric Sensor Using Single Chirality Near-Infrared Fluorescent Carbon Nanotubes: Application to in Vivo Monitoring. Small 2015, 11, 3973–3984. [Google Scholar] [CrossRef] [Green Version]
- Bisker, G.; Dong, J.; Park, H.D.; Iverson, N.M.; Ahn, J.; Nelson, J.T.; Landry, M.P.; Kruss, S.; Strano, M.S. Protein-targeted corona phase molecular recognition. Nat. Commun. 2016, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lambreva, M.D.; Lavecchia, T.; Tyystjärvi, E.; Antal, T.K.; Orlanducci, S.; Margonelli, A.; Rea, G. Potential of carbon nanotubes in algal biotechnology. Photosynth Res. 2015, 125, 451–471. [Google Scholar] [CrossRef]
- Serag, M.F.; Braeckmans, K.; Habuchi, S.; Kaji, N.; Bianco, A.; Baba, Y. Spatiotemporal visualization of subcellular dynamics of carbon nanotubes. Nano Lett. 2012, 12, 6145–6151. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Chen, B.; Wang, Q.; Shi, X.; Xiao, Z.; Lin, J.; Fang, X. Carbon nanotubes as molecular transporters for walled plant cells. Nano Lett. 2009, 9, 1007–1010. [Google Scholar] [CrossRef]
- Serag, M.F.; Kaji, N.; Venturelli, E.; Okamoto, Y.; Terasaka, K.; Tokeshi, M.; Mizukami, H.; Braeckmans, K.; Bianco, A.; Baba, Y. Functional platform for controlled subcellular distribution of carbon nanotubes. ACS Nano 2011, 5, 9264–9270. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.H.; Misra, R.P.; Giraldo, J.P.; Kwak, S.Y.; Son, Y.; Landry, M.P.; Swan, J.W.; Blankschtein, D.; Strano, M.S. Lipid exchange envelope penetration (leep) of nanoparticles for plant engineering: A universal localization mechanism. Nano Lett. 2016, 16, 1161–1172. [Google Scholar] [CrossRef] [Green Version]
- Jorio, A.; Saito, R.; Dresselhaus, G.; Dresselhaus, M.S. Raman Spectroscopy in Graphene Related Systems; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2011. [Google Scholar] [CrossRef]
- Lamprecht, C.; Gierlinger, N.; Heister, E.; Unterauer, B.; Plochberger, B.; Brameshuber, M.; Hinterdorfer, P.; Hild, S.; Ebner, A. Mapping the intracellular distribution of carbon nanotubes after targeted delivery to carcinoma cells using confocal Raman imaging as a label-free technique. J. Phys. Condens. Matter. 2012, 24. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Luo, F.; Li, J.; Zhu, B.; Wang, G.-X. Biocompatibility assessment of single-walled carbon nanotubes using Saccharomyces cerevisiae as a model organism. J. Nanobiotechnol. 2018, 16, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Ouyang, S.; Mu, L.; An, J.; Zhou, Q. Effects of Graphene Oxide and Oxidized Carbon Nanotubes on the Cellular Division, Microstructure, Uptake, Oxidative Stress, and Metabolic Profiles. Environ. Sci. Technol. 2015, 49, 10825–10833. [Google Scholar] [CrossRef]
- Moudříková, Š.; Mojzeš, P.; Zachleder, V.; Pfaff, C.; Behrendt, D.; Nedbal, L. Raman and fluorescence microscopy sensing energy-transducing and energy-storing structures in microalgae. Algal Res. 2016, 16, 224–232. [Google Scholar] [CrossRef]
- Hollricher, O.; Ibach, W. High-Resolution Optical and Confocal Microscopy. In Confocal Raman Microscopy; Dieing, T., Hollricher, O., Toporski, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1–11. [Google Scholar] [CrossRef] [Green Version]
- Jehlička, J.; Edwards, H.G.M.; Oren, A. Raman spectroscopy of microbial pigments. Appl. Environ. Microbiol. 2014, 80, 3286–3295. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Yin, H.; Huang, W.E. Raman activated cell sorting. Curr. Opin. Chem. Biol. 2016, 33, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Moudříková, Š.; Nedbal, L.; Solovchenko, A.; Mojzeš, P. Raman microscopy shows that nitrogen-rich cellular inclusions in microalgae are microcrystalline guanine. Algal Res. 2017, 23, 216–222. [Google Scholar] [CrossRef]
- Butler, H.J.; Ashton, L.; Bird, B.; Cinque, G.; Curtis, K.; Dorney, J.; Esmonde-White, K.; Fullwood, N.J.; Gardner, B.; Martin-Hirsch, P.L.; et al. Using Raman spectroscopy to characterize biological materials. Nat. Protoc. 2016, 11, 664–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Chiu, L.; Ho, S.H.; Shimada, R.; Ren, N.Q.; Ozawa, T. Rapid in vivo lipid/carbohydrate quantification of single microalgal cell by Raman spectral imaging to reveal salinity-induced starch-to-lipid shift. Biotechnol. Biofuels 2017, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Jie, D.; Cuello, J.J.; Johnson, D.J.; Qiu, Z.; He, Y. Microalgal detection by Raman microspectroscopy. Trends Anal. Chem. 2014, 53, 33–40. [Google Scholar] [CrossRef]
- Valentini, F.; Amine, A.; Orlanducci, S.; Terranova, M.L.; Palleschi, G. Carbon Nanotube Purification: Preparation and Characterization of Carbon Nanotube Paste Electrodes. Anal. Chem. 2003, 75, 5413–5421. [Google Scholar] [CrossRef]
- Zheng, M.; Jagota, A.; Semke, E.D.; Diner, B.A.; Mclean, R.S.; Lustig, S.R.; Richardson, R.E.; Tassi, N.G. DNA-assisted dispersion and separation of carbon nanotubes. Nat. Mater. 2003, 2, 338–342. [Google Scholar] [CrossRef]
- Kupis-Rozmysłowicz, J.; Antonucci, A.; Boghossian, A.A. Review–Engineering the selectivity of the DNA-SWCNT sensor. ECS J. Solid State Sci. Technol. 2016, 5, M3067–M3074. [Google Scholar] [CrossRef]
- Landry, M.P.; Vuković, L.; Kruss, S.; Bisker, G.; Landry, A.M.; Islam, S.; Jain, R.; Schulten, K.; Strano, M.S. Comparative dynamics and sequence dependence of DNA and RNA binding to single walled carbon nanotubes. J. Phys. Chem. C 2015, 119, 10048–10058. [Google Scholar] [CrossRef] [Green Version]
- Davies, D.R.; Plaskitt, A. Genetical and structural analyses of cell-wall formation in Chlamydomonas reinhardi. Genet. Res. 1971, 17, 33–43. [Google Scholar] [CrossRef]
- Voigt, J.; Hinkelmann, B.; Harris, E.H. Production of cell wall polypeptides by different cell wall mutants of the unicellular green alga Chlamydomonas reinhardtii. Microbiol. Res. 1997, 152, 189–198. [Google Scholar] [CrossRef]
- Rasband, W.S.; Image, J.U.S. National Institutes of Health, Bethesda, Maryland, USA. Available online: https://imagej.nih.gov/ij/ (accessed on 28 November 2018).
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Sprotocols, Chlamydomonas Fixation for Transmission Electron Microscopy. Available online: https://protocols.scienceexchange.com/protocols/chlamydomonas-fixation-for-transmission-electron-microscopy (accessed on 28 November 2018).
- Yang, C.; Mamouni, J.; Tang, Y.; Yang, L. Antimicrobial activity of single-walled carbon nanotubes: Length effect. Langmuir 2010, 26, 16013–16019. [Google Scholar] [CrossRef] [PubMed]
- Engel, B.D.; Schaffer, M.; Cuellar, L.K.; Villa, E.; Plitzko, J.M.; Baumeister, W. Native architecture of the Chlamydomonas chloroplast revealed by in situ cryo-electron tomography. Elife 2015. [Google Scholar] [CrossRef]
- Davies, D.R.; Plaskitt, A. Cell wall organisation in Chlamydomonas reinhardi. Genet. Res. 1971, 17, 33–43. [Google Scholar] [CrossRef]
- Shafer-Peltier, K.E.; Haka, A.S.; Motz, J.T.; Fitzmaurice, M.; Dasari, R.R.; Feld, M.S. Model-based biological Raman spectral imaging. J. Cell. Biochem. 2002, 87, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Pedrós, R.; Moya, I.; Goulas, Y.; Jacquemoud, S. Chlorophyll fluorescence emission spectrum inside a leaf. Photochem. Photobiol. Sci. 2008, 7, 498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moudříková, Š.; Sadowsky, A.; Metzger, S.; Nedbal, L.; Mettler-Altmann, T.; Mojzeš, P. Quantification of Polyphosphate in Microalgae by Raman Microscopy and by a Reference Enzymatic Assay. Anal. Chem. 2017, 89, 12006–12013. [Google Scholar] [CrossRef]
- Sharma, B.; Frontiera, R.R.; Henry, A.I.; Ringe, E.; van Duyne, R.P. SERS: Materials, applications, and the future. Mater. Today 2012, 15, 16–25. [Google Scholar] [CrossRef]
- Kagan, M.R.; McCreery, R.L. Reduction of Fluorescence Interference in Raman Spectroscopy via Analyte Adsorption on Graphitic Carbon. Anal. Chem. 1994, 66, 4159–4165. [Google Scholar] [CrossRef]
- Ling, X.; Huang, S.; Deng, S.; Mao, N.; Kong, J.; Dresselhaus, M.S.; Zhang, J. Lighting Up the Raman Signal of Molecules in the Vicinity of Graphene Related Materials. Acc. Chem. Res. 2015, 48, 1862–1870. [Google Scholar] [CrossRef]
- Zhu, Z.; Yang, R.; You, M.; Zhang, X.; Wu, Y.; Tan, W. Single-walled carbon nanotube as an effective quencher. Anal. Bioanal. Chem. 2010, 396, 73–83. [Google Scholar] [CrossRef]
- Casey, J.P.; Bachilo, S.M.; Weisman, R.B. Efficient photosensitized energy transfer and near-ir fluorescence from porphyrin-swnt complexes. J. Mater. Chem. 2008, 18, 1510–1516. [Google Scholar] [CrossRef]
- Chitta, R.; Sandanayaka, A.S.D.; Schumacher, A.L.; D’Souza, L.; Araki, Y.; Ito, O.; D’Souza, F.J. Donor–Acceptor nanohybrids of zinc naphthalocyanine or zinc porphyrin noncovalently linked to single-wall carbon nanotubes for photoinduced electron transfer. Phys. Chem. C 2007, 111, 6947–6955. [Google Scholar] [CrossRef]
- Lin, S.; Keskar, G.; Wu, Y.; Wang, X.; Mount, A.S.; Klaine, S.J.; Moore, J.M.; Rao, A.M.; Ke, P.C. Detection of phospholipid-carbon nanotube translocation using fluorescence energy transfer. Appl. Phys. Lett. 2006, 89, 143118. [Google Scholar] [CrossRef]
- Chiu, C.F.; Dementev, N.; Borguet, E.J. Fluorescence Quenching of Dyes Covalently Attached to Single-Walled Carbon Nanotubes. Phys. Chem. A 2011, 9579–9584. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Cui, D.; Ozkan, C.S.; Ozkan, M.; Xu, P.; Huang, T.; Liu, F.; Chen, H.; Li, Q.; He, R.; et al. Effects of carbon nanotubes on photoluminescence properties of quantum dots. Phys. Chem. C 2008, 112, 939–944. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, W.; Jia, M.; Wei, L.; Fan, X.-F.; Kuo, J.-L.; Chen, Y.; Chan-Park, M.B.; Xia, A.; Li, L.-J. Energy Transfer from Photo-Excited Fluorene Polymers to Single-Walled Carbon Nanotubes. J. Phys. Chem. C 2009, 113, 14946–14952. [Google Scholar] [CrossRef]
- Stoll, M.; Rafailov, P.M.; Frenzel, W.; Thomsen, C. Electrochemical and Raman measurements on single-walled carbon nanotubes. Chem. Phys. Lett. 2003, 375, 625–631. [Google Scholar] [CrossRef]
- Gupta, S.; Hughes, M.; Windle, A.H.; Robertson, J.J. Charge transfer in carbon nanotube actuators investigated using in situ Raman spectroscopy. Appl. Phys. 2004, 95, 2038–2048. [Google Scholar] [CrossRef]
- King, A.a.K.; Hanus, M.J.; Harris, A.T.; Minett, A.I. Nanocarbon-chlorophyll hybrids: Self assembly and photoresponse. Carbon N. Y. 2014, 80, 746–754. [Google Scholar] [CrossRef]
- Dorogi, M.; Bálint, Z.; Mikó, C.; Vileno, B.; Milas, M.; Hernádi, K.; László, F.; Váró, G.; Nagy, L. Stabilization effect of single-walled carbon nanotubes on the functioning of photosynthetic reaction centers. J. Phys. Chem. B 2006, 110, 21473–21479. [Google Scholar] [CrossRef]
- Magyar, M.; Hajdu, K.; Szabò, T.; Hernàdi, K.; Dombi, A.; Horvàth, E.; Magrez, A.; Forrò, L.; Nagy, L. Long term stabilization of reaction center protein photochemistry by carbon nanotubes. Phys. Status Solidi Basic Res. 2011, 248, 2454–2457. [Google Scholar] [CrossRef]
- Dewi, H.A.; Sun, G.; Zheng, L.; Lim, S. Interaction and charge transfer between isolated thylakoids and multi-walled carbon nanotubes. Phys. Chem. Chem. Phys. 2015, 17, 3435–3440. [Google Scholar] [CrossRef] [PubMed]
- Wiwatowski, K.; Duzyńska, A.; Świniarski, M.; Szalkowski, M.; Zdrojek, M.; Judek, J.; Mackowski, S.; Kaminska, I. Energy transfer from natural photosynthetic complexes to single-wall carbon nanotubes. J. Lumin. 2016, 170, 855–859. [Google Scholar] [CrossRef]
- Maslenkova, L.T.; Peeva, V.N.; Markovska, Y.K.; Zeinalov, Y. Thermoluminescence Study of Photosystem II Activity in Resurrection Plant Haberlea rhodopensis during Desiccation. In Handbook of Plant and Crop Stress, 3rd ed.; Pessarakli, M., Ed.; CRC Press, Taylor&Francis Group: Boca Raton, FL, USA; London, UK; New York, NY, USA, 2010; pp. 435–446. [Google Scholar] [CrossRef]
- Holzinger, A.; Karsten, U. Desiccation stress and tolerance in green algae: Consequences for ultrastructure, physiological and molecular mechanisms. Front. Plant Sci. 2013, 4, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, T.; Sugiura, M. Flash-induced FTIR difference spectra of the water oxidizing complex in moderately hydrated photosystem II core films: Effect of hydration extent on S-state transitions. Biochemistry 2002, 41, 2322–2330. [Google Scholar] [CrossRef]
- Butler, W.L. On the primary nature of fluorescence yield changes associated with photosynthesis. Proc. Natl. Acad. Sci. USA 1972, 69, 3420–3422. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SWCNTs | Length (±SD), nm |
|---|---|
| Pristine SWCNTs | Average 800 |
| (refer to product specifications) | (range 300–2300) |
| Purified unshortened SWCNTs | 800 ± 200 |
| Purified shortened SWCNTs tip sonication applied for 15 min | 710 ± 100 |
| tip sonication applied for 30 min | 700 ± 110 |
| tip sonication applied for 60 min | 650 ± 150 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orlanducci, S.; Fulgenzi, G.; Margonelli, A.; Rea, G.; Antal, T.K.; Lambreva, M.D. Mapping Single Walled Carbon Nanotubes in Photosynthetic Algae by Single-Cell Confocal Raman Microscopy. Materials 2020, 13, 5121. https://doi.org/10.3390/ma13225121
Orlanducci S, Fulgenzi G, Margonelli A, Rea G, Antal TK, Lambreva MD. Mapping Single Walled Carbon Nanotubes in Photosynthetic Algae by Single-Cell Confocal Raman Microscopy. Materials. 2020; 13(22):5121. https://doi.org/10.3390/ma13225121
Chicago/Turabian StyleOrlanducci, Silvia, Gianluca Fulgenzi, Andrea Margonelli, Giuseppina Rea, Taras K. Antal, and Maya D. Lambreva. 2020. "Mapping Single Walled Carbon Nanotubes in Photosynthetic Algae by Single-Cell Confocal Raman Microscopy" Materials 13, no. 22: 5121. https://doi.org/10.3390/ma13225121
APA StyleOrlanducci, S., Fulgenzi, G., Margonelli, A., Rea, G., Antal, T. K., & Lambreva, M. D. (2020). Mapping Single Walled Carbon Nanotubes in Photosynthetic Algae by Single-Cell Confocal Raman Microscopy. Materials, 13(22), 5121. https://doi.org/10.3390/ma13225121