The Impact of Zinc Oxide Nanoparticles on Male (In)Fertility




Abstract
:1. Introduction
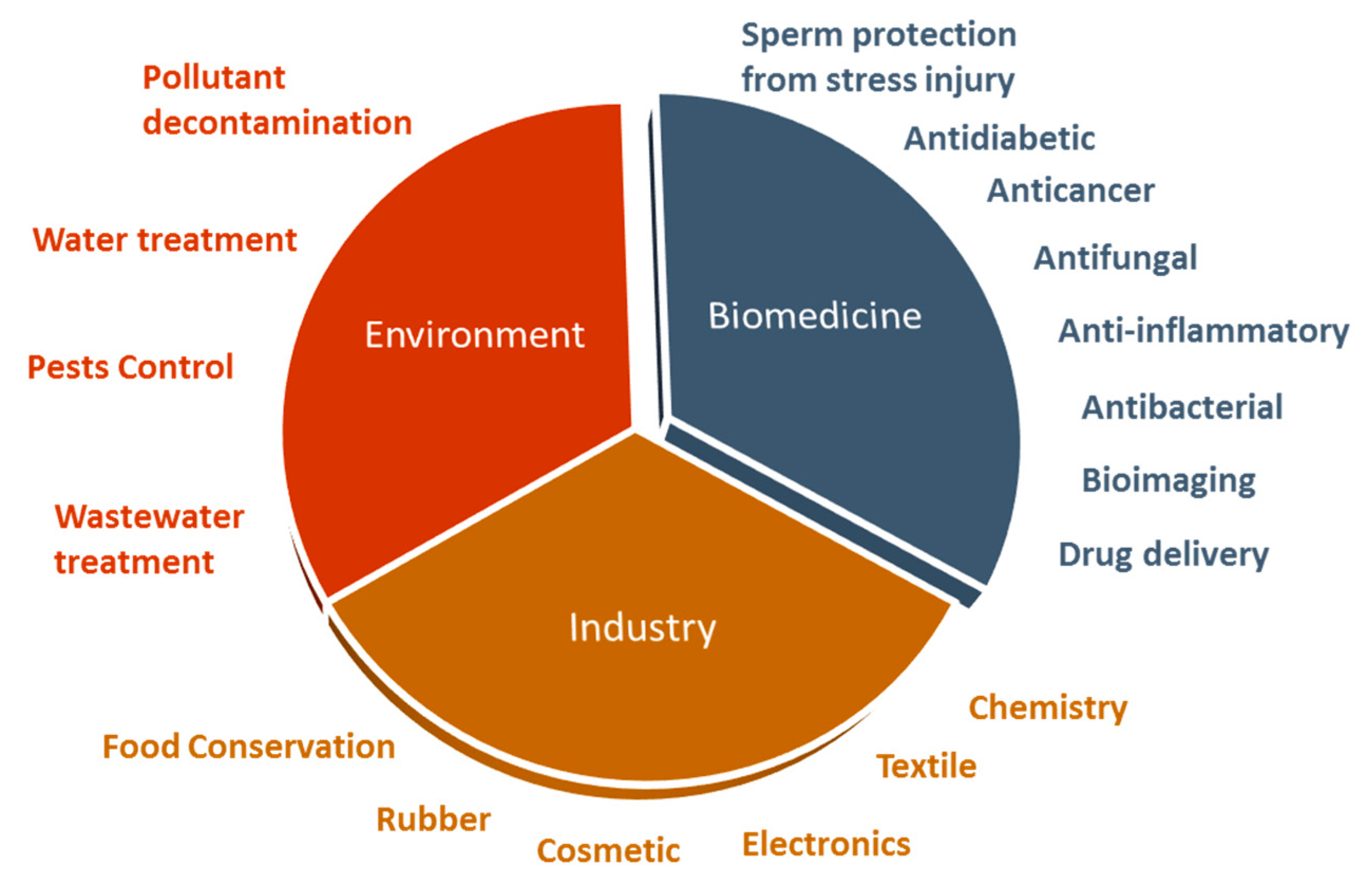
2. ZnO Nanoparticles, A Variety of Biomedical Applications
3. ZnO Nanoparticles: Route of Exposure and Accumulation in Organism
4. ZnO Nanoparticles and their Effects on Male Reproductive System—Analysis of In Vitro and In Vivo Studies
4.1. In Vitro Studies
4.2. In Vivo Studies
4.2.1. In Vivo Studies in Mammalian Animal Models
4.2.2. In Vivo Studies in Non-Mammalian Animal Models
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Buzea, C.; Pacheco, I.I.; Robbie, K. Nanomaterials and nanoparticles: Sources and toxicity. Biointerphases 2007, 2, 17–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United Kingdom Royal Society. Nanoscience and nanotechnologies: Opportunities and uncertainties. Neuroradiology 2004, 46, 618. [Google Scholar] [CrossRef]
- Jeevanandam, J.; Barhoum, A.; Chan, Y.S.; Dufresne, A.; Danquah, M.K. Review on nanoparticles and nanostructured materials: History, sources, toxicity and regulations. Beilstein J. Nanotechnol. 2018, 9, 1050–1074. [Google Scholar] [CrossRef] [Green Version]
- Hulla, J.E.; Sahu, S.C.; Hayes, A.W. Nanotechnology: History and future. Hum. Exp. Toxicol. 2015, 34, 1318–1321. [Google Scholar] [CrossRef] [PubMed]
- Król, A.; Pomastowski, P.; Rafińska, K.; Railean-Plugaru, V.; Buszewski, B. Zinc oxide nanoparticles: Synthesis, antiseptic activity and toxicity mechanism. Adv. Colloid Interface Sci. 2017, 249, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Heiligtag, F.J.; Niederberger, M. The fascinating world of nanoparticle research. Mater. Today 2013, 16, 262–271. [Google Scholar] [CrossRef]
- Wang, N.; Tong, T.; Xie, M.; Gaillard, J.-F. Lifetime and dissolution kinetics of zinc oxide nanoparticles in aqueous media. Nanotechnology 2016, 27, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Saptarshi, S.R.; Duschl, A.; Lopata, A.L. Biological reactivity of zinc oxide nanoparticles with mammalian test systems: An overview. Nanomedicine 2015, 10, 2075–2092. [Google Scholar] [CrossRef] [PubMed]
- Senapati, V.A.; Kumar, A. ZnO nanoparticles dissolution, penetration and toxicity in human epidermal cells. Influence of pH. Environ. Chem. Lett. 2018, 16, 1129–1135. [Google Scholar] [CrossRef]
- Cho, W.S.; Duffin, R.; Howie, S.E.; Scotton, C.J.; Wallace, W.A.; MacNee, W.; Bradley, M.; Megson, I.L.; Donaldson, K. Progressive severe lung injury by zinc oxide nanoparticles; the role of Zn2+ dissolution inside lysosomes. Part. Fibre Toxicol. 2011, 8, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madhumitha, G.; Elango, G.; Roopan, S.M. Biotechnological aspects of ZnO nanoparticles: Overview on synthesis and its applications. Appl. Microbiol. Biotechnol. 2016, 100, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Pi, J.; Cai, J. The Advancing of Zinc Oxide Nanoparticles for Biomedical Applications. Bioinorg. Chem. Appl. 2018, 2018, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Larson, J.K.; Carvan, M.J.; Hutz, R.J. Engineered Nanomaterials: An Emerging Class of Novel Endocrine Disruptors1. Biol. Reprod. 2014, 91, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Agarwal, A. Spermatogenesis: An Overview. In Sperm Chromatin; Springer: Berlin/Heidelberg, Germany, 2011; pp. 127–160. [Google Scholar]
- Guraya, S.S. Biology of Spermatogenesis and Spermatozoa in Mammals; Springer: Berlin/Heidelberg, Germany, 1987. [Google Scholar]
- Hess, R.A.; de Franca, L.R. Spermatogenesis and Cycle of the Seminiferous Epithelium. Adv. Exp. Med. Biol. 2008, 636, 1–15. [Google Scholar] [CrossRef]
- Pereira, C.D.; Serrano, J.B.; Martins, F.; da Cruz e Silva, O.A.B.; Rebelo, S. Nuclear envelope dynamics during mammalian spermatogenesis: New insights on male fertility. Biol. Rev. 2019, 94, 1195–1219. [Google Scholar] [CrossRef]
- Wang, C.; Hu, X.; Gao, Y.; Ji, Y. ZnO Nanoparticles Treatment Induces Apoptosis by Increasing Intracellular ROS Levels in LTEP-a-2 Cells. BioMed Res. Int. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Umrani, R.D.; Paknikar, K.M. Zinc oxide nanoparticles show antidiabetic activity in streptozotocin-induced Type 1 and 2 diabetic rats. Nanomedicine 2014, 9, 89–104. [Google Scholar] [CrossRef]
- Nagajyothi, P.; Sreekanth, T.; Tettey, C.O.; Jun, Y.I.; Mook, S.H. Characterization, antibacterial, antioxidant, and cytotoxic activities of ZnO nanoparticles using Coptidis Rhizoma. Bioorg. Med. Chem. Lett. 2014, 24, 4298–4303. [Google Scholar] [CrossRef]
- Rad, S.S.; Sani, A.M.; Mohseni, S. Biosynthesis, characterization and antimicrobial activities of zinc oxide nanoparticles from leaf extract of Mentha pulegium (L.). Microb. Pathog. 2019, 131, 239–245. [Google Scholar] [CrossRef]
- De Souza, R.C.; Haberbeck, L.U.; Riella, H.G.; Ribeiro, D.H.B.; Carciofi, B.A.M. Antibacterial activity of zinc oxide nanoparticles synthesized by solochemical process. Braz. J. Chem. Eng. 2019, 36, 885–893. [Google Scholar] [CrossRef] [Green Version]
- Dutta, R.; Nenavathu, B.P.; Gangishetty, M.K.; Reddy, A. Studies on antibacterial activity of ZnO nanoparticles by ROS induced lipid peroxidation. Colloids Surf. B Biointerfaces 2012, 94, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-Y.; Xiong, H.-M. Photoluminescent ZnO nanoparticles and their biological applications. Materials 2015, 8, 3101–3127. [Google Scholar] [CrossRef]
- Lipovsky, A.; Nitzan, Y.; Gedanken, A.; Lubart, R. Antifungal activity of ZnO nanoparticles-the role of ROS mediated cell injury. Nanotechnology 2011, 22, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, A.H.; Possebon, L.; Costa, S.D.S. Adverse effects of Metal-based Nanoparticles on Male Reproductive Cells. In Top 10 Contributions on Environmental Health, 2nd ed.; AvidScience: Berlin, Germany, 2018; pp. 1–19. [Google Scholar]
- Pasquet, J.; Chevalier, Y.; Couval, E.; Bouvier, D.; Noizet, G.; Morlière, C.; Bolzinger, M.-A. Antimicrobial activity of zinc oxide particles on five micro-organisms of the Challenge Tests related to their physicochemical properties. Int. J. Pharm. 2014, 460, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Applerot, G.; Lipovsky, A.; Dror, R.; Perkas, N.; Nitzan, Y.; Lubart, R.; Gedanken, A. Enhanced antibacterial actiwity of nanocrystalline ZnO due to increased ROS-mediated cell injury. Adv. Funct. Mater. 2009, 19, 842–852. [Google Scholar] [CrossRef]
- Jalal, R.; Goharshadi, E.K.; Abareshi, M.; Moosavi, M.; Yousefi, A.; Nancarrow, P. ZnO nanofluids: Green synthesis, characterization, and antibacterial activity. Mater. Chem. Phys. 2010, 121, 198–201. [Google Scholar] [CrossRef]
- Jones, N.; Ray, B.; Ranjit, K.T.; Manna, A.C. Antibacterial activity of ZnO nanoparticle suspensions on a broad spectrum of microorganisms. FEMS Microbiol Lett. 2008, 279, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Jiang, Y.; Ding, Y.; Povey, M.; York, D. Investigation into the antibacterial behaviour of suspensions of ZnO nanoparticles (ZnO nanofluids). J. Nanopart. Res. 2007, 9, 479–489. [Google Scholar] [CrossRef]
- Stoimenov, P.K.; Klinger, R.L.; Marchin, G.L.; Klabunde, K.J. Metal oxide nanoparticles as bactericidal agents. Langmuir 2002, 18, 6679–6686. [Google Scholar] [CrossRef]
- Heinlaan, M.; Ivask, A.; Blinova, I.; Dubourguier, H.C.; Kahru, A. Toxicity of nanosized and bulk ZnO, CuO and TiO2 to bacteria Vibrio fischeri and crustaceans Daphnia magna and Thamnocephalus platyurus. Chemosphere 2008, 71, 1308–1316. [Google Scholar] [CrossRef]
- Huang, Z.; Zheng, X.; Yan, D.; Yin, G.; Liao, X.; Kang, Y.; Yao, Y.; Huang, D.; Hao, B. Toxicological effect of ZnO nanoparticles based on bacteria. Langmuir 2008, 24, 4140–4144. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhu, L.; Lin, D.V. Toxicity of ZnO nanoparticles to escherichia Coli: Mechanism and the influence of medium components. Environ. Sci. Technol. 2011, 45, 1977–1983. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.K.; Mishra, H.; Ekielski, A.; Talegaonkar, S.; Vaidya, B. Zinc oxide nanoparticles: A promising nanomaterial for biomedical applications. Drug Discov. Today 2017, 22, 1825–1834. [Google Scholar] [CrossRef]
- Sawai, J.; Yoshikawa, T. Quantitative evaluation of antifungal activity of metallic oxide powders (MgO, CaO and ZnO) by an indirect conductimetric assay. J. Appl. Microbiol. 2004, 96, 803–809. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Liu, Y.; Mustapha, A.; Lin, M. Antifungal activity of zinc oxide nanoparticles against Botrytis cinerea and Penicillium expansum. Microbiol. Res. 2011, 166, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lee, J.S.; Kim, D.; Zhu, L. Exploration of Zinc Oxide Nanoparticles as a Multitarget and Multifunctional Anticancer Nanomedicine. ACS Appl. Mater. Interfaces 2017, 9, 39971–39984. [Google Scholar] [CrossRef]
- Ostrovsky, S.; Kazimirsky, G.; Gedanken, A.; Brodie, C. Selective cytotoxic effect of ZnO nanoparticles on glioma cells. Nano Res. 2009, 2, 882–890. [Google Scholar] [CrossRef] [Green Version]
- De Berardis, B.; Civitelli, G.; Condello, M.; Lista, P.; Pozzi, R.; Arancia, G.; Meschini, S. Exposure to ZnO nanoparticles induces oxidative stress and cytotoxicity in human colon carcinoma cells. Toxicol. Appl. Pharmacol. 2010, 246, 116–127. [Google Scholar] [CrossRef]
- Setyawati, M.I.; Tay, C.Y.; Leong, D.T. Effect of zinc oxide nanomaterials-induced oxidative stress on the p53 pathway. Biomaterials 2013, 34, 10133–10142. [Google Scholar] [CrossRef]
- Hanley, C.; Layne, J.; Punnoose, A.; Reddy, K.M.; Coombs, I.; Coombs, A.; Feris, K.; Wingett, D. Preferential killing of cancer cells and activated human T cells using ZnO nanoparticles. Nanotechnology 2008, 19, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Tripathy, N.; Ahmad, R.; Ko, H.A.; Khang, G.; Hahn, Y.B. Enhanced anticancer potency using an acid-responsive ZnO-incorporated liposomal drug-delivery system. Nanoscale 2015, 7, 4088–4096. [Google Scholar] [CrossRef]
- Rasmussen, J.W.; Martinez, E.; Louka, P.; Wingett, D.G. Zinc oxide nanoparticles for selective destruction of tumor cells and potential for drug delivery applications. Expert. Opin. Drug Deliv. 2010, 7, 1063–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, H.; Kumar, K.; Choudhary, C.; Mishra, P.K.; Vaidya, B. Development and characterization of metal oxide nanoparticles for the delivery of anticancer drug. Artif. Cells Nanomed. Biotechnol. 2016, 44, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, S.B.; Sarrafzadeh, M.H.; Fakhroueian, Z.; Shahriari, S.; Khorramizadeh, M.R. Functionalization of ZnO nanoparticles by 3-mercaptopropionic acid for aqueous curcumin delivery: Synthesis, characterization, and anticancer assessment. Mater. Sci. Eng. C 2017, 79, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, H.; Nakara, A.; Shanmugam, V.K. Anti-inflammatory mechanism of various metal and metal oxide nanoparticles synthesized using plant extracts: A review. Biomed. Pharmacother. 2019, 109, 2561–2572. [Google Scholar] [CrossRef] [PubMed]
- Nagajyothi, P.C.; Cha, S.J.; Yang, I.J.; Sreekanth, T.V.M.; Kim, K.J.; Shin, H.M. Antioxidant and anti-inflammatory activities of zinc oxide nanoparticles synthesized using Polygala tenuifolia root extract. J. Photochem. Photobiol. B Biol. 2015, 146, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, M.A.; Norredin, A. The Potential Contribution of Nanoparticles in the Treatment of Inflammatory Diseases. In Inflammation-From Molecular Basis to Clinical Outcomes Cause; IntechOpen: London, UK, 2019; pp. 1–15. [Google Scholar]
- Ilves, M.; Palomäki, J.; Vippola, M.; Lehto, M.; Savolainen, K.; Savinko, T.; Alenius, H. Topically applied ZnO nanoparticles suppress allergen induced skin inflammation but induce vigorous IgE production in the atopic dermatitis mouse model. Part. Fibre Toxicol. 2014, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, C.; Hipler, U.C.; Boldt, S.; Strehle, J.; Wollina, U. Skin-protective effects of a zinc oxide-functionalized textile and its relevance for atopic dermatitis. Clin. Cosmet. Investig. Dermatol. 2013, 6, 115–121. [Google Scholar] [CrossRef] [Green Version]
- El-Gharbawy, R.M.; Emara, A.M.; Abu-Risha, S.E.S. Zinc oxide nanoparticles and a standard antidiabetic drug restore the function and structure of beta cells in Type-2 diabetes. Biomed. Pharmacother. 2016, 84, 810–820. [Google Scholar] [CrossRef]
- Wahba, N.S.; Shaban, S.F.; Kattaia, A.A.A.; Kandeel, S.A. Efficacy of zinc oxide nanoparticles in attenuating pancreatic damage in a rat model of streptozotocin-induced diabetes. Ultrastruct. Pathol. 2016, 40, 358–373. [Google Scholar] [CrossRef]
- Alkaladi, A.; Abdelazim, A.M.; Afifi, M. Antidiabetic activity of zinc oxide and silver nanoparticles on streptozotocin-induced diabetic rats. Int. J. Mol. Sci. 2014, 15, 2015–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afifi, M.; Almaghrabi, O.A.; Kadasa, N.M. Ameliorative Effect of Zinc Oxide Nanoparticles on Antioxidants and Sperm Characteristics in Streptozotocin-Induced Diabetic Rat Testes. BioMed Res. Int. 2015, 2015, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, D.A.; Abdelrahman, S.A. The possible protective role of zinc oxide nanoparticles (ZnO NPs) on testicular and epididymal structure and sperm parameters in nicotine-treated adult rats (a histological and biochemical study). Cell Tissue Res. 2019, 375, 543–558. [Google Scholar] [CrossRef] [PubMed]
- Torabi, F.; Shafaroudi, M.M.; Rezaei, N. Combined protective effect of zinc oxide nanoparticles and melatonin on cyclophosphamide-induced toxicity in testicular histology and sperm parameters in adult Wistar rats. Int. J. Reprod. Biomed. 2017, 15, 403–412. [Google Scholar] [CrossRef] [Green Version]
- El-Maddawy, Z.K.; Abd El Naby, W.S.H. Protective effects of zinc oxide nanoparticles against doxorubicin induced testicular toxicity and DNA damage in male rats. Toxicol. Res. 2019, 8, 654–662. [Google Scholar] [CrossRef]
- Isaac, A.V.; Kumari, S.; Nair, R.; Urs, D.R.; Salian, S.R.; Kalthur, G.; Adiga, S.K.; Manikkath, J.; Mutalik, S.; Sachdev, D.; et al. Supplementing zinc oxide nanoparticles to cryopreservation medium minimizes the freeze-thaw-induced damage to spermatozoa. Biochem. Biophys. Res. Commun. 2017, 494, 656–662. [Google Scholar] [CrossRef]
- Walker, N.J.; Bucher, J.R. A 21st century paradigm for evaluating the health hazards of nanoscale materials? Toxicol. Sci. 2009, 110, 251–254. [Google Scholar] [CrossRef] [Green Version]
- Lan, Z.; Yang, W.X. Nanoparticles and spermatogenesis: How do nanoparticles affect spermatogenesis and penetrate the blood-testis barrier. Nanomedicine 2012, 7, 579–596. [Google Scholar] [CrossRef]
- Chen, A.; Feng, X.; Sun, T.; Zhang, Y.; An, S.; Shao, L. Evaluation of the effect of time on the distribution of zinc oxide nanoparticles in tissues of rats and mice: A systematic review. IET Nanobiotechnol. 2016, 10, 97–106. [Google Scholar] [CrossRef]
- Abbasalipourkabir, R.; Moradi, H.; Zarei, S.; Asadi, S.; Salehzadeh, A.; Ghafourikhosroshahi, A.; Mortazavi, M.; Ziamajidi, N. Toxicity of zinc oxide nanoparticles on adult male Wistar rats. Food Chem. Toxicol. 2015, 84, 154–160. [Google Scholar] [CrossRef]
- Hao, Y.; Liu, J.; Feng, Y.; Yu, S.; Zhang, W.; Li, L.; Min, L.; Zhang, H.; Shen, W.; Zhao, Y. Molecular evidence of offspring liver dysfunction after maternal exposure to zinc oxide nanoparticles. Toxicol. Appl. Pharmacol. 2017, 329, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Kuang, H.; Yang, P.; Yang, L.; Aguilar, Z.P.; Xu, H. Size dependent effect of ZnO nanoparticles on endoplasmic reticulum stress signaling pathway in murine liver. J. Hazard. Mater. 2016, 317, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.Y.; Chiu, J.I.; Cheng, Y.F.; Lee, H.Y.; Wang, J.Y.; Hsu, H.Y.; Chiu, W.H. The role of hypoxia-inducible factor-1α in zinc oxide nanoparticle-induced nephrotoxicity in vitro and in vivo. Part. Fibre Toxicol. 2015, 13, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.Y.; Wong, E.W.P.; Yan, H.H.N.; Mruk, D.D. Regulation of Spermatogenesis in the Microenvironment of the Seminiferous Epithelium: New Insights and Advances. Mol. Cell. Endocrinol. 2010, 315, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kretser, D.M.; Loveland, K.L.; Meinhardt, A.; Simorangkir, D.; Wreford, N. Spermatogenesis. Hum. Reprod. 1998, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Xu, C.; Ji, G.; Liu, H.; Mo, Y.; Tollerud, D.J.; Gu, A.; Zhang, Q. Sublethal effects of zinc oxide nanoparticles on male reproductive cells. Toxicol. In Vitro 2016, 35, 131–138. [Google Scholar] [CrossRef]
- Han, Z.; Yan, Q.; Ge, W.; Liu, Z.-G.; Gurunathan, S.; De Felici, M.; Shen, W.; Zhang, X.-F. Cytotoxic effects of ZnO nanoparticles on mouse testicular cells. Int. J. Nanomed. 2016, 11, 5187–5203. [Google Scholar] [CrossRef] [Green Version]
- Li, M.W.M.; Wong, V.; Szeto, L.; Uffelman, K.; Fantus, I.G.; Lewis, G.F. Tumor necrosis factor α reversibly disrupts the blood-testis barrier and impairs Sertoli-germ cell adhesion in the seminiferous epithelium of adult rat testes. J. Endocrinol. 2006, 190, 313–329. [Google Scholar] [CrossRef] [Green Version]
- Hellani, A.; Ji, J.; Mauduit, C.; Deschildre, C.; Tabone, E.; Benahmed, M. Developmental and hormonal regulation of the expression of oligodendrocyte-specific protein/claudin 11 in mouse testis. Endocrinology 2000, 141, 3012–3019. [Google Scholar] [CrossRef]
- Barkhordari, A.; Hekmatimoghaddam, S.; Jebali, A.; Khalili, M.A.; Talebi, A.; Noorani, M. Effect of zinc oxide nanoparticles on viability of human spermatozoa. Iran. J. Reprod. Med. 2013, 11, 767–771. [Google Scholar]
- Shen, J.; Yang, D.; Zhou, X.; Wang, Y.; Tang, S.; Yin, H.; Wang, J.; Chen, R.; Chen, J. Role of Autophagy in Zinc Oxide Nanoparticles-Induced Apoptosis of Mouse LEYDIG Cells. Int. J. Mol. Sci. 2019, 20, 4042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talebi, A.R.; Khorsandi, L.; Moridian, M. The effect of zinc oxide nanoparticles on mouse spermatogenesis. J. Assist. Reprod. Genet. 2013, 30, 1203–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mozaffari, Z.; Parivar, K.; Roodbari, N.H.; Irani, S. Histopathological evaluation of the toxic effects of zinc oxide (ZnO) nanoparticles on testicular tissue of NMRI adult mice. Adv. Stud. Biol. 2015, 7, 275–291. [Google Scholar] [CrossRef] [Green Version]
- Hussein, M.M.A.; Ali, H.A.; Saadeldin, I.M.; Ahmed, M.M. Querectin Alleviates Zinc Oxide Nanoreprotoxicity in Male Albino Rats. J. Biochem. Mol. Toxicol. 2016, 30, 489–496. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, B.; Huo, L.; Polli, R.; Qiu, L.; Collier, N.; Zhang, B.; Pan, X. ZnO nanoparticles enhanced germ cell apoptosis in Caenorhabditis elegans. In comparison with ZnCl2. Toxicol. Sci. 2017, 156, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Salman, R.A. The Influence of ZnO NPs on Reproductive System Tissues of Albino Male Mice. Histopathological Study. Int. J. Sci. Res. 2017, 6, 2021–2025. [Google Scholar] [CrossRef]
- Srivastav, A.K.; Kumar, A.; Prakash, J.; Singh, D.; Jagdale, P.; Shankar, J.; Kumar, M. Genotoxicity evaluation of zinc oxide nanoparticles in Swiss mice after oral administration using chromosomal aberration, micronuclei, semen analysis, and RAPD profile. Toxicol. Ind. Health 2017, 33, 821–834. [Google Scholar] [CrossRef]
- Aporvari, H.A.; Mamoei, M.; Vakili, S.T.; Zareei, M.; Davachi, N.D. The effect of oral administration of zinc oxide nanoparticles on quantitative and qualitative properties of arabic ram sperm and some antioxidant parameters of seminal plasma in the non-breeding season. Arch. Razi Inst. 2018, 73, 121–129. [Google Scholar] [CrossRef]
- Khoobbakht, Z.; Mohammadi, M.; Mehr, M.R.A.; Mohammadghasemi, F.; Sohani, M.M. Comparative effects of zinc oxide, zinc oxide nanoparticle and zinc-methionine on hatchability and reproductive variables in male Japanese quail. Anim. Reprod. Sci. 2018, 192, 84–90. [Google Scholar] [CrossRef]
- Radhi, M.J.; Al-bairuty, G. Effect of Zinc oxide nanoparticles (ZnO-NPs) on weights of some reproductive organs and sperm abnormalities in the tail of epididymis of albino mice. J. Pharm. Sci. Res. 2019, 11, 243–246. [Google Scholar]
- Mesallam, D.I.; Deraz, R.H.; Aal, S.M.A.; Ahmed, S.M. Toxicity of Subacute Oral Zinc Oxide Nanoparticles on Testes and Prostate of Adult Albino Rats and Role of Recovery. J. Histol. Histopathol. 2019, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Chen, B.; Hong, W.; Chen, L.; Yao, L.; Zhao, Y.; Aguilar, Z.P.; Xu, H. ZnO Nanoparticles Induced Male Reproductive Toxicity Based on the Effects on the Endoplasmic Reticulum Stress Signaling Pathway. Int. J. Nanomed. 2019, 14, 9563–9576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewing, L.L.; Zirkin, B. Leydig cell structure and steroidogenic function. Recent Prog. Horm. Res. 1983, 39, 599–635. [Google Scholar] [CrossRef] [PubMed]
- Payne, A.H.; Hardy, M.P. The Leydig Cell in Health and Disease; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Wang, H.; Wang, Q.; Zhao, X.F.; Liu, P.; Meng, X.H.; Yu, T.; Ji, Y.L.; Zhang, H.; Zhang, C.; Zhang, Y.; et al. Cypermethrin exposure during puberty disrupts testosterone synthesis via downregulating StAR in mouse testes. Arch. Toxicol. 2010, 84, 53–61. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| ZnO NPs’ Characteristics | Objective | Cell Type | ZnO NPs Concentration (μg/mL) | Parameters | Results | Reference |
|---|---|---|---|---|---|---|
| Size: 50 nm Shape: amorphous | Evaluate the cytotoxicity of ZnO NPs on viability of spermatozoa | Spermatozoa (Human) | 10, 100, 500, 1000 | —Viability | —The toxicity depends on concentration and time of exposure; —Higher concentrations and higher exposure periods induce higher toxicity | [74] |
| Size: 70 nm Shape: spherical Nature: crystalline Dispersion: polydisperse and agglomerate in quasi-spherical and hexagonal structures Surface roughness: high (22.9 nm) | Investigate the toxicity of ZnO NPs in testicular cells | Leydig cell Sertoli cell (Mouse) | 0, 5, 10, 15, 20 | —Cellular uptake of ZnO NPs —Viability —MMP and ROS levels —Apoptosis and DNA Damage | —ZnO NP aggregates in the cytoplasm and in nucleus; —The toxicity depends on the concentration (≥10 μg/ml) and the exposure time (≥6 h), and not on Zn2+ release; —ROS production increase, leading to loss of MMP originating apoptosis of Leydig and Sertoli cells and DNA damage | [71] |
| Size: 177 nm Shape: spheroid or ellipsoid Zeta Potential: −27.4 ± 1.0 mV Purity: >97% | Explore the effects of sublethal doses of ZnO NPs and their underlying mechanisms on male germ cells | Sertoli cell Spermatocyte (Mouse) | 0, 0.04, 0.08, 0.4, 0.8, 4, 8, and 16 | —Viability —ROS, GSH, and MDA levels; —Permeability, MMP, and cytochrome C —BTB junction proteins levels; —Erk1/2 and TNF-α levels; —DNA damage and cell cycle; | —The sublethal dose of ZnO NP is 8 µg/mL; —GSH levels decrease and MDA levels increase; —Sertoli cell membrane disruption and cellular invasion; —ROS levels production increase compromising BTB, by down-regulating the expression of BTB junction proteins, causing DNA damage and cell cycle arrest at S-phase at spermatocytes | [70] |
| Size: 30 nm; HS: 66.36 ± 0.39 nm; Zeta Potential: 38.25 ± 1.06 mV | Investigate whether oxidative stress was involved in ZnO NPs-induced apoptosis and autophagy of mouse Leydig cells, and to determine the role of autophagy in ZnO NPs-induced apoptosis. | Leydig cell (Mouse) | 0, 2, 3, 4, and 8 | —Cell Viability —Bax, Bcl-2, cleaved Caspase-3, cleaved Caspase-8, LC3-I, LC3-II, Atg5, and Beclin 1 protein level from testis —Cell biochemistry: SOD, GPx, MDA, GSH | —Cell viability inhibition and apoptosis induction by oxidative stress; —Autophagy plays a cytoprotective role in ZnO NPs-induced | [75] |
| ZnO NPs Characteristics | Objective | Animal Model/Tissue or Organ of Study | Administration via of Exposure | Evaluated Parameters | ZnO NPs Concentration | Results | Reference |
|---|---|---|---|---|---|---|---|
| Evaluate the effects of ZnO NPs on spermatogenesis | NMRI mice Semen Testis | Oral | —Epididymal sperm; —Testicular histology; —SE morphometry | 0, 5, 50, and 300 mg/kg | —Cytotoxicity in testicular germ cells in a dose-dependent manner (≥50 mg/kg): testis histological alterations; —Increased sperm abnormalities; —Reduction of sperm and Leydig cells number | [76] | |
| Size: 10–30 nm; SA: 20/30 m2/g; Colour: milky white; Crystal phase: single; CM: nearly spherical; Density: 5.606 g/cm3 Purity: ≥99% | Investigate the effects of ZnO NPs on adult male Wistar rats | Wistar rats Epididymis Sperm Blood | Ip | —Epididymal sperm; —Serum biochemistry: SOD, GPx, MDA, TOS, TAC | 0, 50, 100, 150, and 200 mg/kg | —Viability and sperm number decrease (≥50 mg/kg); —Poor sperm quality (≥100 mg/kg); —Antioxidant capacity decrease (200 mg/kg) | [64] |
| Size: 20 nm SA: > 90 m2/g Colour: white CM: Nearly spherical Purity: ≥99% | Investigate the effects of ZnO NPs at different doses on testis of adult mice | NMRI male mice Testis | Ip | —Testicular histology | 0, 250, 500, and 700 mg/kg daily | —Testis histological alterations; —Reduction and degeneration of spermatogonia, primary spermatocyte, spermatid, sperm, and Leydig cells; —Spermatogenesis arrest | [77] |
| Size: 70 nm Shape: spherical Nature: crystalline Dispersion: polydisperse Surface roughness: high (22.9 nm) | Investigate the toxicity of ZnO NPs in testicular cells | Cd1 mice (21 day old) Epididymis Sperm Testis | Intravenous | —Sperm morphology; —SE morphometry | 0, 1, and 5 mg/kg | Spermatogenesis damage by alteration of SE inducing sperm abnormalities (≥5 mg/kg, ≥49 days) | [71] |
| Size: <50 nm; SA: >10.8 m2/g; Purity: >97%; | Evaluate the toxicological effect of ZnO NPs on male fertility and the amelioration with querectin in Wistar Han rats. | Wistar Han rats Epididymis Sperm Testis | Intragastric intubation | —Testis biochemistry: MDA, CAT, SOD, GPx, GSH; —Epididymal sperm; —Testicular histology; —Serum testosterone level; 3β-HSD; 17β-HSD; Nr5A1 mRNA levels | 0, 100, and 400 mg/kg | —Decrease of sperm live cell and Leydig cell number; —Serum testosterone level decrease and increase of abnormal sperm —Atrophy, and necrosis of ST in a dose-dependent manner; —Antioxidant capacity decrease and oxidative stress increase | [78] |
| Size: 17.9 ± 7–3 nm Distribution range: 1–55 nm HS: 721 ± 109.5 nm Purity: ≈100% Surface area: 15–25 g/m2; | Unravel the effects of ZnO NPs exposure on germ cell apoptosis and apoptosis-related gene expressions | Caenorhabditis elegans Egg | Mix of ZnO NPs or ZnCl2 into nematode growth medium (NGM) agar | —Apoptosis genes expression: ced-13, ced-3, ced-4, ced-9, cep-1, dpl-1, efl-1, efl-2, egl-1, egl-38, lin-35, pax-2, and sir-2.1. | 0, 6.14 X10−1, 61.4, and 614 µM | Apoptosis of germ cells (≥61,4 µM) by upregulation of apoptosis genes (cep-1, cep-13, efl-2, egl-1, lin-35, and sir-2.1 (≥614 µM). Enhanced apoptotic effects were not fully attributed to ionic Zn, ZnO NPs also have the capacity to affect apoptotic machinery | [79] |
| Size: 80 nm | Investigate side effects of various doses of ZnO NPs on the reproductive system of albino mice | Adult albino mice Testis Prostate Seminal vesicles Epididymis | Oral | —Male reproductive system histology | 0, 150, and 350 mg/kg | —Cytotoxicity of testicular tissue in a dose-dependent manner; —Damages in all tissues of reproductive system (testis, seminal vesicles, prostate, and epididymis (350 mg/kg)) | [80] |
| Size: 39.45 +19.88 nm HS: 447.5 nm Shape: hexagonal Aggregation: large and irregular Polydispersity index: 0.13 nm Zeta Potential: −32.1 mV | Evaluate the genotoxic effect of ZnO NPs in Swiss mice | Swiss male mice Semen Liver Bone Marrow | Oral | —Semen; —Genotoxicity in blood samples: CA, MnPCEs, DNA damage; —ROS levels in liver | 300–2000 mg/kg | —Genotoxic effects in a dose-dependent manner by ROS generation (2000 mg/kg): leading to genomic integrity and anomalies in spermatogenesis; —Chromosomal alterations and generation of micronucleus in bone marrow cells of male mice. | [81] |
| Size: 10–30 nm | Investigate the effect of ZnO NPs on some of the antioxidant parameters of semen plasma, quantitative and qualitative properties of Arabic ram sperm | Arabic sheep Semen | Oral | —Semen; —Membrane integrity; —SOD and TAC | 0, 40, and 80 mg/kg | Improves the qualitative and quantitative properties of sperm and some antioxidant parameters of seminal plasma, neutralizing the ROS effects (80 mg/kg). | [82] |
| Size: 10–30 nm Purity: 99.9% | Investigate the effects of different zinc source (nano, organic, and inorganic) supplementations on the reproduction of male Japanese quail. | Japanese quail chick (one-day old) Testis Eggs | Oral | —Index of cloacal gland size; —GSI; STD; GET; —Serum testosterone level; —Fertility; —Hatchability | 0, 25, and 50 mg/kg | Detrimental effects on reproduction, by reducing hatchability and, also, inducing abnormalities in Japanese quail embryos | [83] |
| Size: 50 nm; Shape: cube; Colour: White Purity: 99.99%; | Evaluate the effects of ZnO NPs on the weight of epididymis, testis, seminal vesicle, and prostate and identify abnormalities of epididymis sperm of albino rats | NMRI male mice Testis Seminal Vesicles Prostate Sperm Epididymis | Oral | —Testis, epididymis, seminal vesicle, and prostate histology; —Testis, epididymis, seminal vesicle, and prostate weights | 0, 100, and 200 mg/kg | —Testicular and epididymis weight decrease and hypertrophy of seminal vesicle and prostate; —Increase of epididymal sperm abnormalities | [84] |
| Size: <100 nm; Purity: ≥99.5% Colour: white | Detect the effects of ZnO NPs on the testes and prostate of adult albino rats and recovery | Male albino rats Serum Testis Prostate | Oral | —Serum biochemistry: MDA, GSH, CAT, SOD; —Testicular and prostatic cytokines: TNF-α, IL-4; —DNA fragmentation; —Testis and prostate histology | 0 and 422 mg/kg | —Increase of oxidative stress and decrease of antioxidant capacity in serum; —Inflammatory response and DNA fragmentation increase, in testis and prostate and histological changes; —Limited exposure to ZnO NPs allows recovery of damaged tissue | [85] |
| Size: 30 nm; HS: 66.36 ± 0.39 nm; Zeta Potential: 38.25 ± 1.06 mV | Investigate whether oxidative stress was involved in ZnO NPs-induced apoptosis and autophagy of mouse Leydig cells, and to determine the role of Autophagy in ZnO NPs-induced apoptosis. | Male Kunming mice Serum Epididymis Sperm Testis | Intragastric | —Serum testosterone level; —Testis and epididymis histology; —Bax, cleaved Caspase-3, cleaved Caspase-8, LC3-I, LC3-II, Atg5, and Beclin 1 protein level from testis | 0, 100, 200, and 400 mg/kg | —Disruption and atrophy of the SE by apoptosis and germ cell depletion; —Decrease of epididymal sperm density and serum testosterone level; —Induction of autophagy in testis tissue | [75] |
| Size: 30 nm; Shape: spherical | Evaluate the potential reproductive risks in males exposed by gavage to various doses of ZnO NPs | Male Kunming mice Serum Epididymis Sperm Testis | Ip | —Serum testosterone level; —Zinc accumulation; —Testis histology —Gene expression in Testis: BIP, XBP1s, IRE1α, JNK, C/EBP, CHOP, Bax, Bcl-2, Caspase 12, Caspase 13, StAR, P450sc | 0, 50, 150, and 450 mg/kg | —Accumulation of nanoparticles contributed to seminiferous tubules degeneration and sperm cell number diminution via apoptosis and ER-stress signalling pathway; —Decrease testosterone production through the downregulation of StAR. | [86] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinho, A.R.; Rebelo, S.; Pereira, M.d.L. The Impact of Zinc Oxide Nanoparticles on Male (In)Fertility. Materials 2020, 13, 849. https://doi.org/10.3390/ma13040849
Pinho AR, Rebelo S, Pereira MdL. The Impact of Zinc Oxide Nanoparticles on Male (In)Fertility. Materials. 2020; 13(4):849. https://doi.org/10.3390/ma13040849
Chicago/Turabian StylePinho, Ana Rita, Sandra Rebelo, and Maria de Lourdes Pereira. 2020. "The Impact of Zinc Oxide Nanoparticles on Male (In)Fertility" Materials 13, no. 4: 849. https://doi.org/10.3390/ma13040849
APA StylePinho, A. R., Rebelo, S., & Pereira, M. d. L. (2020). The Impact of Zinc Oxide Nanoparticles on Male (In)Fertility. Materials, 13(4), 849. https://doi.org/10.3390/ma13040849