
Protein-Mediated Precipitation of Calcium Carbonate



Abstract
:1. Introduction
2. Results and Discussion
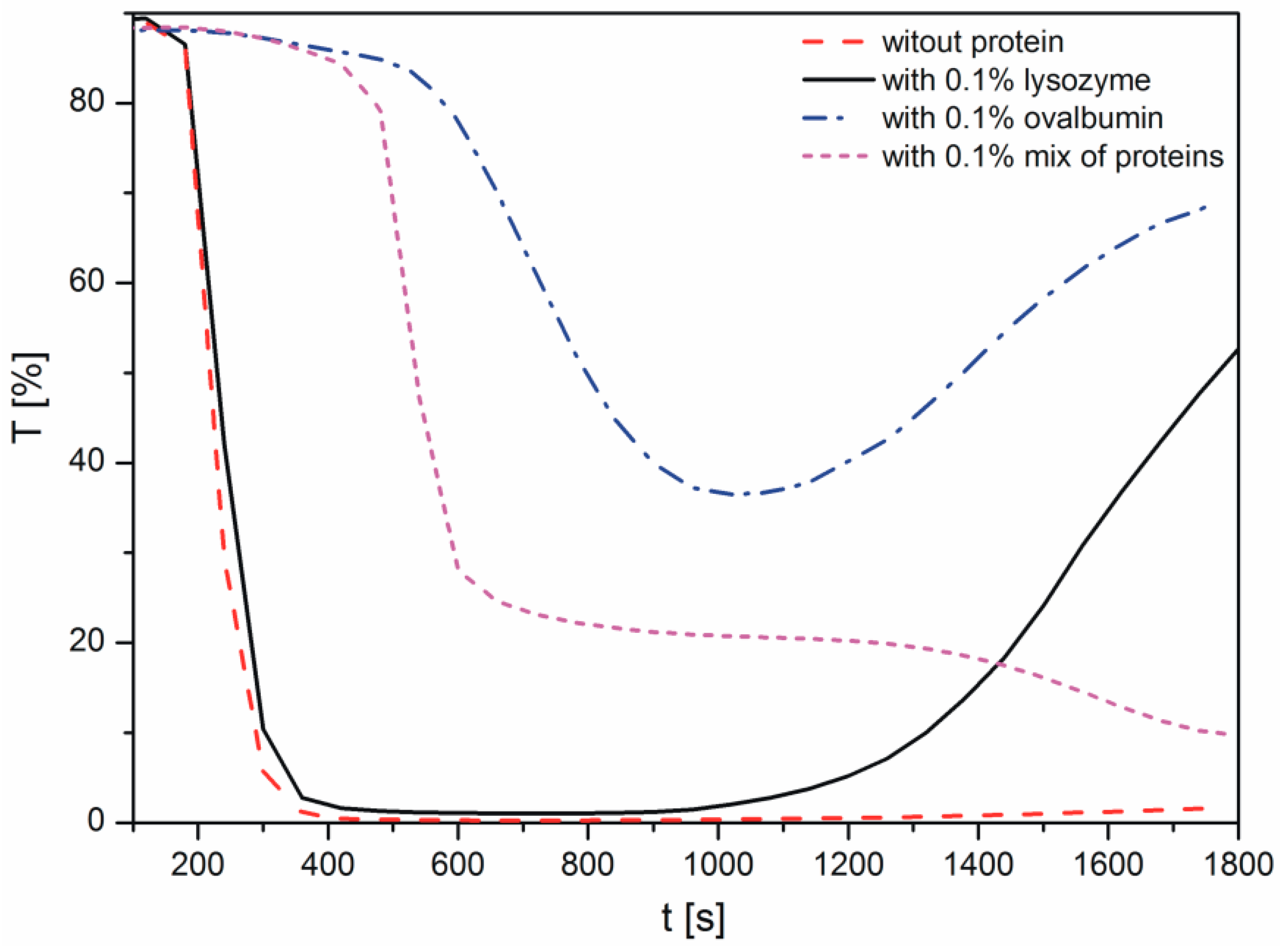
2.1. Kinetics of Calcium Carbonate Crystals Formation
2.2. SEM Analysis of the Calcium Carbonate Crystals
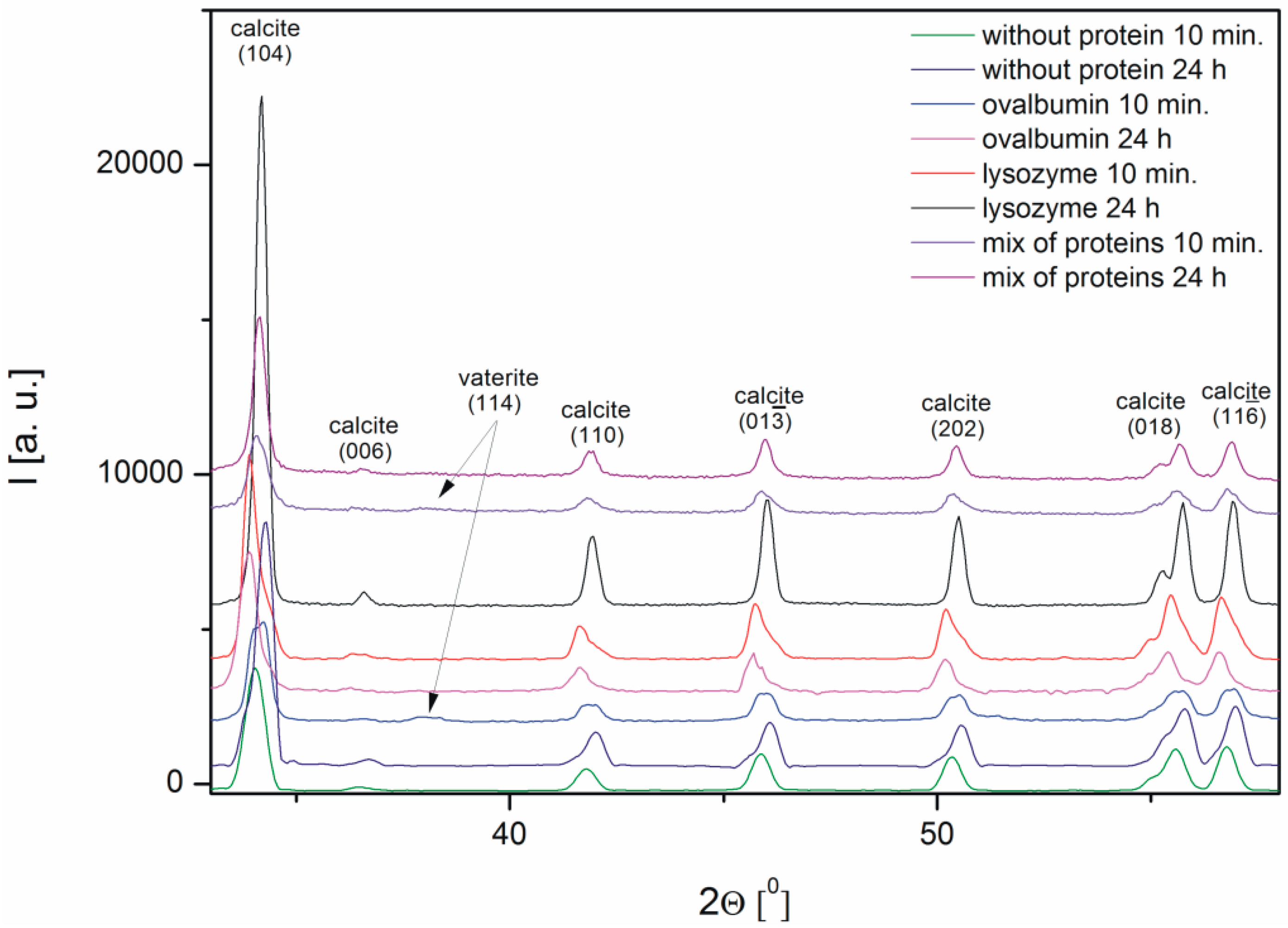
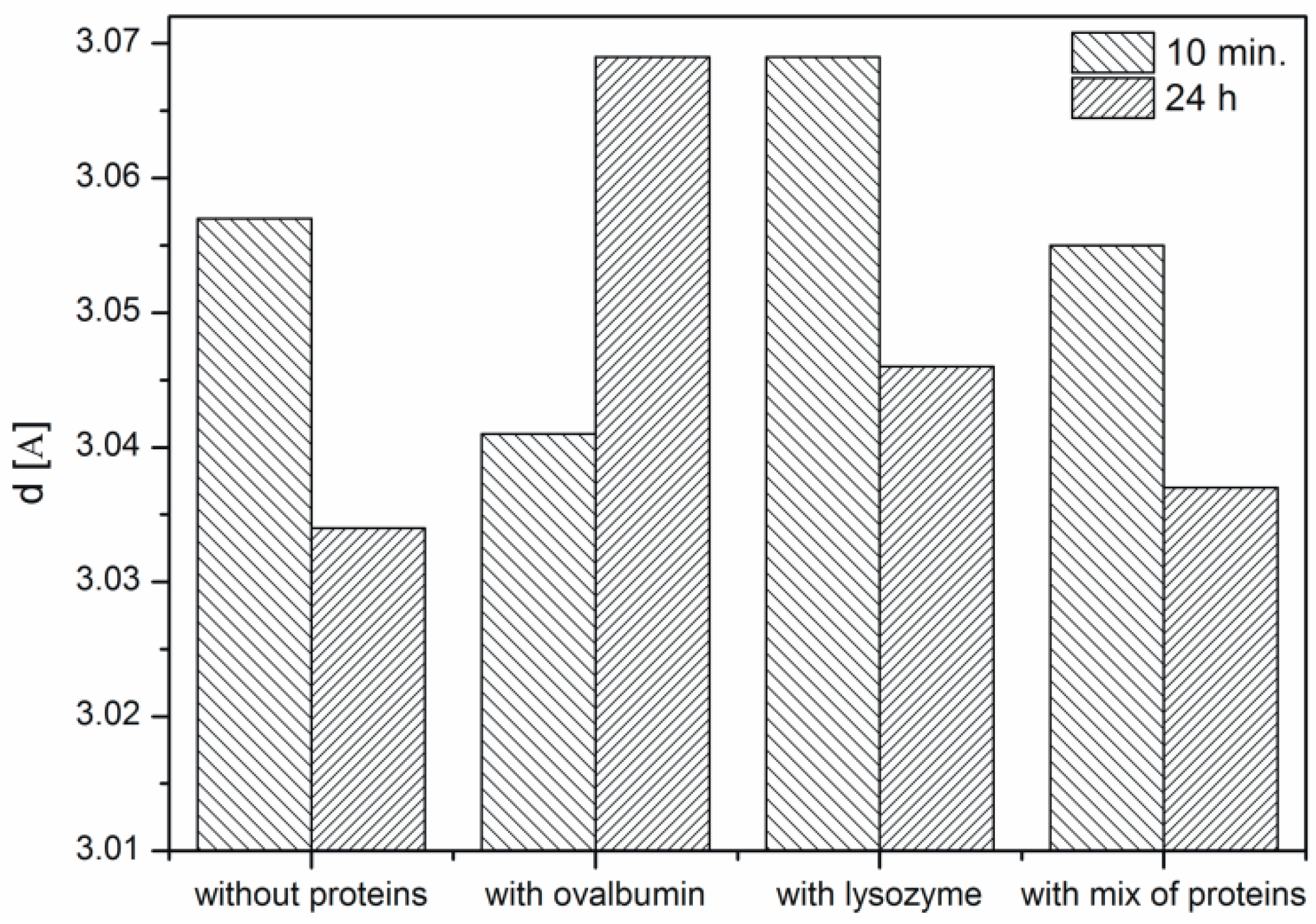
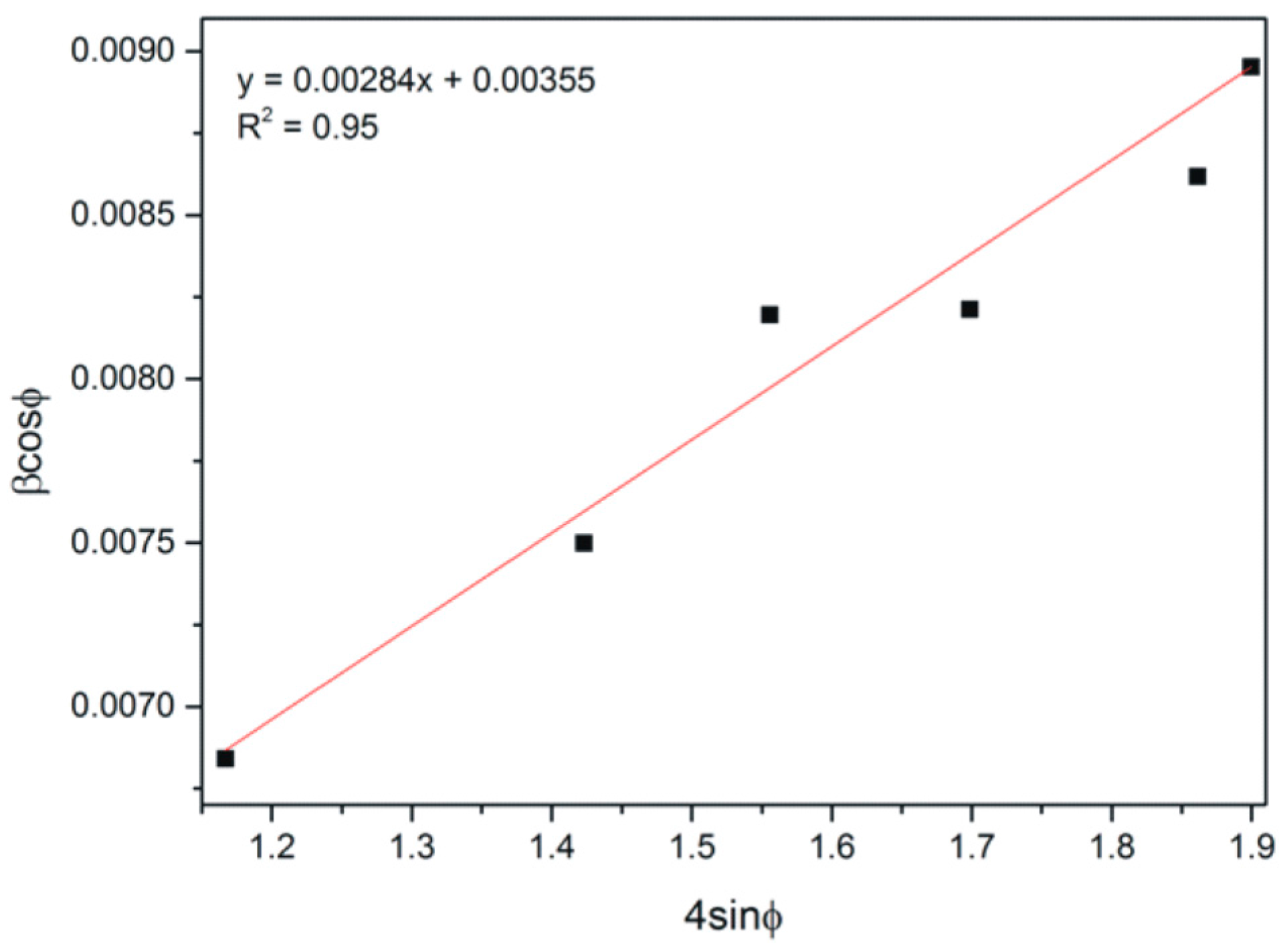
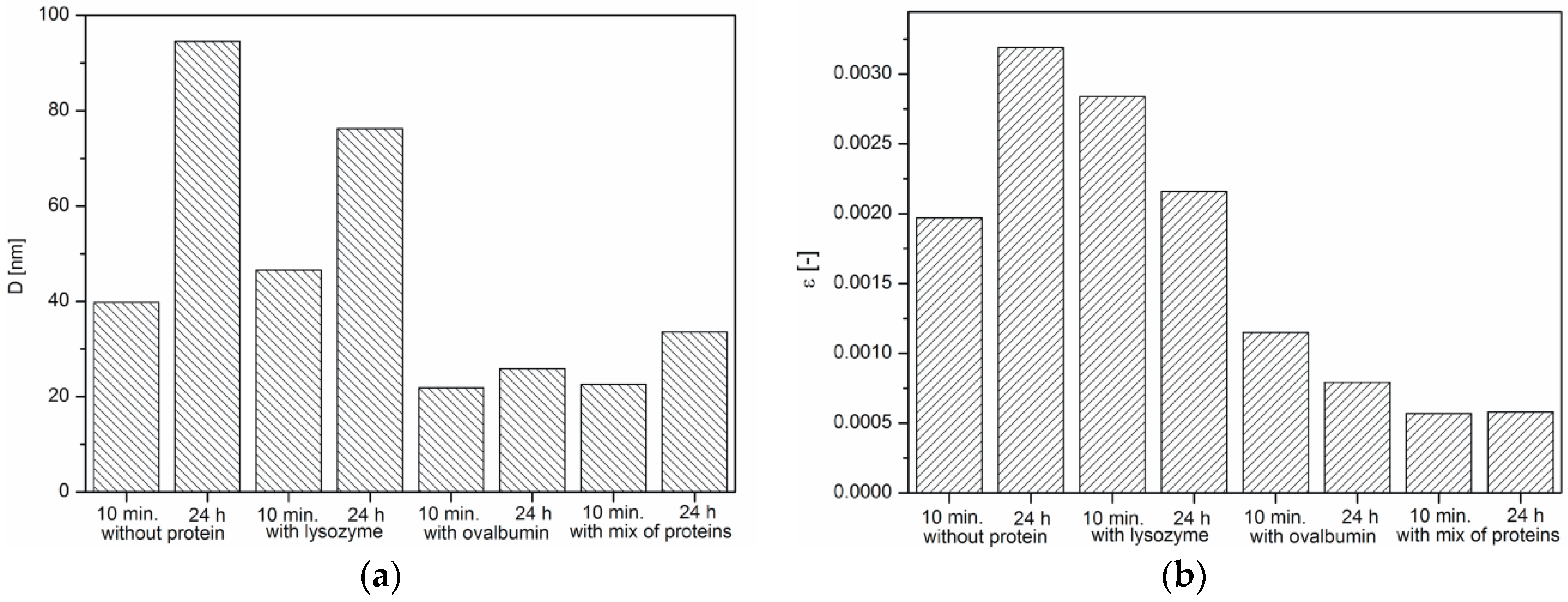
2.3. XRD Analysis of the Calcium Carbonate Crystals
- β—half-width of the (FWHM) peak (rad),
- φ—the angle of the highlight for the given band interference (rad),
- k—the Scherrer constant (the 0.9 value),
- λ—the wavelength of the X-ray beam (nm),
- D—the average crystallite size (nm),
- ε—microstrain.
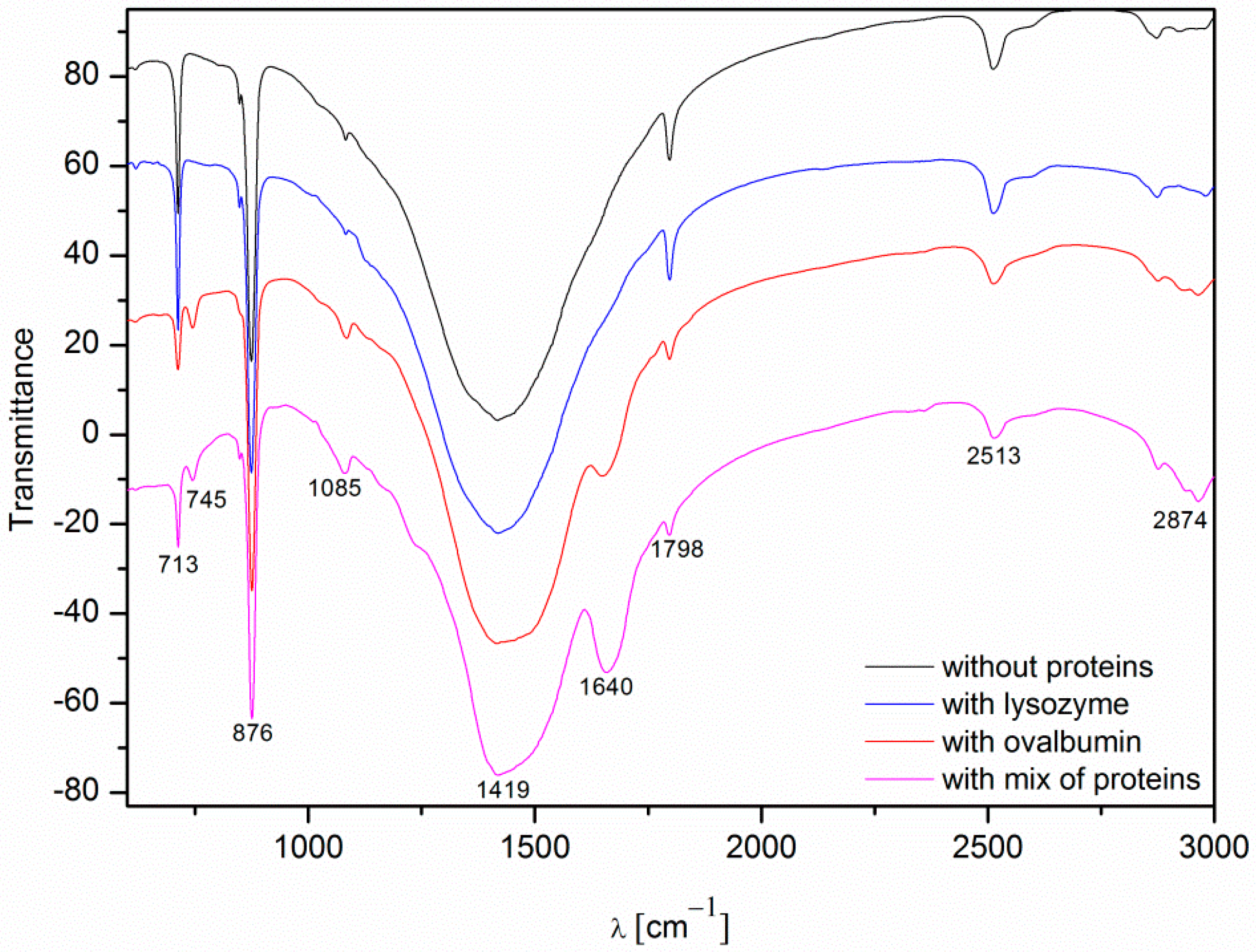
2.4. FTIR Analysis of Calcium Carbonate Crystals
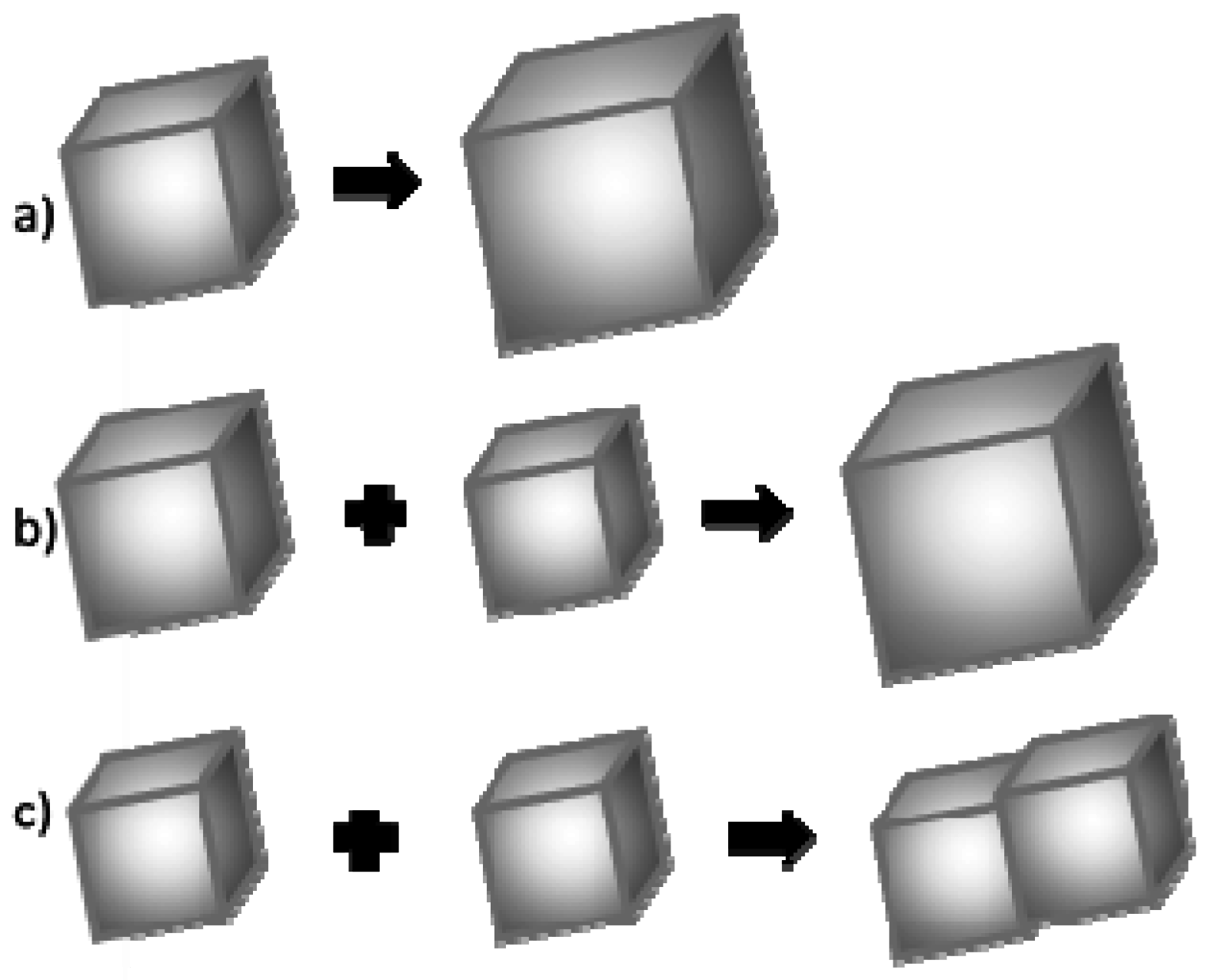
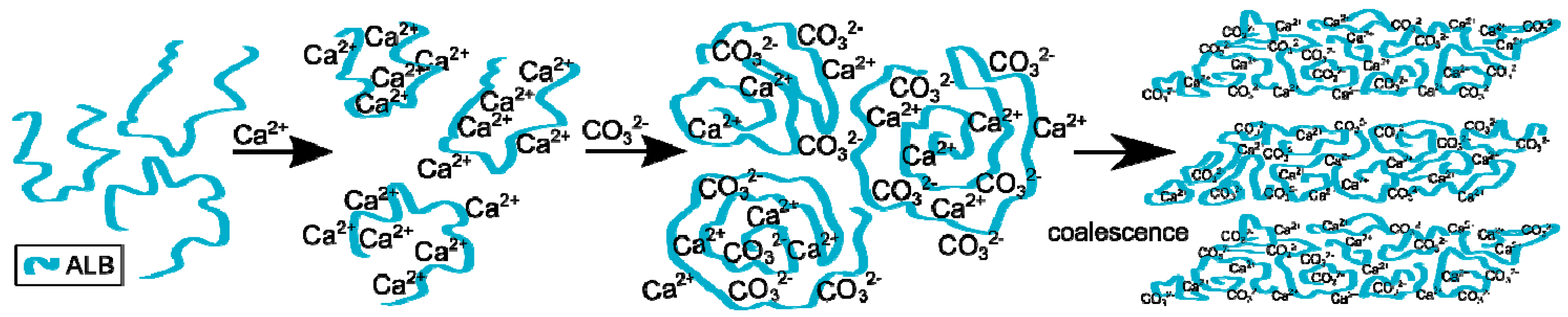
2.5. Mechanism of CaCO3 Crystallization
- (a)
- Association of more atoms/ions to the existing crystal;
- (b)
- Ostwald ripening, i.e., dissolution of smaller crystal grains and growth of larger ones;
- (c)
- Coalescence.
3. Experimental
3.1. Materials
3.2. Methods
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zheng, L.; Hu, Y.; Ma, Y.; Zhou, Y.; Nie, F.; Liu, X.; Pei, C. Egg-white-mediated crystallization of calcium carbonate. J. Cryst. Growth 2012, 361, 217–224. [Google Scholar] [CrossRef]
- Njegić-Džakula, B.; Reggi, M.; Falini, G.; Weber, I.; Brecevic, L.; Kralj, D. The influence of a protein fragment extracted from abalone shell green layer on the precipitation of calcium carbonate polymorphs in aqueous media. Croat. Chem. Acta 2013, 86, 39–47. [Google Scholar] [CrossRef]
- Mann, S.G. Biomineralization: Principles and Concepts in Bioinorganic Materials Chemistry; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Wang, X.; Kong, R.; Pan, X.; Xu, H.; Xia, D.; Shan, H.; Lu, J.R. Role of ovalbumin in the Stabilization of Metastable Vaterite in Calcium Carbonate Biomineralization. J. Phys. Chem. B 2009, 113, 8975–8982. [Google Scholar] [CrossRef] [PubMed]
- Meldrum, F.C.; Cölfen, H. Controlling mineral morphologies and structures in biological and synthetic systems. Chem. Rev. 2008, 108, 4332–4432. [Google Scholar] [CrossRef] [PubMed]
- Fakhrullin, R.F.; Bikmullin, A.G.; Nurgaliev, D.K. Magnetically responsive calcium carbonate microcrystal. ACS Appl. Mater. Interfaces 2009, 1, 1847–1851. [Google Scholar] [CrossRef] [PubMed]
- Park, W.K.; Ko, S.; Lee, S.W.; Cho, K.; Ahn, J.; Han, C. Effects of magnesium chloride and organic additives on the synthesis of aragonite precipitated calcium carbonate. J. Cryst. Growth 2008, 310, 2593–2601. [Google Scholar] [CrossRef]
- Declet, A.; Reyes, E.; Suárez, O.M. Calcium carbonate precipitation: A review of the carbonate crystallization process and applications in bioinspired composites. Rev. Adv. Mater. Sci. 2016, 44, 87–107. [Google Scholar]
- Li, W.; Liu, L.; Chen, W.; Yu, L.; Li, W.; Yu, H. Calcium carbonate precipitation and crystal morphology induced by microbial carbonic anhydrase and other biological factors. Process. Biochem. 2010, 45, 1017–1021. [Google Scholar] [CrossRef]
- Kirboga, S.; Öner, M. Investigation of calcium carbonate precipitation in the presence of carboxymethyl inulin. CrystEngComm 2013, 15, 3678–3686. [Google Scholar] [CrossRef]
- Feng, J.; Wu, G.; Qing, C. Biomimetic synthesis of hollow calcium carbonate with the existence of the agar matrix and bovine serum albumin. Mater. Sci. Eng. C 2016, 58, 409–411. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Hernández, A.; Rodríguez-Navarro, A.B.; Gómez-Morales, J.; Jiménez-Lopez, C.; Nys, Y.; García-Ruiz, J.M. Influence of Model Globular Proteins with Different Isoelectric Points on the Precipitation of Calcium Carbonate. Cryst. Growth Des. 2008, 8, 1495–1502. [Google Scholar] [CrossRef]
- Hu, Y.; Ma, Y.; Zhou, Y.; Nie, F.; Duan, X.; Pei, C. Hen eggwhite-mediated stack crystallization of calcium carbonate. J. Cryst. Growth 2010, 312, 831–836. [Google Scholar] [CrossRef]
- Yang, H.; Yao, W.; Yang, L.; Ma, X.; Wang, H.; Ye, F.; Wong, K. The self-assembly of CaCO3 crystals in the presence of protein. J. Cryst. Growth 2009, 311, 2682–2688. [Google Scholar] [CrossRef]
- Manoli, F.; Kanakis, J.; Malkaj, P.; Dalas, E. The effect of aminoacids on the crystal growth of calcium carbonate. J. Cryst. Growth 2002, 236, 363–370. [Google Scholar] [CrossRef]
- Sadowski, Z.; Polowczyk, I.; Frąckowiak, A.; Koźlecki, T.; Chibowski, S. Bioinspired synthesis of calcium carbonate colloid particles. Physicochem. Probl. Miner. Process. 2010, 44, 205–214. [Google Scholar]
- Polowczyk, I.; Bastrzyk, A.; Koźlecki, T.; Sadowski, Z. Calcium carbonate mineralization. Part 1: The effect of poly(ethylene glycol) concentration of the formation of precipitate. Physicochem. Probl. Miner. Process. 2013, 49, 631–639. [Google Scholar]
- Boyjoo, Y.; Pareek, V.K.; Liu, J. Synthesis of micro and nano-sized calcium carbonate particles and their applications. J. Mater. Chem. A 2014, 2, 14270–14288. [Google Scholar] [CrossRef]
- Rodríguez-Navarro, A.B.; Marie, P.; Nys, Y.; Hincke, M.T.; Gautron, J. Amorphous calcium carbonate controls avian eggshell mineralization: A new paradigm for understanding rapid eggshell calcification. J. Struct. Biol. 2015, 190, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Lopez, C.; Rodriguez-Navarro, A.; Dominguez-Vera, J.M.; Garcia-Ruiz, J.M. Influence of lysozyme on the precipitation of calcium carbonate: A kinetic and morphologic study. Geochim. Cosmochim. Acta 2003, 67, 1667–1676. [Google Scholar] [CrossRef]
- Wang, X.; Sun, H.; Xia, Y.; Chen, C.; Xu, H.; Shan, H.; Lu, J.R. Lysozyme mediated calcium carbonate mineralization. J. Colloid Interface Sci. 2009, 332, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Ma, Y.; Cölfen, H. Biomimetic mineralization. J. Matter. Chem. 2007, 17, 415–449. [Google Scholar] [CrossRef]
- Lakshminarayanan, R.; Loh, X.J.; Gayathri, S.; Sindhu, S.; Banerjee, Y.; Kini, R.M.; Valiyaveettil, S. Formation of transient amorphous calcium carbonate precursor in quail eggshell mineralization: An in vitro study. Biomacromolecules 2006, 7, 3202–3209. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Wang, M.; Wang, X.; Wu, C.; Xu, H.; Lu, J.R. Crystal Growth of Calcite Mediated by Ovalbumin and Lysozyme: Atomic Force Microscopy Study. Cryst. Growth Des. 2013, 13, 1583–1589. [Google Scholar] [CrossRef]
- Voinescu, A.E.; Touraud, D.; Lecker, A.; Pfitzner, A.; Kunz, W.; Ninham, B.W. Mineralization of CaCO3 in the presence of egg white lysozyme. Langmuir 2007, 23, 12269–12274. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Dai, S.; Hu, B.; Du, Z. Effect of Langmuir monolayer of bovine serum albumin protein on the morphology of calcium carbonate. Mater. Sci. Eng. 2009, 29, 1998–2002. [Google Scholar] [CrossRef]
- Faatz, M.; Gröhn, F.; Wegner, G. Amorphous calcium carbonate: Synthesis and potential intermediate in biomineralization. Adv. Mater. 2004, 16, 996–1000. [Google Scholar] [CrossRef]
- Hernández-Hernández, A.; Vidal, M.L.; Gómez-Morales, J.; Rodríguez-Navarro, A.B.; Labas, V.; Gautron, J.; Nys, Y.; García Ruiz, J.M. Influence of eggshell matrix proteins on the precipitation of calcium carbonate (CaCO3). J. Cryst. Growth 2008, 310, 1754–1759. [Google Scholar] [CrossRef]
- Mengual, O.; Meunier, G.; Cayré, I.; Puech, K.; Snabre, P. TURBISCAN MA 2000: Multiple light scattering measurement for concentrated emulsion and suspension instability analysis. Talanta 1999, 50, 445–456. [Google Scholar] [CrossRef]
- Pipich, V.; Balz, M.; Wolf, S.E.; Tremel, W.; Schwahn, D. Nucleation and growth of CaCO3 mediated by the egg-white protein ovalbumin: A time-resolved in situ study using small-angle neutron scattering. J. Am. Chem. Soc. 2008, 130, 6879–6892. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.E.; Leiterer, J.; Pipich, V.; Barrea, R.; Emmerling, F.; Tremel, W. Strong stabilization of amorphous calcium carbonate emulsion by ovalbumin: Gaining insight into the mechanism of ‘polymer-induced liquid precursor’ processes. J. Am. Chem. Soc. 2011, 133, 12642–12649. [Google Scholar] [CrossRef] [PubMed]
- Song, S.M.; Kim, I.H. Biomineralization of calcium carbonate by adding aspartic acid and lysozyme. Korean J. Chem. Eng. 2011, 28, 1749–1753. [Google Scholar] [CrossRef]
- Kim, S.; Park, C.B. Dopamine-Induced Mineralization of Calcium Carbonate Vaterite Microspheres. Langmuir 2010, 26, 14730–14736. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liu, F.; Zhao, S.; Su, Y.; Wang, D.; Shen, Q. Crystallization and oriented attachment of monohydrocalcite and its crystalline phase transformation. CrystEngComm 2013, 15, 509–515. [Google Scholar] [CrossRef]
- Williamson, G.K.; Hall, W.H. X-ray line broadening from filed aluminium and wolfram. Acta Metall. 1953, 1, 22–31. [Google Scholar] [CrossRef]
- Kim, Y.; Semsarilar, M.; Carloni, J.D.; Cho, K.R.; Kulak, A.N.; Polishchuk, I.; Hendley, C.T.; Smeets, P.J.M.; Fielding, L.A.; Pokroy, B.; et al. Structure and properties of nanocomposites formed by the occlusion of block copolymer worms and vesicles within calcite crystals. Adv. Funct. Mater. 2016, 26, 1382–1392. [Google Scholar] [CrossRef]
- Kim, Y.; Schenk, A.S.; Ihli, J.; Kulak, A.N.; Hetherington, N.B.J.; Tang, C.C.; Schmahl, W.W.; Griesshaber, E.; Hyett, G.; Meldrum, F.C. A critical analysis of calcium carbonate mesocrystals. Nat. Commun. 2014. [Google Scholar] [CrossRef] [PubMed]
- Akdogan, E.K.; Leonard, M.R.; Safari, A. Size effects in ferroelectric ceramics. In Handbook of Low and High Dielectric Constant Materials and Their Application; Nalwa, H.S., Ed.; Academic Press: San Diego, CA, USA, 1999; Volume 2, pp. 60–112. [Google Scholar]
- Menahem, T.; Mastai, Y. Controlled crystallization of calcium carbonate superstructures in macroemulsions. J. Cryst. Growth 2008, 310, 3552–3556. [Google Scholar] [CrossRef]
- Xu, X.; Zhao, Y.; Lai, Q.; Hao, Y. Effect of polyethylene glycol on phase and morphology of calcium carbonate. J. Appl. Polym. Sci. 2011, 119, 319–324. [Google Scholar] [CrossRef]
- Yang, L.; She, L.; Zhou, J.; Cao, Y.; Ma, X. Interaction of lysozyme during calcium carbonate precipitation at supramolecular level. Inorg. Chem. Commun. 2006, 9, 164–166. [Google Scholar] [CrossRef]
- Ichikawa, K.; Shimomura, N.; Yamada, M.; Ohkubo, N. Control of calcium carbonate polymorphism and morphology through biomimetic mineralization by means of nanotechnology. Chem. Eur. J. 2003, 9, 3235–3241. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Han, H.; Ding, S.; Li, Z.; Kong, X. Effect of silk sericin on morphology and structure of calcium carbonate crystal. Front. Mater. Sci. 2013, 7, 177–183. [Google Scholar] [CrossRef]
- Zhu, W.; Lin, J.; Cai, C.; Lu, Y. Biomimetic mineralization of calcium carbonate mediated by a polypeptide-based copolymer. J. Mater. Chem. B 2013, 1, 841–849. [Google Scholar] [CrossRef]
- Matsumoto, T.; Chiba, J.; Inoue, H. Effect of pH on colloidal properties of native ovalbumin aqueous systems. Colloid Polym. Sci. 1992, 270, 687–693. [Google Scholar] [CrossRef]
- Chibowski, E.; Hołysz, L.; Szcześ, A. Time dependent changes in zeta potential of freshly precipitated calcium carbonate. Colloids Surf. A 2003, 222, 41–54. [Google Scholar] [CrossRef]
- Bönnemann, H.; Richards, R.M. Nanoscopic metal particles—Synthetic methods and potential applications. Eur. J. Inorg. Chem. 2001, 2001, 2455–2480. [Google Scholar] [CrossRef]
- De Yoreo, J.J.; Gilbert, P.U.P.A.; Sommerdijk, N.A.J.M.; Penn, R.L.; Whitelam, S.; Joester, D.; Zhang, H.; Rimer, J.D.; Navrotsky, A.; Banfield, J.F.; et al. Crystallization by particle attachment in synthetic, biogenic, and geologic environments. Science 2015, 349, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, R.; El-Sayed, M. Catalysis with transition metal nanoparticles in colloidal solution: Nanoparticle shape dependence and stability. J. Phys. Chem. B 2005, 109, 12663–12676. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Rieger, J.; Huber, K. Analysis of the nucleation and growth of amorphous CaCO3 by means of time-resolved static light scattering. Langmuir 2008, 24, 8262–8271. [Google Scholar] [CrossRef] [PubMed]
- Wojtas, M.; Hołubowicz, R.; Poznar, M.; Maciejewska, M.; Ożyhar, A.; Dobryszycki, P. Calcium ion binding properties and the effect of phosphorylation on the intrinsically disordered starmaker protein. Biochemistry 2015, 54, 6525–6534. [Google Scholar] [CrossRef] [PubMed]
- Damodaran, S.; Anand, K.; Razumovsky, L. Competitive adsorption of egg white proteins at the air-water interface: Direct evidence for electrostatic complex formation between lysozyme and other egg proteins at the interface. J. Agric. Food Chem. 1998, 46, 872–876. [Google Scholar] [CrossRef]
- Curtis, R.A.; Lue, L. A molecular approach to bioseparations: Protein–protein and protein–salt interactions. Chem. Eng. Sci. 2006, 61, 907–923. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wavenumber [cm−1] | Assignment |
|---|---|
| ~713 | In-plane deformation mode of CO32− in calcite |
| ~745 | In-plane deformation mode of CO32− in vaterite |
| ~876 | Out-of-plane deformation mode of CO32− |
| ~1085 | Symmetric C–O stretching mode |
| ~1420 | Asymmetric C–O stretching mode |
| ~1640 | C=O stretching mode in I amide |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polowczyk, I.; Bastrzyk, A.; Fiedot, M. Protein-Mediated Precipitation of Calcium Carbonate. Materials 2016, 9, 944. https://doi.org/10.3390/ma9110944
Polowczyk I, Bastrzyk A, Fiedot M. Protein-Mediated Precipitation of Calcium Carbonate. Materials. 2016; 9(11):944. https://doi.org/10.3390/ma9110944
Chicago/Turabian StylePolowczyk, Izabela, Anna Bastrzyk, and Marta Fiedot. 2016. "Protein-Mediated Precipitation of Calcium Carbonate" Materials 9, no. 11: 944. https://doi.org/10.3390/ma9110944
APA StylePolowczyk, I., Bastrzyk, A., & Fiedot, M. (2016). Protein-Mediated Precipitation of Calcium Carbonate. Materials, 9(11), 944. https://doi.org/10.3390/ma9110944