
Approaches to Parameter Estimation from Model Neurons and Biological Neurons


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Model Neurons: Lessons Learnt from Well-Posed Models
2.1. Choosing Neuron Models
2.2. Observability
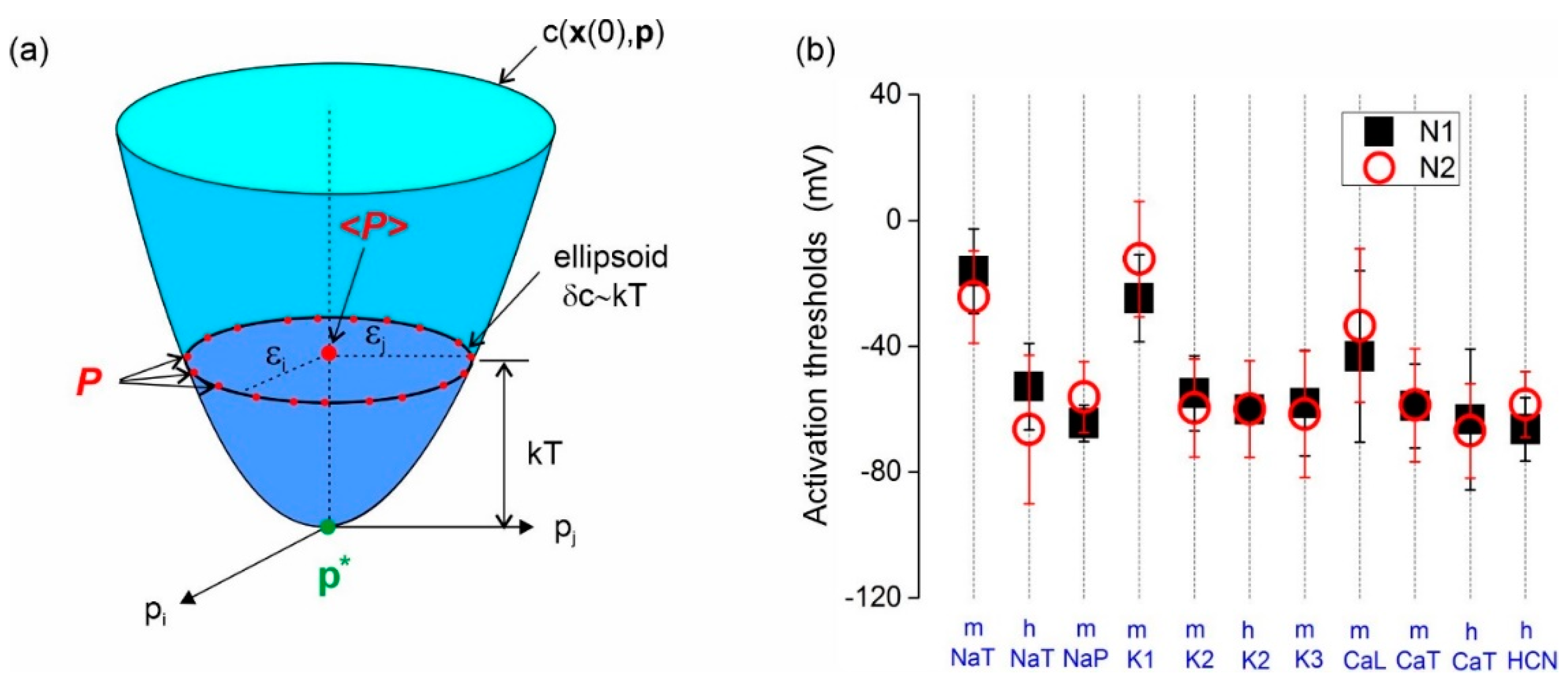
2.3. Identifiability
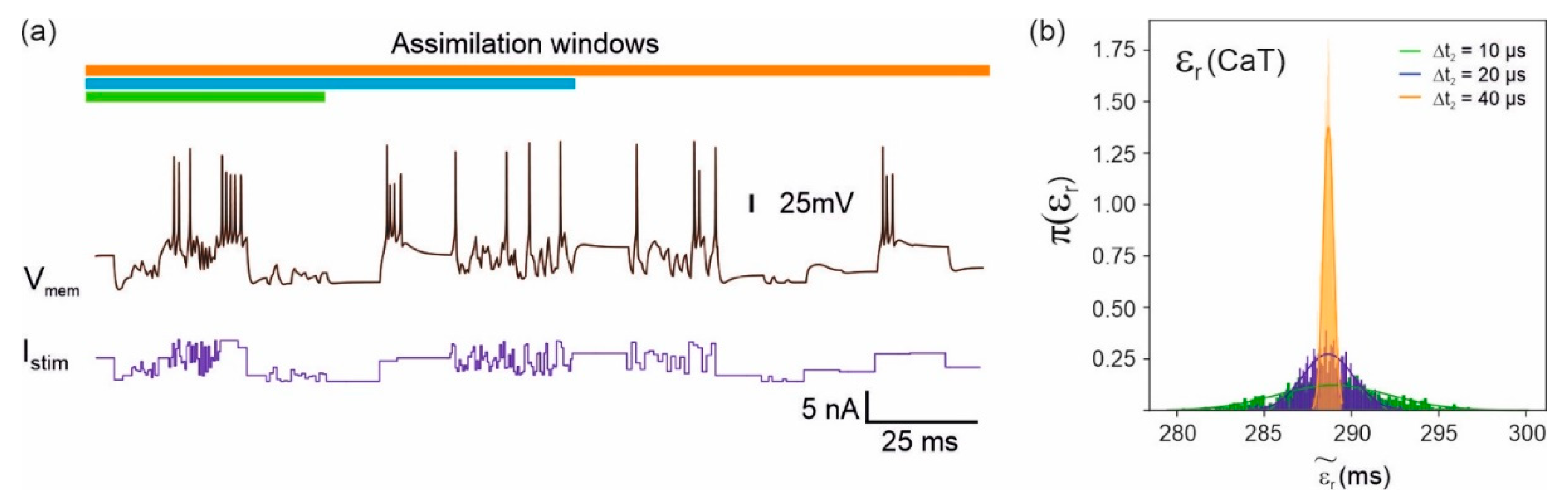
2.4. Noise and Effect of Assimilation Window over Parameter Distribution Adaptive Sampling
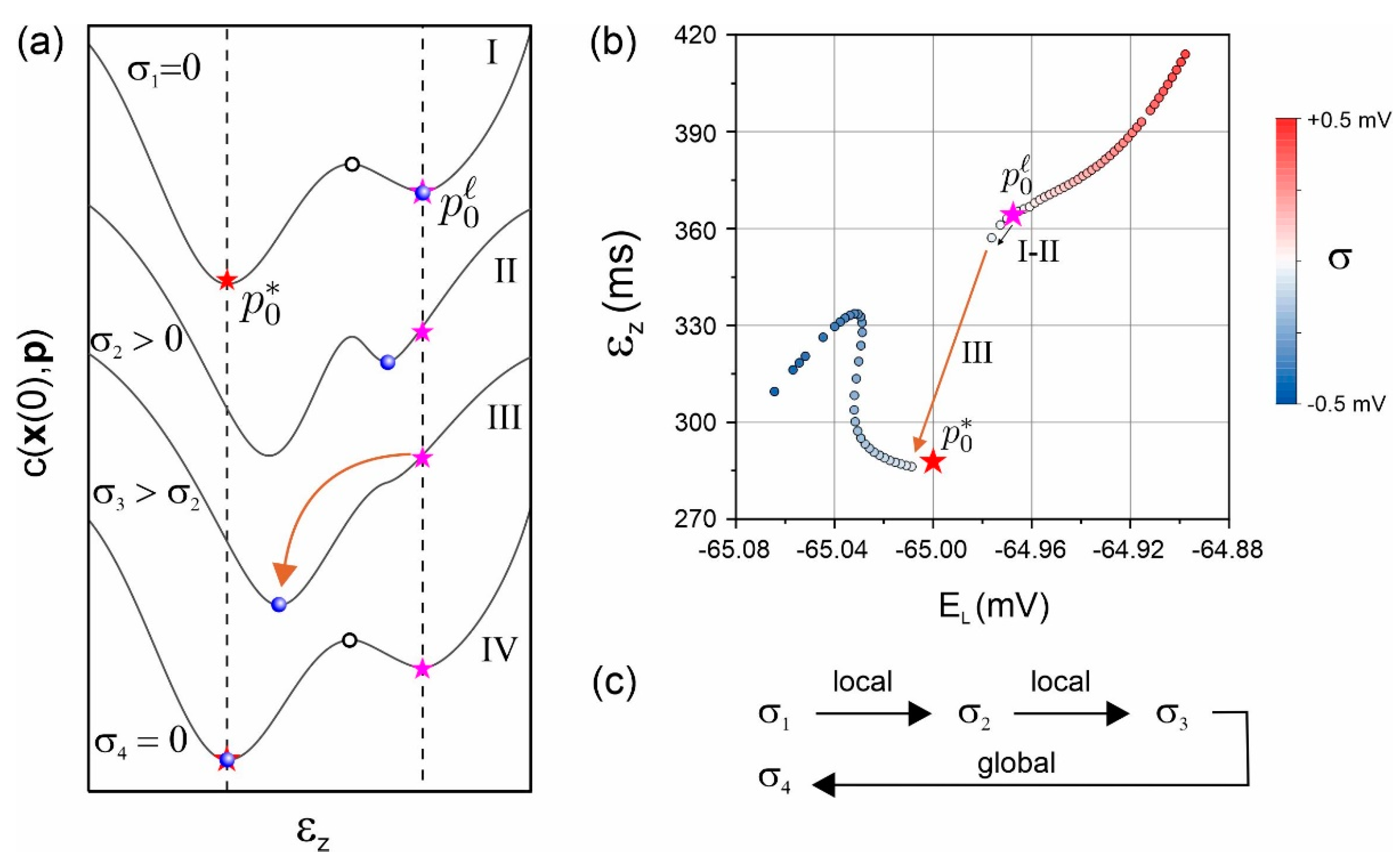
2.5. Stochastic Regularization of Convergence to Local Minima Vicinal to the Global Minimum
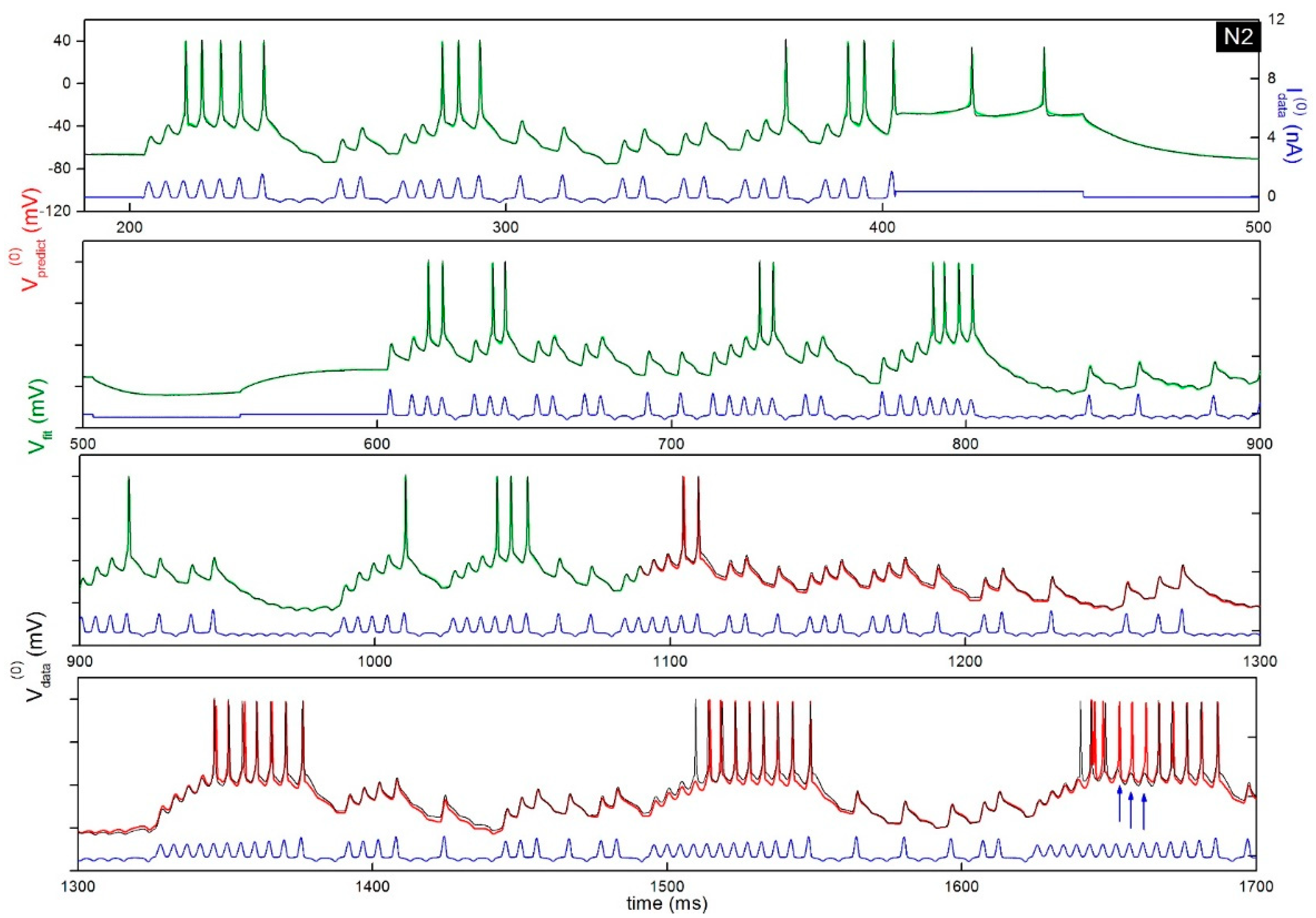
3. Biological Neurons: Estimating Parameters from Guessed Models
3.1. Including Model Error in the Cost Function
3.2. Supplementing Gradient Descent with Statistical Inference
3.3. Aggregate State Variables
4. Discussion
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pathak, J.; Hunt, B.; Girvan, M.; Lu, Z.; Ott, E. Model-Free Prediction of Large Spatiotemporally Chaotic Systems from Data: A Reservoir Computing Approach. Phys. Rev. Lett. 2018, 120, 024102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, D.; Schrittwieser, J.; Simonyan, K.; Antonoglou, I.; Huang, A.; Guez, A.; Hubert, T.; Baker, L.; Lai, M.; Bolton, A.; et al. Mastering the game of Go without human knowledge. Nature 2017, 550, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Kalnay, E. Atmospheric Modeling, Data Assimilation and Predictability; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Tabeart, J.M.; Dance, S.L.; Haben, S.A.; Lawless, A.S.; Nichols, N.K.; Waller, J.A. The conditioning of least-squares problems in variational data assimilation. Numer. Linear Algebra Appl. 2018, 25, e2165. [Google Scholar] [CrossRef]
- Abarbanel, H.D.I. Predicting the Future: Completing Models of Observed Complex Systems; Springer: Heidelberg, Germany, 2013. [Google Scholar]
- Abarbanel, H.D.I. The Statistical Physics of Data Assimilation and Machine Learning; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar] [CrossRef]
- Meliza, C.D.; Kostuk, M.; Huang, H.; Nogaret, A.; Margoliash, D.; Abarbanel, H.D.I. Estimating parameters and predicting membrane voltages with conductance-based neuron models. Biol. Cybern. 2014, 108, 495–516. [Google Scholar] [CrossRef]
- Nogaret, A.; Meliza, C.D.; Margoliash, D.; Abarbanel, H.D.I. Automatic Construction of Predictive Neuron Models through Large Scale Assimilation of Electrophysiological Data. Sci. Rep. 2016, 6, 32749. [Google Scholar] [CrossRef] [Green Version]
- Kostuk, M.; Toth, B.A.; Meliza, C.D.; Margoliash, D.; Abarbanel, H.D.I. Dynamical estimation of neuron and network properties II: Path integral Monte Carlo methods. Biol. Cybern. 2012, 106, 155–167. [Google Scholar] [CrossRef]
- Toth, B.A.; Kostuk, M.; Meliza, C.D.; Margoliash, D.; Abarbanel, H.D.I. Dynamical estimation of neuron and network properties I: Variational methods. Biol. Cybern. 2011, 105, 217–237. [Google Scholar] [CrossRef]
- Taylor, J.D.; Winnall, S.; Nogaret, A. Estimation of neuron parameters from imperfect observations. PLOS Comput. Biol. 2020, 16, e1008053. [Google Scholar] [CrossRef]
- Hartoyo, A.; Casdusch, P.J.; Lilley, D.T.J.; Hicks, D.J. Parameter estimation and identifiability in a neural population model for electro-cortical activity. PLoS Comp. Biol. 2019, 15, e1006694. [Google Scholar] [CrossRef] [Green Version]
- Brookings, T.; Goeritz, M.L.; Marder, E. Automatic parameter estimation of multicompartmental neuron models via minimization of trace error with control adjustment. J. Neurophysiol. 2014, 112, 2332–2348. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, E. Statistical data assimilation for estimating electrophysiology simultaneously with connectivity within a biological neuronal network. Phys. Rev. E 2020, 101, 012415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, E.; Abarbanel, H.D.I. Model of the songbird nucleus HVC as a network of central pattern generators. J. Neurophysiol. 2016, 116, 2405–2419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prinz, A.A.; Billimoria, C.P.; Marder, E. Alternative to hand-tuning conductance-based models: Constriction and analysis of databases of model neurons. J. Neurophysiol. 2003, 90, 4015. [Google Scholar] [CrossRef] [PubMed]
- Gouwens, N.W.; Sorensen, S.A.; Baftizadeh, F.; Budzillo, A.; Lee, B.R.; Jarsky, T.; Alfiler, L.; Baker, K.; Barkan, E.; Berry, K.; et al. Integrated Morphoelectric and Transcriptomic Classification of Cortical GABAergic Cells. Cell 2020, 183, 953.e19. [Google Scholar] [CrossRef] [PubMed]
- Griffin, M.; Khan, R.; Basu, S.; Smith, S. Ion Channels as Therapeutic Targets in High Grade Gliomas. Cancers 2020, 12, 3068. [Google Scholar] [CrossRef] [PubMed]
- Molenaar, R.J. Ion Channels in Glioblastoma. ISRN Neurol. 2011, 2011, 590249. [Google Scholar] [CrossRef] [Green Version]
- Chakroborty, S.; Stutzmann, G.E. Calcium channelopathies and Alzheimer’s disease: Insight into therapeutic success and failures. Eur. J. Pharmacol. 2014, 739, 83–95. [Google Scholar] [CrossRef]
- Huys, Q.J.M.; Ahrens, M.B.; Paninski, L. Efficient Estimation of Detailed Single-Neuron Models. J. Neurophysiol. 2006, 96, 872–890. [Google Scholar] [CrossRef]
- Eiben, A.E.; Smith, J.E. Introduction to Evolutionary Computing; Springer: Heidelberg, Germany, 2003. [Google Scholar]
- Druckmann, S.; Banitt, Y.; A Gidon, A.; Schürmann, F.; Markram, H.; Segev, I. A novel multiple objective optimization framework for constraining conductance-based neuron models by experimental data. Front. Neurosci. 2007, 1, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, R.; Tsubo, Y.; Shinomoto, S. Made-to-order spiking neuron model equipped with a multi-timescale adaptive threshold. Front. Comput. Neurosci. 2009, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Marasco, A.; Limongiello, A.; Migliore, M. Fast and accurate low-dimensional reduction of biophysically detailed neuron models. Sci. Rep. 2012, 2, srep00928. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; De Schutter, E. Complex Parameter Landscape for a Complex Neuron Model. PLoS Comput. Biol. 2006, 2, e94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhry, L.; Pace, M.; Saïghi, S. Global parameter estimation of an Hodgkin–Huxley formalism using membrane voltage recordings: Application to neuro-mimetic analog integrated circuits. Neurocomputing 2012, 81, 75–85. [Google Scholar] [CrossRef]
- Hendrickson, E.B.; Edgerton, J.R.; Jaeger, D. The use of automated parameter searches to improve ion channel kinetics for neural modeling. J. Comput. Neurosci. 2011, 31, 329–346. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.; Eberhardt, R. Particle swarm optimization. In Proceedings of the ICNN’95-International Conference On Neural Networks IEEE, Perth, Australia, 27 November–1 December 1995; pp. 1942–1948. [Google Scholar]
- Wächter, A.; Biegler, L.T. On the implementation of an interior-point filter line-search algorithm for large-scale nonlinear programming. Math. Program. 2005, 106, 25–57. [Google Scholar] [CrossRef]
- Boyd, S.; Vanderberghe, L. Convex Optimization; Cambridge University Press: Cambridge, UK, 2004. [Google Scholar]
- Vavoulis, D.V.; Straub, V.; Aston, J.A.D.; Feng, J. A Self-Organizing State-Space-Model Approach for Parameter Estimation in Hodgkin-Huxley-Type Models of Single Neurons. PLoS Comput. Biol. 2012, 8, e1002401. [Google Scholar] [CrossRef]
- Marder, E.; Taylor, A.L. Multiple models to capture the variability in biological neurons and networks. Nat. Neurosci. 2011, 14, 133–138. [Google Scholar] [CrossRef]
- A Prinz, A.; Bucher, D.; Marder, E. Similar network activity from disparate circuit parameters. Nat. Neurosci. 2004, 7, 1345–1352. [Google Scholar] [CrossRef]
- Goaillard, J.-M.; Taylor, A.L.; Schulz, D.; Marder, E. Functional consequences of animal-to-animal variation in circuit parameters. Nat. Neurosci. 2009, 12, 1424–1430. [Google Scholar] [CrossRef] [Green Version]
- Marder, E.; Goaillard, J.-M. Variability, compensation and homeostasis in neuron and network function. Nat. Rev. Neurosci. 2006, 7, 563–574. [Google Scholar] [CrossRef]
- Lederman, D.; Patel, R.; Itani, O.; Rotstein, H.G. Parameter Estimation in the Age of Degeneracy and Unidentifiability. Mathematics 2022, 10, 170. [Google Scholar] [CrossRef]
- O’Leary, T.; Sutton, A.C.; Marder, E. Computational models in the age of large data sets. Curr. Opin. Neurobiol. 2015, 32, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutenkunst, R.N.; Waterfall, J.; Casey, F.P.; Brown, K.S.; Myers, C.R.; Sethna, J.P. Universally Sloppy Parameter Sensitivities in Systems Biology Models. PLOS Comput. Biol. 2007, 3, 1871–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushinsky, D.; Morozova, E.O.; Marder, E. In vivo effects of temperature on the heart and pyloric rhythms in the crab, Cancer borealis. J. Exp. Biol. 2019, 222, jeb199190. [Google Scholar] [CrossRef] [Green Version]
- Haley, J.A.; Hampton, D.; Marder, E. Two central pattern generators from the crab, Cancer borealis respond robustly and differentially to extreme extracellular pH. eLife 2018, 7, e41977. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar] [CrossRef]
- Huguenard, J.R.; Mc Cormick, D.A. Simulation of the currents involved in rhythmic oscillations in thalamic neurons. J. Neurophysiol. 1992, 68, 1373. [Google Scholar] [CrossRef]
- Otopalik, A.G.; Sutton, A.C.; Banghart, M.; Marder, E. When complex neuronal structures may not matter. eLife 2017, 6, 2066. [Google Scholar] [CrossRef] [Green Version]
- Lilacci, G.; Khammash, M. Parameter estimation and model selection in computational biology. PLoS Comp. Biol. 2012, 6, e1000696. [Google Scholar] [CrossRef] [Green Version]
- Kahl, D.; Kschischo, M. Searching for Errors in Models of Complex Dynamic Systems. Front. Physiol. 2021, 11, 612590. [Google Scholar] [CrossRef]
- Markram, H.; Sakman, B. Calcium transients in dendrites of neocortical neurons evoked by single threshold excitatory postsynaptic potentials via low-voltage-activated calcium channels. Proc. Nat. Acad. Sci. USA 1994, 91, 5207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Negro, C.A.; Morgado-Valle, C.; Hayes, J.A.; Mackay, D.D.; Pace, R.W.; Crowder, E.A.; Feldman, J.L. Sodium and Calcium Current-Mediated Pacemaker Neurons and Respiratory Rhythm Generation. J. Neurosci. 2005, 25, 446–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kastellakis, G.; Silva, A.J.; Poirazi, P. Linking Memories across Time via Neuronal and Dendritic Overlaps in Model Neurons with Active Dendrites. Cell Rep. 2016, 17, 1491–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spruston, N.; Schiller, Y.; Stuart, G.; Sakmann, B. Activity-Dependent Action Potential Invasion and Calcium Influx into Hippocampal CA1 Dendrites. Science 1995, 268, 297–300. [Google Scholar] [CrossRef]
- Abu-Hassan, K.; Taylor, J.D.; Morris, P.G.; Donati, E.; Bortolotto, Z.A.; Indiveri, G.; Paton, J.F.R.; Nogaret, A. Optimal neuron models. Nat. Commun. 2019, 10, 5309. [Google Scholar] [CrossRef]
- Takens, F. Detecting Strange Attractors in Turbulence in Dynamic Systems in Turbulence; Springer: Warwick, UK, 1981; pp. 366–381. [Google Scholar]
- Aeyels, D. Generic Observability of Differentiable Systems. SIAM J. Control Optim. 1981, 19, 595–603. [Google Scholar] [CrossRef]
- Parlitz, U.; Schumann-Bischoff, J.; Luther, S. Local observability of state variables and parameters in nonlinear modeling quantified by delay reconstruction. Chaos Interdiscip. J. Nonlinear Sci. 2014, 24, 24411. [Google Scholar] [CrossRef] [Green Version]
- Parlitz, U.; Schumann-Bischoff, J.; Luther, S. Quantifying uncertainty in state and parameter estimation. Phys. Rev. E 2014, 89, 050902. [Google Scholar] [CrossRef] [Green Version]
- Parlitz, U.; Merkwirth, C. Prediction of Spatiotemporal Time Series Based on Reconstructed Local States. Phys. Rev. Lett. 2000, 84, 1890–1893. [Google Scholar] [CrossRef]
- Schumann-Bischoff, J.; Parlitz, U. State and parameter estimation using unconstrained optimization. Phys. Rev. E 2011, 84, 056214. [Google Scholar] [CrossRef]
- Schumann-Bischoff, J.; Luther, S.; Parlitz, U. Estimability and dependency analysis of model parameters based on delay coordinates. Phys. Rev. E 2016, 94, 032221. [Google Scholar] [CrossRef] [PubMed]
- Letellier, C.; Aguirre, L.; Maquet, J. How the choice of the observable may influence the analysis of nonlinear dynamical systems. Commun. Nonlinear Sci. Numer. Simul. 2006, 11, 555–576. [Google Scholar] [CrossRef]
- Letellier, C.; Aguirre, L.A.; Maquet, J. Relation between observability and differential embeddings for nonlinear dynamics. Phys. Rev. E 2005, 71, 066213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguirre, L.A.; Portes, L.L.; Letellier, C. Observability and synchronization of neuron models. Chaos Interdiscip. J. Nonlinear Sci. 2017, 27, 103103. [Google Scholar] [CrossRef] [Green Version]
- Tabak, J.; Moore, L. Simulation and parameter estimation study of a simple neuronal model of rhythm generation: Role of NMDA and non-NMDA receptors. J. Comput. Neurosci. 1998, 5, 209–235. [Google Scholar] [CrossRef]
- Moye, M.J.; Diekman, C.O. Data Assimilation Methods for Neuronal State and Parameter Estimation. J. Math. Neurosci. 2018, 8, 11. [Google Scholar] [CrossRef]
- Rey, D.; Eldridge, M.; Morone, U.; Abarbanel, H.D.I.; Parlitz, U.; Schumann-Bischoff, J. Using waveform information in nonlinear data assimilation. Phys. Rev. E 2014, 90, 062916. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Rey, D.; Kadakia, N.; Eldridge, M.; Morone, U.I.; Rozdeba, P.; Abarbanel, H.D.I.; Quinn, J.C. Systematic variational method for statistical nonlinear state and parameter estimation. Phys. Rev. E 2015, 92, 052901. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Chen, G.; Cao, J.; Lü, J.; Parlitz, U. Parameter identification of dynamical systems from time series. Phys. Rev. E 2007, 75, 067201. [Google Scholar] [CrossRef]
- Raue, A.; Becker, V.; Klingmüller, U.; Timmer, J. Identifiability and observability analysis for experimental design in nonlinear dynamical models. Chaos Interdiscip. J. Nonlinear Sci. 2010, 20, 045105. [Google Scholar] [CrossRef]
- Kuwana, S.-I.; Tsunekawa, N.; Yanagawa, Y.; Okada, Y.; Kuribayashi, J.; Obata, K. Electrophysiological and morphological characteristics of GABAergic respiratory neurons in the mouse pre-Bötzinger complex. Eur. J. Neurosci. 2006, 23, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.S.; Taylor, J.; Nogaret, A. Local inhibitory networks support up to (N−1)!/lnN2 limit cycles in the presence of electronic noise and heterogeneity. Phys. Rev. Res. 2021, 3, 043097. [Google Scholar] [CrossRef]
- Nogaret, A.; King, A. Inhibition delay increases neural network capacity through Sterling transform. Phys. Rev. E 2018, 97, 030301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, A.S.; Taylor, J.D.; Nogaret, A. Dual Mechanism for the Emergence of Synchronization in Inhibitory Neural Networks. Sci. Rep. 2018, 8, 11431. [Google Scholar] [CrossRef] [Green Version]
- Cover, T.M.; Thomas, J.A. Elements of Information Theory; Wiley: Hoboken, NJ, USA, 1991. [Google Scholar]
- Hermann, R.; Krener, A. Nonlinear controllability and observability. IEEE Trans. Autom. Control 1977, 22, 728–740. [Google Scholar] [CrossRef] [Green Version]
- Villaverde, A.F.; Barreiro, A.; Papachristodoulou, A. Structural Identifiability of Dynamic Systems Biology Models. PLOS Comput. Biol. 2016, 12, e1005153. [Google Scholar] [CrossRef] [Green Version]
- Faisal, A.A.; Selen, L.P.J.; Wolpert, D.M. Noise in the nervous system. Nat. Rev. Neurosci. 2006, 9, 292. [Google Scholar] [CrossRef]
- Blayo, E.; Bouquet, M.; Cosme, E.; Cugliandolo, L.F. Advanced Data Assimilation for Geosciences. Lecture Notes of the Les Houches School of Physics: Special Issue; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Morse, G.; Stanley, K.O. Simple Evolutionary Optimization Can Rival Stochastic Gradient Descent in Neural Networks. Proc. GECCO 2016, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Bottou, L.; Bousquet, O. The tradeoffs of large-scale learning. Adv. Neural Inf. Processing Syst. 2008, 20, 161. [Google Scholar]
- Abarbanel, H.D.I.; Creveling, D.R.; Jeanne, J.M. Estimation of parameters in nonlinear systems using balanced synchronization. Phys. Rev. E 2008, 77, 016208. [Google Scholar] [CrossRef] [Green Version]
- Abarbanel, H.D.I.; Kostuk, M.; Whartenby, W. Data assimilation with regularized nonlinear instabilities. Q. J. R. Meteorol. Soc. 2010, 136, 769–783. [Google Scholar] [CrossRef]
- Morris, P.G.; Taylor, J.D.; Wells, S.A.; Nogaret, A. Prediction of Pharmacologically-Induced Changes in Specific Ion Channels by Assimilation of Neuronal Membrane Voltage Data. UK Patent 2206436.4, 2022. [Google Scholar]
- Deneux, T.; Kaszas, A.; Szalay, G.; Katona, G.; Lakner, T.; Grinvald, A.; Rózsa, B.; Vanzetta, I. Accurate spike estimation from noisy calcium signals for ultra-fast three-dimensional imaging of large neuronal populations in-vivo. Nat. Commun. 2016, 7, 12190. [Google Scholar] [CrossRef] [PubMed]
- Davidović, A.; Chait, R.; Batt, G.; Ruess, J. Parameter inference for stochastic biochemical models from perturbation experiments parallelized at the single cell level. PLoS Comput. Biol. 2022, 18, e1009950. [Google Scholar] [CrossRef]
- Tikhonov, A.N. On the stability of inverse problems. Doklaady Akad. Nauk. SSSR 1943, 39, 195. [Google Scholar]
- Creveling, D.R.; Gill, P.E.; Abarbanel, H.D. State and parameter estimation in nonlinear systems as an optimal tracking problem. Phys. Lett. A 2008, 372, 2640–2644. [Google Scholar] [CrossRef]
- Wang, J.; Breen, D.; Akinin, A.; Broccard, F.D.; Abarbanel, H.D.I.; Cauwenberghs, G. Assimilation of Biophysical Neuronal Dynamics in Neuromorphic VLSI. IEEE Trans. Biomed. Circuits Syst. 2017, 11, 1258–1270. [Google Scholar] [CrossRef]
- O’Callaghan, E.L.; Chauhan, A.S.; Zhao, L.; Lataro, R.M.; Salgado, H.C.; Nogaret, A.; Paton, J.F.R. Utility of a novel biofeedback device for within breadth modulation of heart rate in rats: A quantitative comparison of vagus nerve vs right atrial pacing. Front. Physiol. 2016, 7, 27. [Google Scholar]
- Jiang, S.; Patel, D.C.; Kim, J.; Yang, S.; Mills, W.A.; Zhang, Y.; Wang, K.; Feng, Z.; Vijayan, S.; Cai, W.; et al. Spatially expandable fiber-based probes as a multifunctional deep brain interface. Nat. Commun. 2020, 11, 6115. [Google Scholar] [CrossRef]
- Trautmann, E.M.; O’Shea, D.J.; Sun, X.; Marshel, J.H.; Crow, A.; Hsueh, B.; Vesuna, S.; Cofer, L.; Bohner, G.; Allen, W.; et al. Dendritic calcium signals in rhesus macaque motor cortex drive an optical brain-computer interface. Nat. Commun. 2021, 12, 3689. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nogaret, A. Approaches to Parameter Estimation from Model Neurons and Biological Neurons. Algorithms 2022, 15, 168. https://doi.org/10.3390/a15050168
Nogaret A. Approaches to Parameter Estimation from Model Neurons and Biological Neurons. Algorithms. 2022; 15(5):168. https://doi.org/10.3390/a15050168
Chicago/Turabian StyleNogaret, Alain. 2022. "Approaches to Parameter Estimation from Model Neurons and Biological Neurons" Algorithms 15, no. 5: 168. https://doi.org/10.3390/a15050168
APA StyleNogaret, A. (2022). Approaches to Parameter Estimation from Model Neurons and Biological Neurons. Algorithms, 15(5), 168. https://doi.org/10.3390/a15050168