The Positive Effect of Different 24-epiBL Pretreatments on Salinity Tolerance in Robinia pseudoacacia L. Seedlings


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and 24-epibrassinolide Treatments
2.2. Groups and Salt Stress Treatments
2.3. Photosynthesis
2.4. Estimation of Membrane Stability Index and Relative Water Content
2.5. Observation of Chloroplast Ultrastructure
2.6. Assays for Malondialdehyde and Antioxidant Enzyme Activities
2.7. Assays for Chlorophyll Pigment Concentration
2.8. Isolation of Chloroplasts for Testing
2.9. Statistical Analysis
3. Results
3.1. Photosynthesis
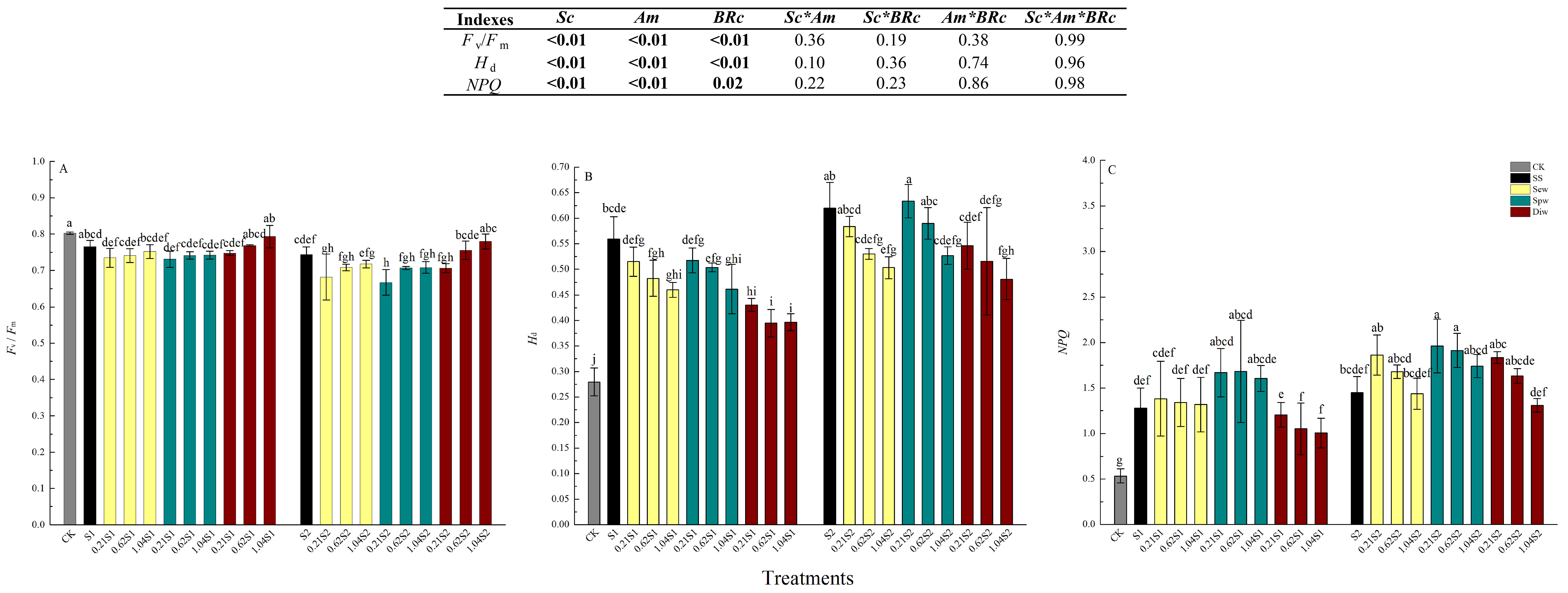
3.2. Chlorophyll Fluorescence
3.3. RWC and MSI
3.4. Photosynthetic Pigments
3.5. MDA, POD, and SOD in Leaves
3.6. Variation in Na+, K+, and Ca2+ Content in Chloroplasts
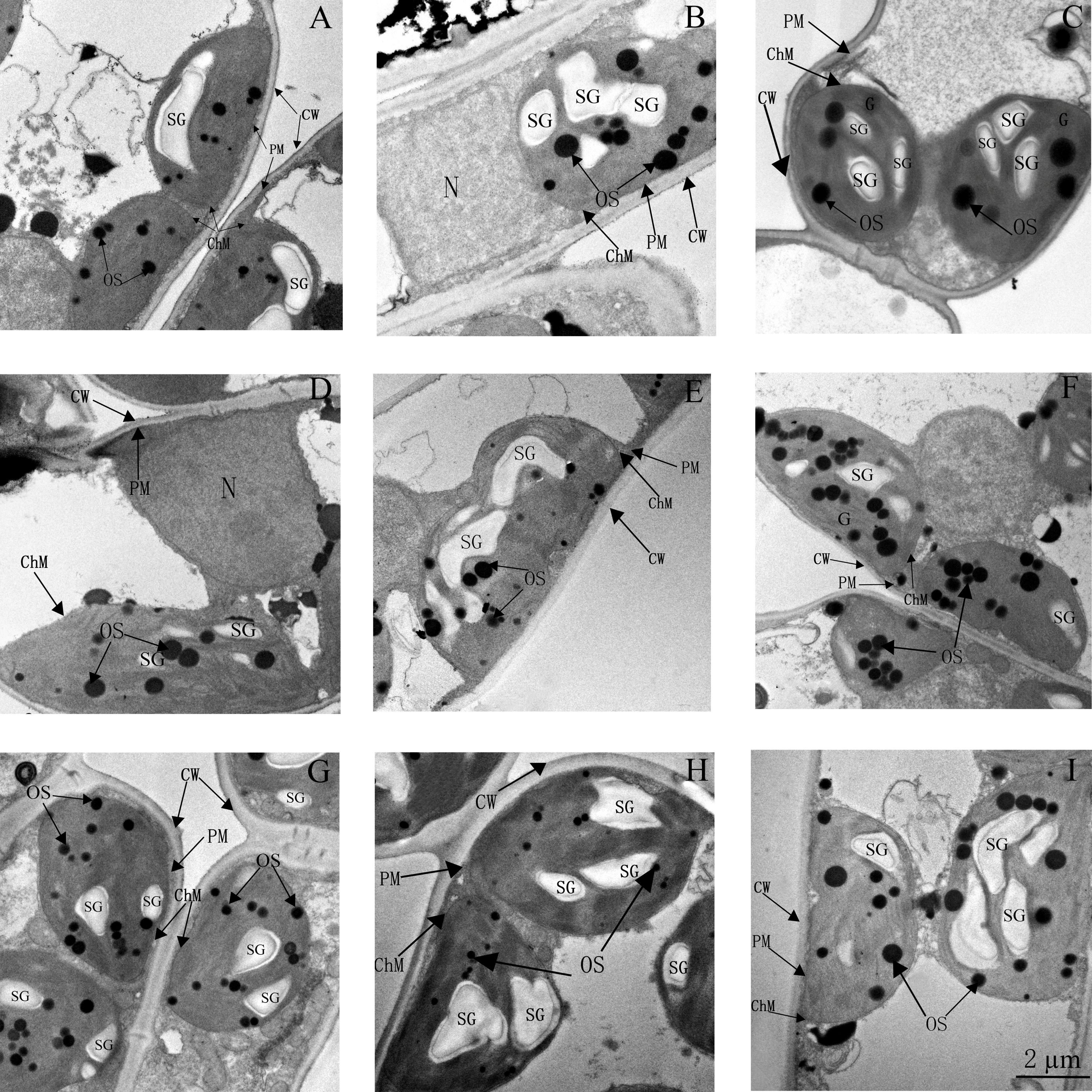
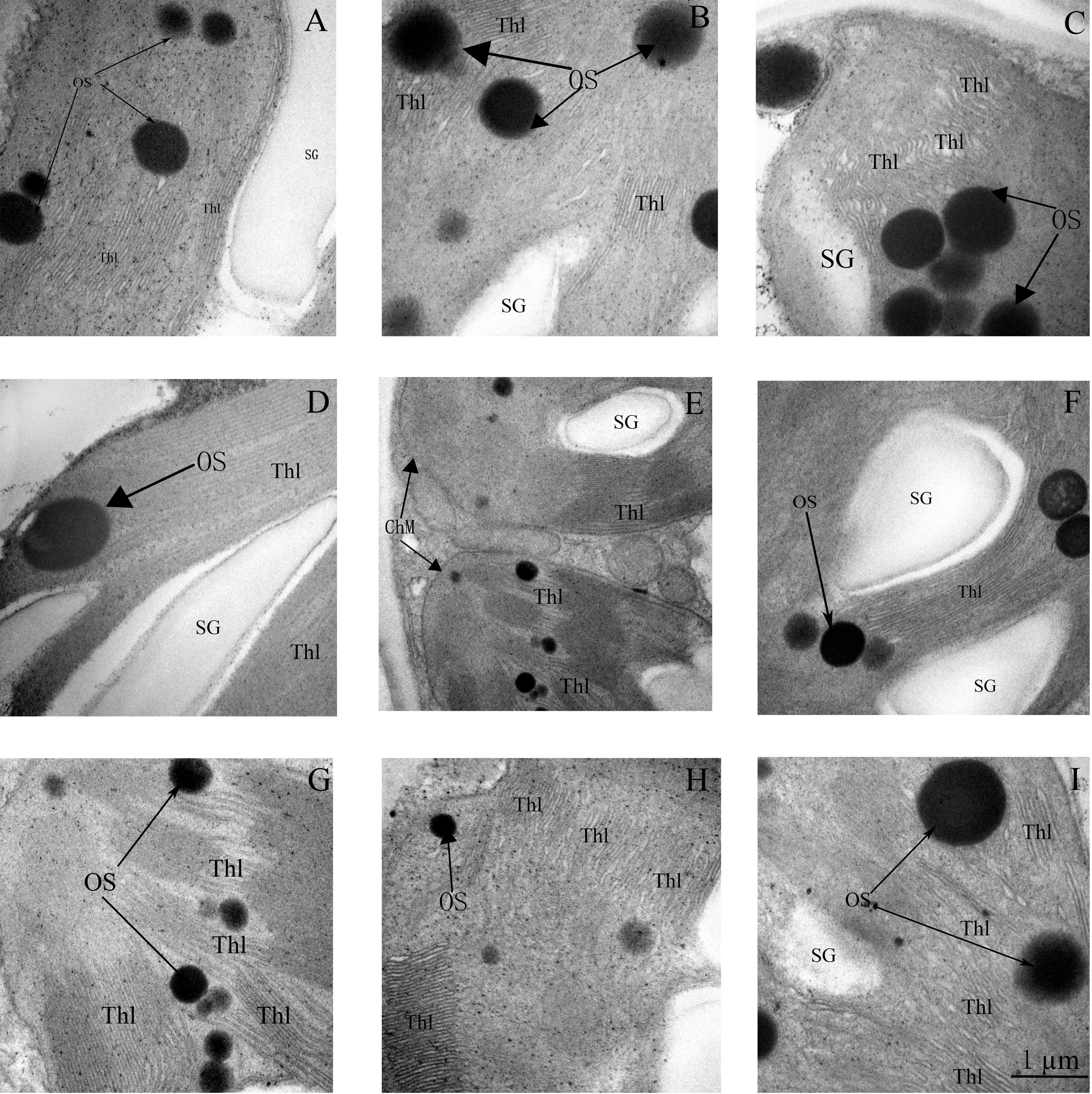
3.7. Ultrastructure of Chloroplasts and Thylakoids
4. Discussion
4.1. Photosynthesis
4.2. Antioxidative Effect in Leaves
4.3. Ion Toxicity and Chloroplast Ultrastructure
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shahid, S.A. Developments in spoil salinity assessment, modeling, mapping, and monitoring from regional to submicroscopic scales. In Developments in Soil Salinity Assessment and Reclamation; Shahid, S.A., Abelfattah, M.A., Taha, F.K., Eds.; Springer: Dordrecht, The Netherland, 2013; pp. 3–44. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Li, J.; Pu, L.; Han, M.; Zhu, M.; Zhang, R.; Xiang, Y. Soil salinization research in China: Advances and prospects. J. Geogra. Sci. 2014, 24, 943–960. [Google Scholar] [CrossRef]
- Chen, X.; Shen, Q.; Xu, Y. Hydraulic properties of typical salt-affected soils in Jiangsu Province, China. Front. Environ. Sci. Eng. China 2007, 1, 443–447. [Google Scholar] [CrossRef]
- DeGomez, T.; Wagner, M.R. Culture and use of black locust. HortTechnology 2001, 11, 279–288. [Google Scholar] [CrossRef]
- Liu, W.N.; Jiang, H.Y.; Yang, M.S. Selection of salt—tolerance on Robinia pseudoacacia seedings and impact of salt stress on seedlings’ physiological characteristics. J. Agric. Univ. Hebei 2010, 3, 62–66. [Google Scholar]
- Sitzia, T.; Campagnaro, T.; Dainese, M.; Cierjacks, A. Forest Ecology and Management Plant species diversity in alien black locust stands: A paired comparison with native stands across a north-Mediterranean range expansion. For. Ecol. Manag. 2012, 285, 85–91. [Google Scholar] [CrossRef]
- Tateno, R.; Tokuchi, N.; Yamanaka, N.; Du, S.; Otsuki, K.; Shimamura, T.; Xue, Z.; Wang, S.; Hou, Q. Comparison of litterfall production and leaf litter decomposition between an exotic black locust plantation and an indigenous oak forest near Yan’an on the Loess Plateau, China. For. Ecol. Manag. 2007, 241, 84–90. [Google Scholar] [CrossRef]
- Akatov, V.V.; Akatova, T.V.; Shadzhe, A.E. Robinia pseudoacacia L. in the Western Caucasus. Rus. J. Biol. Invasion 2016, 7, 105–118. [Google Scholar] [CrossRef]
- Holle, B.V.; Neill, C.; Largay, E.F.; Budreski, K.A.; Ozimec, B.; Clark, S.A.; Lee, K. Ecosystem legacy of the introduced N2-fixing tree Robinia pseudoacacia in a coastal forest. Oecologia 2013, 172, 915–924. [Google Scholar] [CrossRef]
- Mao, P.; Zhang, Y.; Cao, B.; Guo, L.; Shao, H.; Cao, Z.; Jiang, Q.; Wang, X. Effects of salt stress on eco-physiological characteristics in Robinia pseudoacacia based on salt-soil rhizosphere. Sci. Total Environ. 2016, 568, 118–123. [Google Scholar] [CrossRef]
- Zhao, G.; Li, S.; Sun, X.; Wang, Y.; Chang, Z. The role of silicon in physiology of the medicinal plant (Lonicera japonica L.) under salt stress. Sci. Rep. 2015, 5, 12696. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Muneer, S.; Ko, C.H.; Jeong, B.R. Silicon mitigates salinity stress by regulating the physiology, antioxidant enzyme activities, and protein expression in Capsicum annuum “Bugwang”. BioMed Res. Int. 2016, 2016. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, M.; Liu, L.; Meng, F. Physiological and proteomic responses of diploid and tetraploid black locust (Robinia pseudoacacia L.) subjected to salt stress. Int. J. Mol. Sci. 2013, 14, 20299–20325. [Google Scholar] [CrossRef]
- Xi, Z.; Wang, Z.; Fang, Y.; Hu, Z.; Hu, Y.; Deng, M.; Zhang, Z. Effects of 24-epibrassinolide on antioxidation defense and osmoregulation systems of young grapevines (V. vinifera L.) under chilling stress. Plant Growth Regul. 2013, 71, 57–65. [Google Scholar] [CrossRef]
- An, Y.; Zhou, H.; Zhong, M.; Sun, J.; Shu, S. Root proteomics reveals cucumber 24-epibrassinolide responses under Ca (NO3)2 stress. Plant Cell Rep. 2016, 35, 1081–1101. [Google Scholar] [CrossRef]
- Cui, L.; Zou, Z.; Zhang, J.; Zhao, Y.; Yan, F. 24-Epibrassinoslide enhances plant tolerance to stress from low temperatures and poor light intensities in tomato (Lycopersicon esculentum Mill.). Funct. Integr. Genomic. 2016, 16, 29–35. [Google Scholar] [CrossRef]
- Talaat, N.B.; Shawky, B.T. 24-Epibrassinolide ameliorates the saline stress and improves the productivity of wheat (Triticum aestivum L.). Environ. Exp. Bot. 2012, 82, 80–88. [Google Scholar] [CrossRef]
- Azhar, N.; Su, N.; Shabala, L.; Shabala, S. Exogenously applied 24-epibrassinolide (EBL) ameliorates detrimental effects of salinity by reducing K+ efflux via depolarization-activated K+ channels. Plant Cell Physiol. 2017, 58, 802–810. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circ. Calif. Agr. Ex. Stn. 1950, 347, 1–32. [Google Scholar]
- Yue, J.M.; You, Y.H.; Zhang, L.; Fu, Z.Y.; Wang, J.P.; Zhang, J.C.; Guy, R.D. Exogenous 24-epibrassinolide alleviates effects of salt stress on chloroplasts and photosynthesis in Robinia pseudoacacia L. seedlings. J. Plant Growth Reg. 2018, 1–14. [Google Scholar] [CrossRef]
- Sairam, R.K. Effects of homobrassinolide application on plant metabolism and grain yield under irrigated and moisture-stress conditions of two wheat varieties. Plant Growth Regul. 1994, 306, 173–181. [Google Scholar] [CrossRef]
- Hayat, S.; Ali, B.; Aiman Hasan, S.; Ahmad, A. Brassinosteroid enhanced the level of antioxidants under cadmium stress in Brassica juncea. Environ. Exp. Bot. 2007, 60, 33–41. [Google Scholar] [CrossRef]
- He, M.; Shi, D.; Wei, X.; Hu, Y.; Wang, T.; Xie, Y. Gender-related differences in adaptability to drought stress in the dioecious tree Ginkgo biloba. Acta Physiol. Plant. 2016, 38. [Google Scholar] [CrossRef]
- Fridovich, I. Superoxide Dismutases. Annu. Rev. Biochem. 1975, 44, 147–159. [Google Scholar] [CrossRef]
- Hammerschmidt, R.; Nuckles, E.M.; Kuć, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Cerović, Z.G.; Plesnicar, M. An improved procedure for the isolation of intact chloroplasts of high photosynthetic capacity. Biochem. J. 1984, 223, 543–545. [Google Scholar] [CrossRef] [Green Version]
- Song, X.S.; Tiao, C.L.; Shi, K.; Mao, W.H.; Ogweno, J.O.; Zhou, Y.H.; Yu, J.Q. The response of antioxidant enzymes in cellular organelles in cucumber (Cucumis sativus L.) leaves to methyl viologen-induced photo-oxidative stress. Plant Growth Regul. 2006, 49, 85–93. [Google Scholar] [CrossRef]
- Abo-Ogiala, A.; Carsjens, C.; Diekmann, H.; Fayyaz, P.; Herrfurth, C.; Feussner, I.; Polle, A. Temperature-induced lipocalin (TIL) is translocated under salt stress and protects chloroplasts from ion toxicity. J. Plant Physiol. 2014, 171, 250–259. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Zhu, X.H.; Ding, H.D.; Yang, S.J.; Chen, Y.Y. Foliar application of 24-epibrassinolide alleviates high-temperature-induced inhibition of photosynthesis in seedlings of two melon cultivars. Photosynthetica 2013, 51, 341–349. [Google Scholar] [CrossRef]
- Ramakrishna, B.; Rao, S.S.R. 24-Epibrassinolide alleviated zinc-induced oxidative stress in radish (Raphanus sativus L.) seedlings by enhancing antioxidative system. Plant Growth Regul. 2012, 68, 249–259. [Google Scholar] [CrossRef]
- Ali, Q.; Athar, H.U.R.; Ashraf, M. Modulation of growth, photosynthetic capacity and water relations in salt stressed wheat plants by exogenously applied 24-epibrassinolide. Plant Growth Regul. 2008, 56, 107–116. [Google Scholar] [CrossRef]
- Xu, G.; Huang, T.F.; Zhang, X.L.; Duan, B.L. Significance of mesophyll conductance for photosynthetic capacity and water-use efficiency in response to alkaline stress in Populus cathayana seedlings. Photosynthetica 2013, 51, 438–444. [Google Scholar] [CrossRef] [Green Version]
- Kalaji, H.M.; Govindjee; Bosa, K.; Koscielniak, J.; Zuk-Golaszewska, K. Effects of salt stress on photosystem II efficiency and CO2 assimilation of two Syrian barley landraces. Environ. Exp. Bot. 2011, 73, 64–72. [Google Scholar] [CrossRef]
- Oxborough, K. Imaging of chlorophyll a fluorescence: Theoretical and practical aspects of an emerging technique for the monitoring of photosynthetic performance. J. Exp. Bot. 2004, 55, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Shao, M.; He, Y.; Guan, R.; Chu, P.; Jiang, H. Proteome dynamics and physiological responses to short-term salt stress in brassica napus leaves. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Wang, Z.; Zheng, P.; Meng, J.; Xi, Z. Effect of exogenous 24-epibrassinolide on chlorophyll fluorescence, leaf surface morphology and cellular ultrastructure of grape seedlings (Vitis vinifera L.) under water stress. Acta Physiol. Plant. 2015, 37, 1–12. [Google Scholar] [CrossRef]
- Thussagunpanit, J.; Jutamanee, K.; Kaveeta, L.; Chai-arree, W.; Pankean, P.; Homvisasevongsa, S.; Suksamrarn, A. Comparative effects of brassinosteroid and brassinosteroid mimic on improving photosynthesis, lipid peroxidation, and rice seed set under heat stress. J. Plant Growth Regul. 2015, 34, 320–331. [Google Scholar] [CrossRef]
- Plesa, I.; González-Orenga, S.; Al Hassan, M.; Sestras, A.; Vicente, O.; Prohens, J.; Sestras, R.; Boscaiu, M. Effects of Drought and Salinity on European Larch (Larix decidua Mill.) Seedlings. Forests 2018, 9, 320. [Google Scholar] [CrossRef]
- Xia, X.J.; Fang, P.P.; Guo, X.; Qian, X.J.; Zhou, J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Brassinosteroid-mediated apoplastic H2O2-glutaredoxin 12/14 cascade regulates antioxidant capacity in response to chilling in tomato. Plant Cell Environ. 2018, 41, 1052–1064. [Google Scholar] [CrossRef]
- Yadava, P.; Kaushal, J.; Gautam, A.; Parmar, H.; Singh, I. Physiological and biochemical effects of 24-epibrassinolide on heat-stress adaptation in Maize (Zea mays L.). Nat. Sci. 2016, 8, 171–179. [Google Scholar] [CrossRef]
- Tůmová, L.; Tarkowská, D.; Řřová, K.; Marková, H.; Kočová, M.; Rothová, O.; čečetka, P.; Holá, D. Drought-tolerant and drought-sensitive genotypes of maize (Zea mays L.) differ in contents of endogenous brassinosteroids and their drought-induced changes. PLoS ONE 2018, 13, 1–22. [Google Scholar] [CrossRef] [PubMed]
- James, R.A.; Munns, R.; Von Caemmerer, S.; Trejo, C.; Miller, C.; Condon, T. Photosynthetic capacity is related to the cellular and subcellular partitioning of Na+, K+ and Cl− in salt-affected barley and durum wheat. Plant Cell Environ. 2006, 29, 2185–2197. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chen, S.; Deng, L.; Fritz, E.; Hüttermann, A.; Polle, A. Leaf photosynthesis, fluorescence response to salinity and the relevance to chloroplast salt compartmentation and anti-oxidative stress in two poplars. Trees Struct. Funct. 2007, 21, 581–591. [Google Scholar] [CrossRef]
- Shu, S.; Yuan, Y.; Chen, J.; Sun, J.; Zhang, W.; Tang, Y.; Zhong, M.; Guo, S. The role of putrescine in the regulation of proteins and fatty acids of thylakoid membranes under salt stress. Sci. Rep. 2015, 5, 1–16. [Google Scholar] [CrossRef]
- Cao, B.L.; Ma, Q.; Zhao, Q.; Wang, L.; Xu, K. Effects of silicon on absorbed light allocation, antioxidant enzymes and ultrastructure of chloroplasts in tomato leaves under simulated drought stress. Sci. Hortic. 2015, 194, 53–62. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, Q.; Ervin, E.H.; Yang, Z.; Zhang, X. Physiological mechanism of enhancing salt stress tolerance of Perennial ryegrass by 24-epibrassinolide. Fron. Plant Sci. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Kapoor, D.; Rattan, A.; Gautam, V.; Bhardwaj, R. Mercury-induced changes in growth, metal & ions uptake, photosynthetic pigments, osmoprotectants and antioxidant defence system in Raphanus sativus L. seedlings and role of steroid hormone in stress amelioration. J. Pharmacogn. Phytochem. 2016, 5, 259–265. [Google Scholar]
- Fisher, A.B. Redox signaling across cell membranes. Antioxid. Redox Signal. 2009, 11, 1349–1356. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Wali, M.; Gunsè, B.; Llugany, M.; Corrales, I.; Abdelly, C.; Poschenrieder, C.; Ghnaya, T. High salinity helps the halophyte Sesuvium portulacastrum in defense against Cd toxicity by maintaining redox balance and photosynthesis. Planta 2016, 244, 333–346. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Seeds Pretreatment | Seedling Pretreatment | Salt Stress | |
|---|---|---|---|---|
| 24-epiBL(µM) by Seeds Soaking | 24-epiBL(µM) by Foliar Spraying | 24-epiBL(µM) by Roots Dipping | NaCl (mM) | |
| CK | — | — | — | |
| SS-S1 | — | — | 100 | |
| SS-S2 | — | — | 200 | |
| Sew-0.21S1 | 0.21 | — | 100 | |
| Sew-0.62S1 | 0.62 | — | 100 | |
| Sew-1.04S1 | 1.04 | — | 100 | |
| Spw-0.21S1 | 0.21 | 100 | ||
| Spw-0.62S1 | 0.62 | 100 | ||
| Spw-1.04S1 | 1.04 | 100 | ||
| Diw-0.21S1 | 0.21 | 100 | ||
| Diw-0.62S1 | 0.62 | 100 | ||
| Diw-1.04S1 | 1.04 | 100 | ||
| Sew-0.21S2 | 0.21 | — | 200 | |
| Sew-0.62S2 | 0.62 | — | 200 | |
| Sew-1.04S2 | 1.04 | — | 200 | |
| Spw-0.21S2 | 0.21 | 200 | ||
| Spw-0.62S2 | 0.62 | 200 | ||
| Spw-1.04S2 | 1.04 | 200 | ||
| Diw-0.21S2 | 0.21 | 200 | ||
| Diw-0.62S2 | 0.62 | 200 | ||
| Diw-1.04S2 | 1.04 | 200 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, J.; Fu, Z.; Zhang, L.; Zhang, Z.; Zhang, J. The Positive Effect of Different 24-epiBL Pretreatments on Salinity Tolerance in Robinia pseudoacacia L. Seedlings. Forests 2019, 10, 4. https://doi.org/10.3390/f10010004
Yue J, Fu Z, Zhang L, Zhang Z, Zhang J. The Positive Effect of Different 24-epiBL Pretreatments on Salinity Tolerance in Robinia pseudoacacia L. Seedlings. Forests. 2019; 10(1):4. https://doi.org/10.3390/f10010004
Chicago/Turabian StyleYue, Jianmin, Zhiyuan Fu, Liang Zhang, Zihan Zhang, and Jinchi Zhang. 2019. "The Positive Effect of Different 24-epiBL Pretreatments on Salinity Tolerance in Robinia pseudoacacia L. Seedlings" Forests 10, no. 1: 4. https://doi.org/10.3390/f10010004
APA StyleYue, J., Fu, Z., Zhang, L., Zhang, Z., & Zhang, J. (2019). The Positive Effect of Different 24-epiBL Pretreatments on Salinity Tolerance in Robinia pseudoacacia L. Seedlings. Forests, 10(1), 4. https://doi.org/10.3390/f10010004