Nitrogen Nutrition of European Beech Is Maintained at Sufficient Water Supply in Mixed Beech-Fir Stands

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sites and Experimental Approaches
2.2. Soil Sampling and Analyses
2.3. Sampling of Plant Material
2.4. Element and Stable Isotope Analyses of C and N
2.5. Protein and Amino Acid Analyses
2.6. Calculation of Structural N
2.7. N Uptake Experiments
2.8. Statistical Analysis
3. Results
3.1. Inorganic Soil Nitrogen Contents in Fall
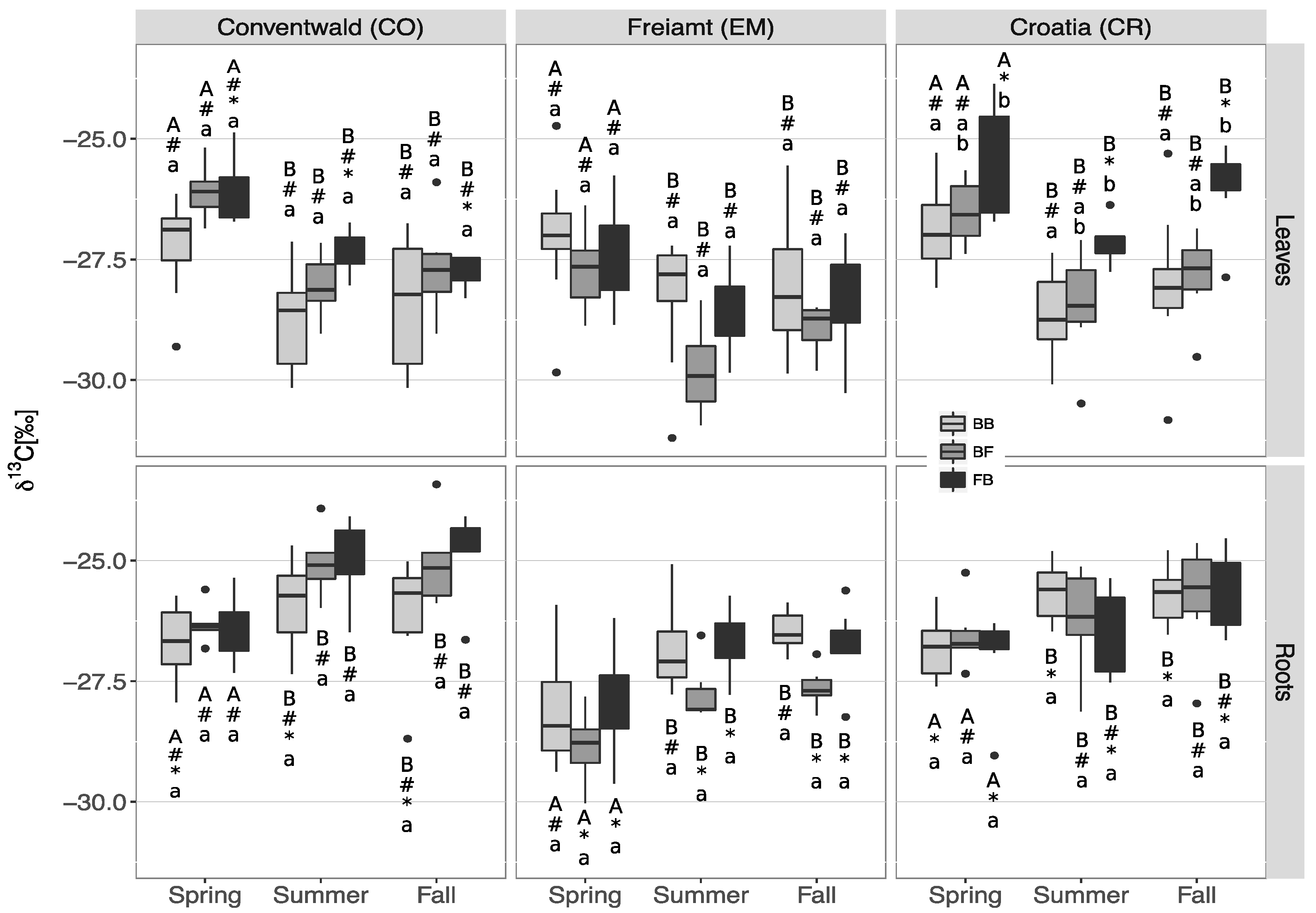
3.2. δ13C Signatures in Leaves/Needles and Roots
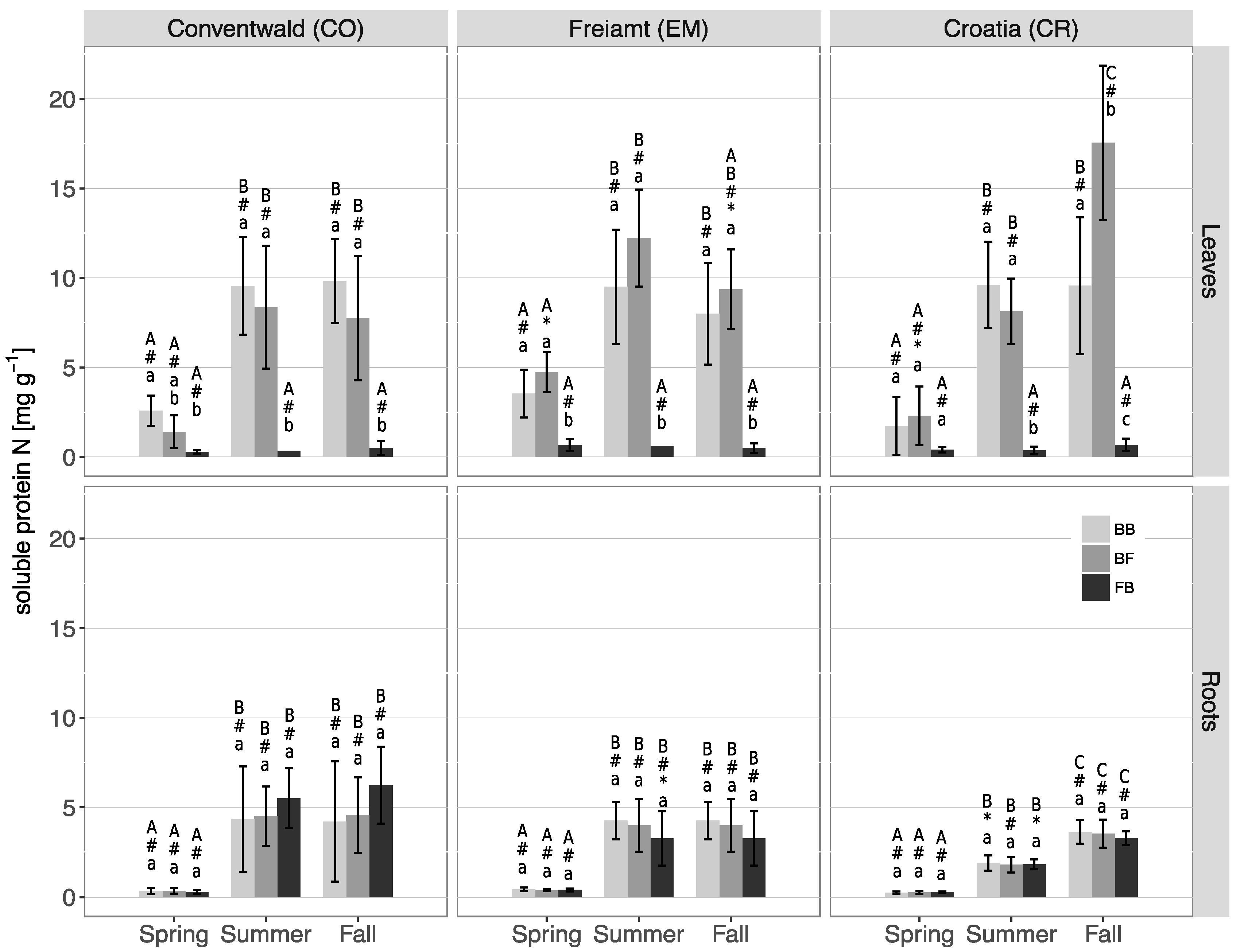
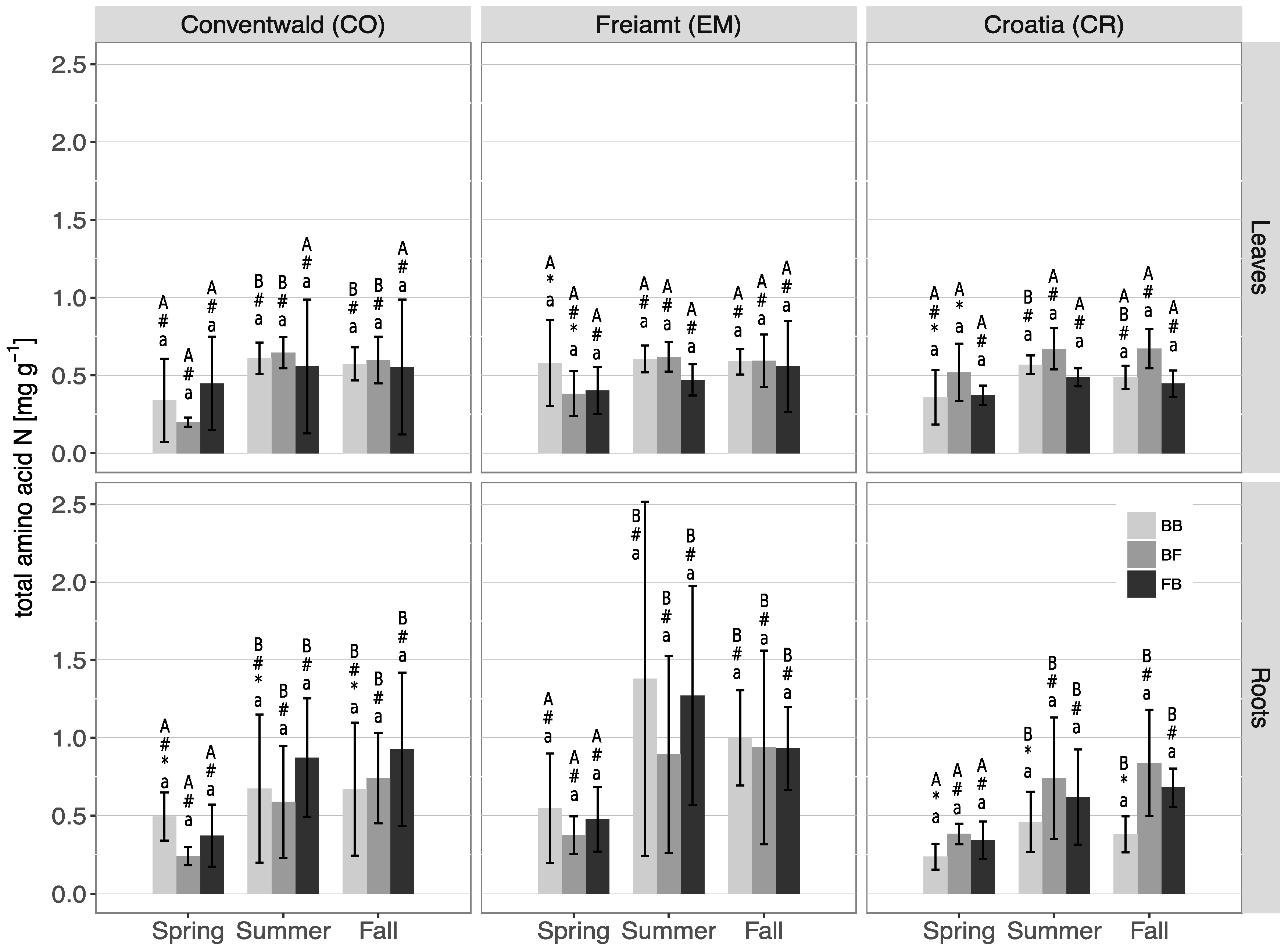
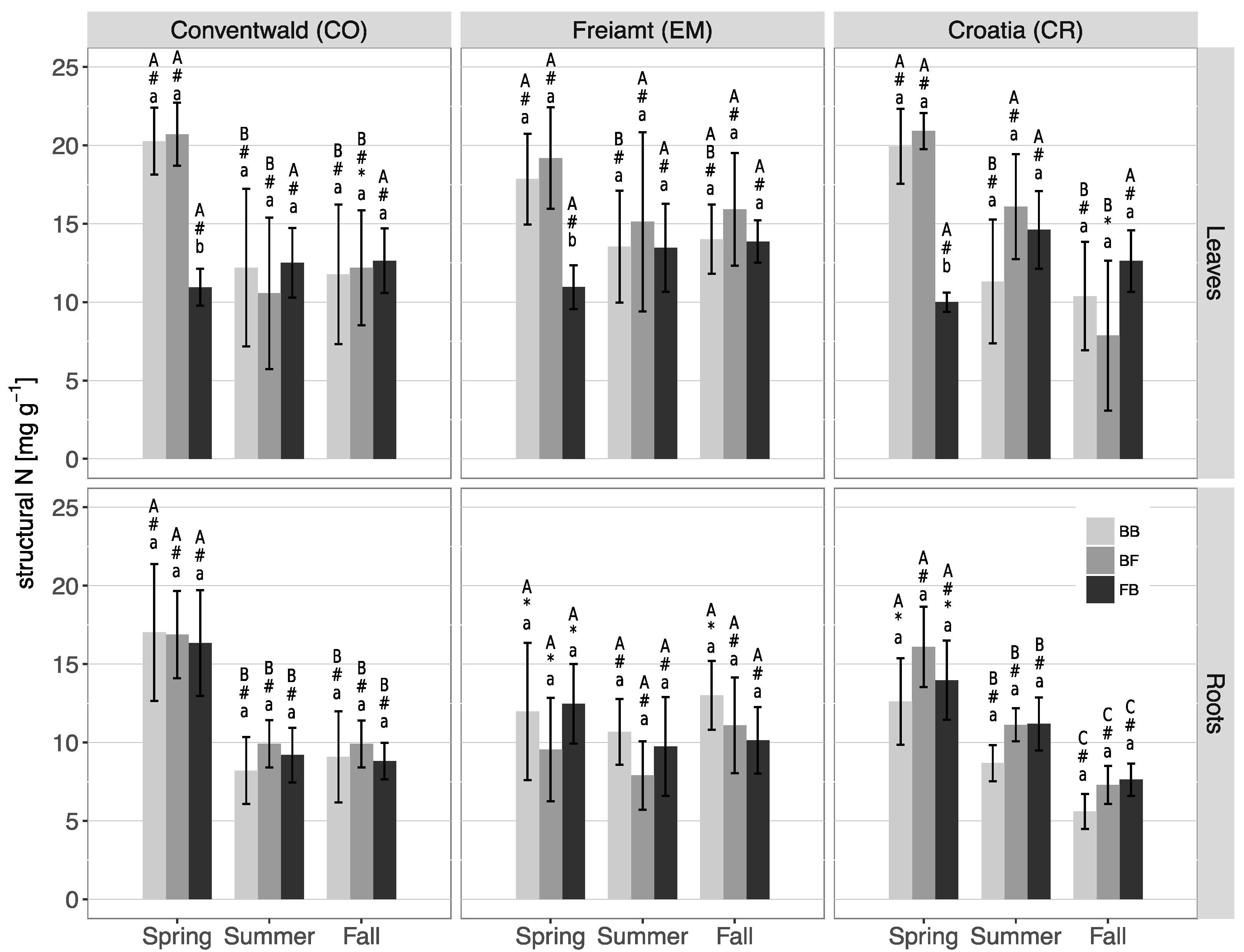
3.3. N Contents and N Partitioning in Leaves/Needles and Roots
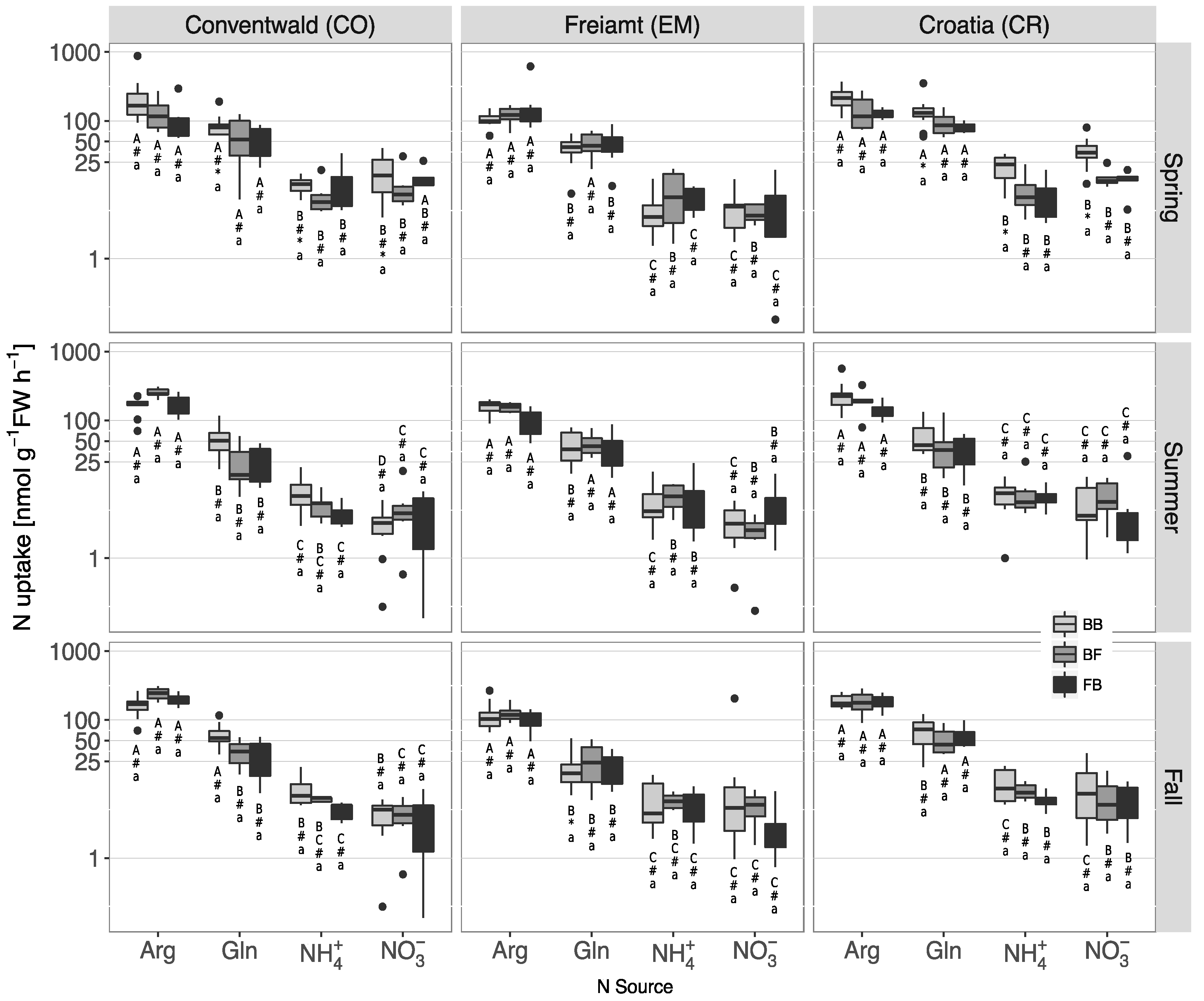
3.4. N Uptake Capacities of the Roots
3.5. Correlations between Environmental-, Soil-, and Plant Parameters
3.5.1. Environmental and Plant Parameters Relations
3.5.2. Soil and Plant Parameter Relations in Fall
3.5.3. Relations of N Uptake and Soil Parameters in Fall
4. Discussion
4.1. Beech and Silver-Fir Performances are Similar at the Croatian and Black Forest Field Sites
4.2. The Performance of Beech Tends to Be Influenced by the Presence of Silver-Fir
4.3. What Is the Driving Force of the N Nutrition of Beech?
4.4. Silver-Fir Total N Contents Indicate the Roots as Storage Location
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Taberlet, P.; Fumagalli, L.; Wust-Saucy, A.G.; Cosson, J.F. Comparative Phylogeography and Postglacial Colonization Routes in Europe. Mol. Ecol. 1998, 7, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Konnert, M.; Bergmann, F. The Geographical Distribution of Genetic Variation of Silver Fir (Abies alba, Pinaceae) in Relation to Its Migration History. Plant Syst. Evol. 1995, 196, 19–30. [Google Scholar] [CrossRef]
- Magri, D.; Vendramin, G.G.; Comps, B.; Dupanloup, I.; Geburek, T.; Gömöry, D.; Latałowa, M.; Litt, T.; Paule, L.; Roure, J.M.; et al. A New Scenario for the Quaternary History of European Beech Populations: Palaeobotanical Evidence and Genetic Consequences. New Phytol. 2006, 171, 199–221. [Google Scholar] [CrossRef] [PubMed]
- Demesure, B.; Comps, B.; Petit, R.J. Chloroplast DNA Phylogeography of the Common Beech (Fagus sylvatica L.) in Europe. Evolution 1996, 50, 2515–2520. [Google Scholar] [CrossRef] [PubMed]
- Ellenberg, H. Vegetation Ecology of Central Europe; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Fang, J.; Lechowicz, M.J. Climatic Limits for the Present Distribution of Beech (Fagus L.) Species in the World. J. Biogeogr. 2006, 33, 1804–1819. [Google Scholar] [CrossRef]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas Mit Den Alpen: In Ökologischer, Dynamischer Und Historischer Sicht; UTB Ulmer: Stuttgart, Germany, 2010; p. 1333. [Google Scholar]
- Frey, W.; Lösch, R. Geobotanik; Spektrum Akademischer Verlag: Heidelberg, Germany, 2010; p. 600. [Google Scholar]
- Ellenberg, H. Vegetation Ecology of Central Europe, 4th ed.; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Tinner, W.; Colombaroli, D.; Heiri, O.; Henne, P.D.; Steinacher, M.; Untenecker, J.; Vescovi, E.; Allen, J.R.M.; Carraro, G.; Conedera, M.; et al. The Past Ecology of Abies alba Provides New Perspectives on Future Responses of Silver Fir Forests to Global Warming. Ecol. Monogr. 2013, 83, 419–439. [Google Scholar] [CrossRef]
- Oberdorfer, E. Süddeutsche Pflanzengesellschaften; Gustav Fischer Verlag: Jena, Germany, 1957; p. 564. [Google Scholar]
- Blanco, E.; Casado, M.A.; Costa, M.; Escribano, R.; García, M.; Génova, M.; Gómez, A.; Gómez, F.; Moreno, J.C.; Morla, C. Los Bosques Ibéricos. Una Interpretación Geobotánica, 4th ed.; Planeta: Barcelona, Spain, 1997; p. 572. [Google Scholar]
- Schraml, U.; Volz, K. Conversion of Coniferous Forests in Social and Political Perspectives. Findings from Selected Countries with Special Respect to Germany. Norway Spruce Convers.—Options Conseq. EFI Res. Rep. 2004, 18, 97–119. [Google Scholar]
- Guckland, A.; Jacob, M.; Flessa, H.; Thomas, F.M.; Leuschne, C. Acidity, Nutrient Stocks, and Organic-Matter Content in Soils of a Temperate Deciduous Forest with Different Abundance of European Beech (Fagus sylvatica L.). J. Plant Nutr. Soil Sci. 2009, 172, 500–511. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. The Stress Concept in Plants: An Introduction. Ann. N. Y. Acad. Sci. 1998, 851, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Kunstler, G.; Curt, T.; Bouchaud, M.; Lepart, J. Growth, Mortality, and Morphological Response of European Beech and Downy Oak along a Light Gradient in Sub-Mediterranean Forest. Can. J. For. Res. 2005, 35, 1657–1668. [Google Scholar] [CrossRef]
- Curt, T.; Coll, L.; Prévosto, B.; Balandier, P.; Kunstler, G.; Truţǎ, E.; Cǎpraru, G.; Surdu, Ş.; Zamfirache, M.M.; Olteanu, Z.; et al. Plasticity in Growth, Biomass Allocation and Root Morphology in Beech Seedlings as Induced by Irradiance and Herbaceous Competition. Ann. For. Sci. 2005, 59, 51–60. [Google Scholar] [CrossRef]
- Bonosi, L. The Influence of Light and Size on Photosynthetic Performance, Light, Interception, Biomass Partitioning and Tree Architecture in Open Grown Acer pseudoplatanus, Fraxinus excelsior and Fagus sylvatica Seedlings; Albert-Ludwigs-Univ., Waldbau-Inst.: Freiburg, Germany, 2006. [Google Scholar]
- Petritan, A.M.; von Lüpke, B.; Petritan, I.C. Influence of Light Availability on Growth, Leaf Morphology and Plant Architecture of Beech (Fagus sylvatica L.), Maple (Acer pseudoplatanus L.) and Ash (Fraxinus excelsior L.) Saplings. Eur. J. For. Res. 2009, 128, 61–74. [Google Scholar] [CrossRef]
- Johnson, J.D.; Tognetti, R.; Michelozzi, M.; Pinzauti, S.; Minotta, G.; Borghetti, M. Ecophysiological Responses of Fagus sylvatica Seedlings to Changing Light Conditions—The Interaction of Light Environment and Soil Fertility on Seedling Physiology. Physiol. Plant 1997, 101, 124–134. [Google Scholar] [CrossRef]
- Simon, J.; Dannenmann, M.; Gasche, R.; Holst, J.; Mayer, H.; Papen, H.; Rennenberg, H. Competition for Nitrogen between Adult European Beech and Its Offspring Is Reduced by Avoidance Strategy. For. Ecol. Manag. 2011, 262, 105–114. [Google Scholar] [CrossRef]
- Petritan, A.M.; Von Lüpke, B.; Petritan, I.C. Effects of Shade on Growth and Mortality of Maple (Acer pseudoplatanus), Ash (Fraxinus excelsior) and Beech (Fagus sylvatica) Saplings. Forestry 2007, 80, 397–412. [Google Scholar] [CrossRef]
- Annighöfer, P. Stress Relief through Gap Creation? Growth Response of a Shade Tolerant Species (Fagus sylvatica L.) to a Changed Light Environment. For. Ecol. Manag. 2018, 415, 139–147. [Google Scholar]
- Fotelli, M.N.; Rennenberg, H.; Holst, T.; Mayer, H.; Geßler, A. Carbon Isotope Composition of Various Tissues of Beech (Fagus sylvatica) Regeneration Is Indicative of Recent Environmental Conditions within the Forest Understorey. New Phytol. 2003, 159, 229–244. [Google Scholar] [CrossRef]
- Peuke, A.D.; Gessler, A.; Rennenberg, H. The Effect of Drought on C and N Stable Isotopes in Different Fractions of Leaves, Stems and Roots of Sensitive and Tolerant Beech Ecotypes. Plant Cell Environ. 2006, 29, 823–835. [Google Scholar] [CrossRef]
- Röhrig, E.; Bartsch, N.; von Lüpke, B.; Dengler, A. Waldbau Auf Ökologischer Grundlage; Ulmer: Stuttgart, Germany, 2006; p. 479. [Google Scholar]
- Schlesinger, W.H.; Dietze, M.C.; Jackson, R.B.; Phillips, R.P.; Rhoades, C.C.; Rustad, L.E.; Vose, J.M. Forest Biogeochemistry in Response to Drought. Glob. Chang. Biol. 2016, 22, 2318–2328. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H. (Ted); et al. A Global Overview of Drought and Heat-Induced Tree Mortality Reveals Emerging Climate Change Risks for Forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Vitali, V.; Forrester, D.I.; Bauhus, J. Know Your Neighbours: Drought Response of Norway Spruce, Silver Fir and Douglas Fir in Mixed Forests Depends on Species Identity and Diversity of Tree Neighbourhoods. Ecosystems 2018, 21, 1215–1229. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Gessler, A. Global Climate Change and Tree Nutrition: Influence of Water Availability. Tree Physiol. 2010, 30, 1221–1234. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global Convergence in the Vulnerability of Forests to Drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Fuhrer, J.; Beniston, M.; Fischlin, A.; Frei, C.; Goyette, S.; Jasper, K.; Pfister, C. Climate Risks and Their Impact on Agriculture and Forests in Switzerland. Clim. Chang. 2006, 79, 79–102. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of Plant Survival and Mortality during Drought: Why Do Some Plants Survive While Others Succumb to Drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J.; et al. Regional Vegetation Die-off in Response to Global-Change-Type Drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef] [PubMed]
- Bigler, C.; Bräker, O.U.; Bugmann, H.; Dobbertin, M.; Rigling, A. Drought as an Inciting Mortality Factor in Scots Pine Stands of the Valais, Switzerland. Ecosystems 2006, 9, 330–343. [Google Scholar] [CrossRef]
- Rennenberg, H.; Seiler, W.; Matyssek, R.; Gessler, A.; Kreuzwieser, J. European Beech (Fagus sylvatica L.)—A Forest Tree without Future in the South of Central Europe. Allg. Forst Und Jagdzeitung 2004, 175, 210–224. [Google Scholar]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; p. 1535. [Google Scholar]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate Change Impacts, Adaptive Capacity, and Vulnerability of European Forest Ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- BMEL. Der Wald in Deutschland: Ausgewählte Ergebnisse Der Dritten Bundeswaldinventur; Bundesministerium Für Ernährung Und Landwirtschaft: Berlin, Germany, 2014. [Google Scholar]
- Puettmann, K.J.; Coates, K.D.; Messier, C. A Critique of Silviculture. Managing for Complexity; Island Press: Washington DC, USA, 2008; p. 189. [Google Scholar]
- Fritz, P.; Jenssen, M. Ökologischer Waldumbau in Deutschland–Fragen Antworten, Perspektiven; Oekom: München, Germany, 2006; p. 351. [Google Scholar]
- Robakowski, P.; Wyka, T.; Samardakiewicz, S.; Kierzkowski, D. Growth, Photosynthesis, and Needle Structure of Silver Fir (Abies alba Mill.) Seedlings under Different Canopies. For. Ecol. Manag. 2004, 201, 211–227. [Google Scholar] [CrossRef]
- Lebourgeois, F.; Gomez, N.; Pinto, P.; Mérian, P. Mixed Stands Reduce Abies alba Tree-Ring Sensitivity to Summer Drought in the Vosges Mountains, Western Europe. For. Ecol. Manag. 2013, 303, 61–71. [Google Scholar] [CrossRef]
- PALUCH, J.G.; GRUBA, P. Effect of Local Species Composition on Topsoil Properties in Mixed Stands with Silver Fir (Abies alba Mill.). Forestry 2012, 85, 413–426. [Google Scholar] [CrossRef]
- Koch, A.S.; Matzner, E. Heterogeneity of Soil and Soil Solution Chemistry under Norway Spruce (Picea abies Karst.) and European Beech (Fagus sylvatica L.) as Influenced by Distance from the Stem Basis. Plant Soil 1993, 151, 227–237. [Google Scholar] [CrossRef]
- Magh, R.-K.; Grün, M.; Knothe, V.E.; Stubenazy, T.; Tejedor, J.; Dannenmann, M.; Rennenberg, H. Silver-Fir (Abies alba MILL.) Neighbors Improve Water Relations of European Beech (Fagus sylvatica L.), but Do Not Affect, N. Nutrition. Trees 2018, 32, 337–348. [Google Scholar] [CrossRef]
- Caldwell, M.M.; Dawson, T.E.; Richards, J.H. Hydraulic Lift: Consequences of Water Efflux from the Roots of Plants. Oecologia 1998, 113, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.; Li, X.; Rennenberg, H.; Näsholm, T. Competition for Nitrogen between European Beech and Sycamore Maple Shifts in Favour of Beech with Decreasing Light Availability. Tree Physiol. 2014, 34, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Klinka, K.; Mathey, A.-H.; Wang, X.; Varga, P.; Chourmouzis, C. Are Mixed-Species Stands More Productive than Single-Species Stands: An Empirical Test of Three Forest Types in British Columbia and Alberta. Can. J. For. Res. 2003, 33, 1227–1237. [Google Scholar] [CrossRef]
- Casper, B.B.; Jackson, R.B. Plant Competition Underground. Annu. Rev. Ecol. Syst. 1997, 28, 545–570. [Google Scholar] [CrossRef]
- Rothe, A.; Binkley, D. Nutritional Interactions in Mixed Species Forests: A. Synthesis. Can. J. For. Res. 2001, 31, 1855–1870. [Google Scholar] [CrossRef]
- Dannenmann, M.; Bimüller, C.; Gschwendtner, S.; Leberecht, M.; Tejedor, J.; Bilela, S.; Gasche, R.; Hanewinkel, M.; Baltensweiler, A.; Kögel-Knabner, I.; et al. Climate Change Impairs Nitrogen Cycling in European Beech Forests. PLoS ONE 2016, 11, e0158823. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, G. Die Untersuchung von Böden—Methodenbuch Band 1, 4th ed.; VDLUFA-Verlag: Darmstadt, Germany, 1991. [Google Scholar]
- Arbeitskreis Standortskartierung. Forstliche Standortsaufnahme: Begriffe, Definitionen, Einteilungen, Kennzeichnungen, Erläuterungen, 7th ed.; Arbeitskreis Standortskartierung in der Arbeitsgemeinschaft Forsteinrichtung: Eching, München, 2016. [Google Scholar]
- Contin, D.R.; Soriani, H.H.; Hernández, I.; Furriel, R.P.M.; Munné-Bosch, S.; Martinez, C.A. Antioxidant and Photoprotective Defenses in Response to Gradual Water Stress under Low and High Irradiance in Two Malvaceae Tree Species Used for Tropical Forest Restoration. Trees 2014, 28, 1705–1722. [Google Scholar] [CrossRef]
- Simon, J.; Waldhecker, P.; Brüggemann, N.; Rennenberg, H. Competition for Nitrogen Sources between European Beech (Fagus sylvatica) and Sycamore Maple (Acer pseudoplatanus) Seedlings. Plant Biol. 2010, 12, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Jansen, K.; Junker, L.V.; Eiblmeier, M.; Kreuzwieser, J.; Gessler, A.; Ensminger, I.; Rennenberg, H. Elevated Temperature Differently Affects Foliar Nitrogen Partitioning in Seedlings of Diverse Douglas Fir Provenances. Tree Physiol. 2014, 34, 1090–1101. [Google Scholar] [CrossRef] [PubMed]
- Winter, H.; Lohaus, G.; Heldt, H.W. Phloem Transport of Amino Acids in Relation to Their Cytosolic Levels in Barley Leaves. Plant Physiol. 1992, 99, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Simon, J.; Kuster, T.M.; Arend, M.; Siegwolf, R.; Rennenberg, H. Nitrogen Partitioning in Oak Leaves Depends on Species, Provenance, Climate Conditions and Soil Type. Plant Biol. 2013, 15, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Dannenmann, M.; Simon, J.; Gasche, R.; Holst, J.; Naumann, P.S.; Kögel-Knabner, I.; Knicker, H.; Mayer, H.; Schloter, M.; Pena, R.; et al. Tree Girdling Provides Insight on the Role of Labile Carbon in Nitrogen Partitioning between Soil Microorganisms and Adult European Beech. Soil Biol. Biochem. 2009, 41, 1622–1631. [Google Scholar] [CrossRef]
- Gessler, A.; Schneider, S.; Sengbusch v., D.; Weber, P.; Hanemann, U.; Huber, C.; Rothe, A.; Kreutzer, K.; Rennenberg, H. Field and Laboratory Experiments on Net Uptake of Nitrate and Ammonium by the Roots of Spruce (Picea abies) and Beech (Fagus sylvatica) Trees. New Phytol. 1998, 138, 275–285. [Google Scholar] [CrossRef]
- Geßler, A.; Kreuzwieser, J.; Dopatka, T.; Rennenberg, H. Diurnal Courses of Ammonium Net Uptake by the Roots of Adult Beech (Fagus sylvatica) and Spruce (Picea abies) Trees. Plant Soil 2002, 240, 23–32. [Google Scholar] [CrossRef]
- Stoelken, G.; Simon, J.; Ehlting, B.; Rennenberg, H. The Presence of Amino Acids Affects Inorganic N Uptake in Non-Mycorrhizal Seedlings of European Beech (Fagus sylvatica L.). Tree Physiol. 2010, 30, 1118–1128. [Google Scholar] [CrossRef] [PubMed]
- Kreuzwieser, J.; Fürniss, S.; Rennenberg, H. Impact of Waterlogging on the N-Metabolism of Flood Tolerant and Non-Tolerant Tree Species. Plant Cell Environ. 2002, 25, 1039–1049. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Dorman, C. Parametrische Statistik. Verteilungen, Maximum Likelihood Und GLM in R; Springer: Heidelberg, Germany, 2013; Volume 53. [Google Scholar]
- Crawley, M.J. Statistik Mit R; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2012; p. 424. [Google Scholar]
- Mellert, K.H.; Göttlein, A. Comparison of New Foliar Nutrient Thresholds Derived from van Den Burg’s Literature Compilation with Established Central European References. Eur. J. For. Res. 2012, 131, 1461–1472. [Google Scholar] [CrossRef]
- Göttlein, A.; Baier, R.; Mellert, K.H. Neue Ernährungskennwerte Für Die Forstlichen Hauptbaumarten in Mitteleuropa—Eine Statistische Herleitung Aus VAN DEN BURG’S Literaturzusammenstellung. Allg. Forst-und Jagdzeitung 2011, 182, 173–186. [Google Scholar]
- Bergman, W. Ernährungsstörungen Bei Kulturpflanzen, Entstehung Und Diagnose; Gustav Fischer Verlag: Jena, Germany, 1983; p. 614. [Google Scholar]
- Peguero-Pina, J.J.; Camarero, J.J.; Abadía, A.; Martín, E.; González-Cascón, R.; Morales, F.; Gil-Pelegrín, E. Physiological Performance of Silver-Fir (Abies alba Mill.) Populations under Contrasting Climates near the South-Western Distribution Limit of the Species. Flora 2007, 202, 226–236. [Google Scholar] [CrossRef]
- Millard, P.; Grelet, G.A. Nitrogen Storage and Remobilization by Trees: Ecophysiological Relevance in a Changing World. Tree Physiol. 2010, 30, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Keitel, C.; Matzarakis, A.; Rennenberg, H.; Gessler, A. Carbon Isotopic Composition and Oxygen Isotopic Enrichment in Phloem and Total Leaf Organic Matter of European Beech (Fagus sylvatica L.) along a Climate Gradient. Plant Cell Environ. 2006, 29, 1492–1507. [Google Scholar] [CrossRef]
- Körner, C.; Farquhar, G.D.; Wong, S.C. Carbon Isotope Discrimination by Plants Follows Latitudinal and Altitudinal Trends. Oecologia 1991, 88, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Hultine, K.R.; Marshall, J.D. Altitude Trends in Conifer Leaf Morphology and Stable Carbon Isotope Composition. Oecologia 2000, 123, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.R.; McGrath, J.F.; Adams, M.A. Water Availability and Carbon Isotope Discrimination in Conifers. Oecologia 2001, 127, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Fotelli, M.N.; Rienks, M.; Rennenberg, H.; Geßler, A. Climate and Forest Management Affect 15N-Uptake, N Balance and Biomass of European Beech Seedlings. Trees—Struct. Funct. 2004, 18, 157–166. [Google Scholar] [CrossRef]
- Peuke, A.D.; Rennenberg, H. Carbon, Nitrogen, Phosphorus, and Sulphur Concentration and Partitioning in Beech Ecotypes (Fagus sylvatica L.): Phosphorus Most Affected by Drought. Trees 2004, 18, 639–648. [Google Scholar] [CrossRef]
- Rennenberg, H.; Loreto, F.; Polle, A.; Brilli, F.; Fares, S.; Beniwal, R.S.; Gessler, A. Physiological Responses of Forest Trees to Heat and Drought. Plant Biol. 2006, 8, 556–571. [Google Scholar] [CrossRef] [PubMed]
- Fotelli, M.N.; Rennenberg, H.; Geßler, A. Effects of Drought on the Competitive Interference of an Early Successional Species (Rubus fruticosus) on Fagus sylvatica L. Seedlings: 15N Uptake and Partitioning, Responses of Amino Acids and Other, N. Compounds. Plant Biol. 2002, 4, 311–320. [Google Scholar] [CrossRef]
- Meier, I.C.; Leuschner, C. Belowground Drought Response of European Beech: Fine Root Biomass and Carbon Partitioning in 14 Mature Stands across a Precipitation Gradient. Glob. Chang. Biol. 2008, 14, 2081–2095. [Google Scholar] [CrossRef]
- Bertness, M.D.; Callaway, R. Positive Interactions in Communities. Trends Ecol. Evol. 1994, 9, 187–191. [Google Scholar] [CrossRef]
- Morin, X.; Fahse, L.; Scherer-Lorenzen, M.; Bugmann, H. Tree Species Richness Promotes Productivity in Temperate Forests through Strong Complementarity between Species. Ecol. Lett. 2011, 14, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Ishii, H.; Asano, S. The Role of Crown Architecture, Leaf Phenology and Photosynthetic Activity in Promoting Complementary Use of Light among Coexisting Species in Temperate Forests. Ecol. Res. 2010, 25, 715–722. [Google Scholar] [CrossRef]
- Sapijanskas, J.; Paquette, A.; Potvin, C.; Kunert, N.; Loreau, M. Tropical Tree Diversity Enhances Light Capture through Plastic Architectural Changes and Spatial and Temporal Niche Differences. Ecology 2014, 95, 2479–2492. [Google Scholar] [CrossRef]
- Forrester, D.I.; Ammer, C.; Annighöfer, P.J.; Barbeito, I.; Bielak, K.; Bravo-Oviedo, A.; Coll, L.; del Río, M.; Drössler, L.; Heym, M.; et al. Effects of Crown Architecture and Stand Structure on Light Absorption in Mixed and Monospecific Fagus sylvatica and Pinus sylvestris Forests along a Productivity and Climate Gradient through Europe. J. Ecol. 2018, 106, 746–760. [Google Scholar] [CrossRef]
- Berger, T.W.; Untersteiner, H.; Toplitzer, M.; Neubauer, C. Nutrient Fluxes in Pure and Mixed Stands of Spruce (Picea abies) and Beech (Fagus sylvatica). Plant Soil 2009, 322, 317–342. [Google Scholar] [CrossRef]
- Rothe, A.; Ewald, J.; Hibbs, D.E. Do Admixed Broadleaves Improve Foliar Nutrient Status of Conifer Tree Crops? For. Ecol. Manag. 2003, 172, 327–338. [Google Scholar] [CrossRef]
- Maire, V.; Wright, I.J.; Prentice, I.C.; Batjes, N.H.; Bhaskar, R.; van Bodegom, P.M.; Cornwell, W.K.; Ellsworth, D.; Niinemets, Ü.; Ordonez, A.; et al. Global Effects of Soil and Climate on Leaf Photosynthetic Traits and Rates. Glob. Ecol. Biogeogr. 2015, 24, 706–717. [Google Scholar] [CrossRef]
- Rennenberg, H.; Dannenmann, M. Nitrogen Nutrition of Trees in Temperate Forests—The Significance of Nitrogen Availability in the Pedosphere and Atmosphere. Forests 2015, 6, 2820–2835. [Google Scholar] [CrossRef] [Green Version]
- Tarvainen, L.; Lutz, M.; Räntfors, M.; Näsholm, T.; Wallin, G. Increased Needle Nitrogen Contents Did Not Improve Shoot Photosynthetic Performance of Mature Nitrogen-Poor Scots Pine Trees. Front. Plant Sci. 2016, 7, 1051. [Google Scholar] [CrossRef] [PubMed]
- Nambiar, E.K.S.; Fife, D.N. Growth and Nutrient Retranslocation in Needles of Radiata Pine in Relation to Nitrogen Supply. Ann. Bot. 1987, 60, 147–156. [Google Scholar] [CrossRef]
- Millard, P.; Proe, M.F. Storage and Internal Cycling of Nitrogen in Relation To Seasonal Growth of Sitka Spruce. Tree Physiol. 1992, 10, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Millard, P.; Hester, A.; Wendler, R.; Baillie, G. Interspecific Defoliation Responses of Trees Depend on Sites of Winter Nitrogen Storage. Funct. Ecol. 2001, 15, 535–543. [Google Scholar] [CrossRef]
- Sadanandan Nambiar, E.K.; Bowen, G.D. Uptake, Distribution and Retranslocation of Nitrogen by Pinus radiata from 15N-Labelled Fertilizer Applied to Podzolized Sandy Soil. For. Ecol. Manag. 1986, 15, 269–284. [Google Scholar] [CrossRef]
- Camm, E. Photosynthetic Responses in Developing and Year-Old Douglas-Fir Needles during New Shoot Development. Trees 1993, 8, 61–66. [Google Scholar] [CrossRef]
- Millard, P.; Sommerkorn, M.; Grelet, G.A. Environmental Change and Carbon Limitation in Trees: A Biochemical, Ecophysiological and Ecosystem Appraisal. New Phytol. 2007, 175, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Chapin, E.S.; Kedrowski, R.A. Seasonal Changes in Nitrogen and Phosphorus Fractions and Autumn Retranslocation in Evergreen and Deciduous Taiga Trees. Ecology 1983, 64, 376–391. [Google Scholar] [CrossRef]
- Stephens, D.W.; Millard, P.; Turnbull, M.H.; Whitehead, D. The Influence of Nitrogen Supply on Growth and Internal Recycling of Nitrogen in Young Nothofagus fusca Trees. Funct. Plant Biol. 2001, 28, 249–255. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pure Beech | LC13 | R13C | Elevation | Temp | |
| L13C | |||||
| R13C | −0.32 ** | ||||
| Elevation | −0.04 | 0.42 **** | |||
| Temp | 0.08 | −0.39 **** | −0.58 **** | ||
| Prec | −0.04 | 0.42 **** | 1.00 **** | −0.58 **** | |
| Mixed Beech | LC13 | R13C | Elevation | Temp | |
| L13 | |||||
| R13C | 0.2 | ||||
| Elevation | 0.40 ** | 0.59 **** | |||
| Temp | −0.46 *** | −0.77 **** | −0.53 **** | ||
| Prec | 0.40 ** | 0.59 **** | 1.00 **** | −0.53 **** | |
| Mixed Fir | LY13C | CY13C | R13C | Elevation | Temp |
| LY13C | |||||
| CY13C | 0.74 **** | ||||
| R13C | 0.05 | 0.17 | |||
| Elevation | 0.66 **** | 0.71 **** | 0.25 | ||
| Temp | −0.46 *** | −0.40 ** | −0.47 *** | −0.67 **** | |
| Prec | 0.66 **** | 0.71 **** | 0.25 | 1.00 **** | −0.67 **** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magh, R.-K.; Yang, F.; Rehschuh, S.; Burger, M.; Dannenmann, M.; Pena, R.; Burzlaff, T.; Ivanković, M.; Rennenberg, H. Nitrogen Nutrition of European Beech Is Maintained at Sufficient Water Supply in Mixed Beech-Fir Stands. Forests 2018, 9, 733. https://doi.org/10.3390/f9120733
Magh R-K, Yang F, Rehschuh S, Burger M, Dannenmann M, Pena R, Burzlaff T, Ivanković M, Rennenberg H. Nitrogen Nutrition of European Beech Is Maintained at Sufficient Water Supply in Mixed Beech-Fir Stands. Forests. 2018; 9(12):733. https://doi.org/10.3390/f9120733
Chicago/Turabian StyleMagh, Ruth-Kristina, Fengli Yang, Stephanie Rehschuh, Martin Burger, Michael Dannenmann, Rodica Pena, Tim Burzlaff, Mladen Ivanković, and Heinz Rennenberg. 2018. "Nitrogen Nutrition of European Beech Is Maintained at Sufficient Water Supply in Mixed Beech-Fir Stands" Forests 9, no. 12: 733. https://doi.org/10.3390/f9120733
APA StyleMagh, R. -K., Yang, F., Rehschuh, S., Burger, M., Dannenmann, M., Pena, R., Burzlaff, T., Ivanković, M., & Rennenberg, H. (2018). Nitrogen Nutrition of European Beech Is Maintained at Sufficient Water Supply in Mixed Beech-Fir Stands. Forests, 9(12), 733. https://doi.org/10.3390/f9120733