Effects of Nutrient Deficiency on Root Morphology and Nutrient Allocation in Pistacia chinensis Bunge Seedlings


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Measurement of Seedling Morphology and Tissue Nutrient Concentration
2.3. Data Calculations
2.4. Statistical Analyses
3. Results
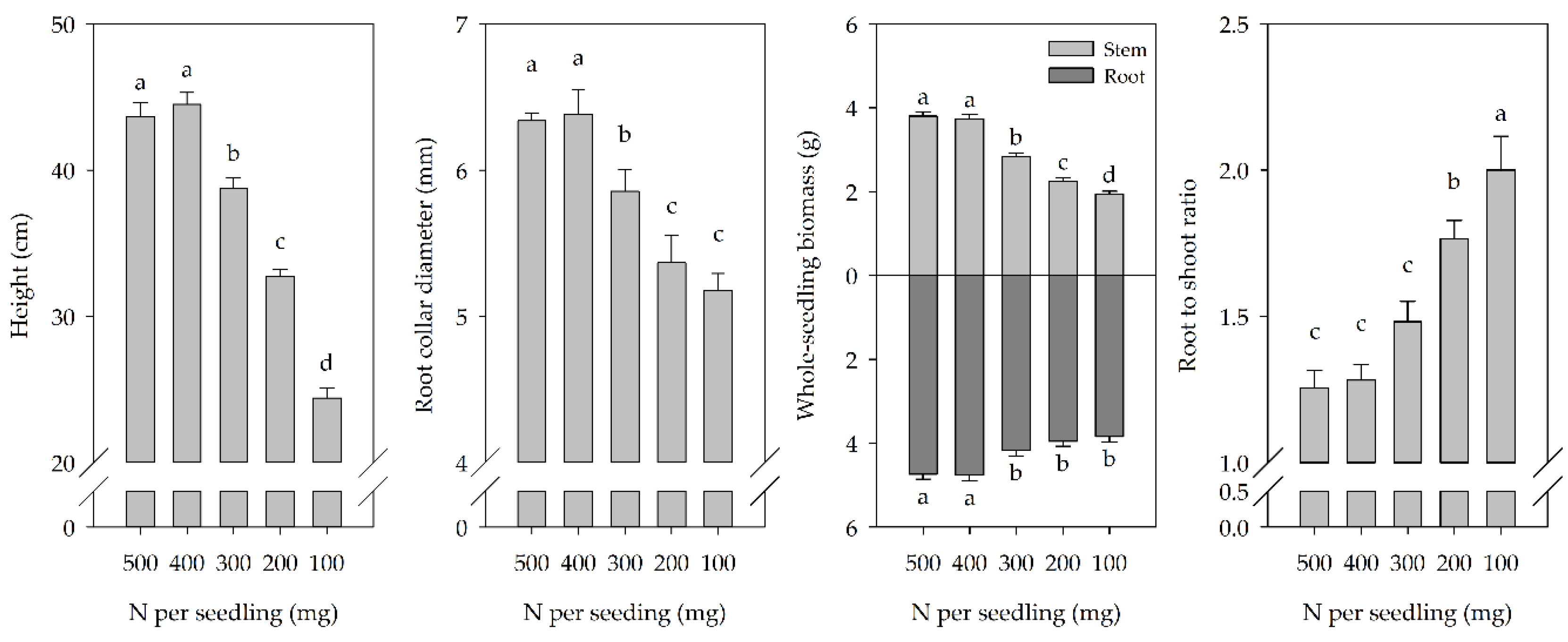
3.1. Seedling Morphology
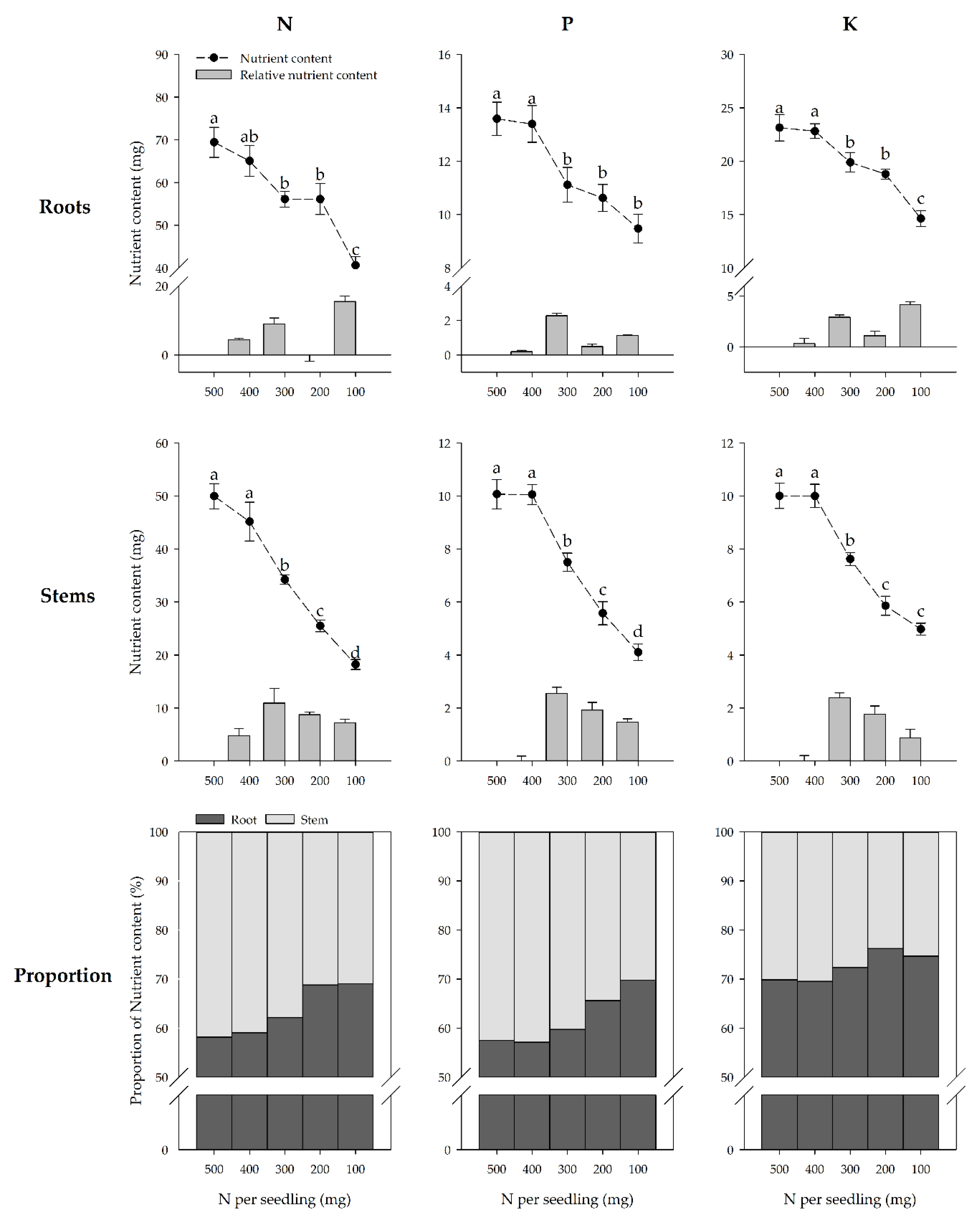
3.2. Nutrient Concentration of the Storage Organs
3.3. Nutrient Accumulation of the Storage Organs
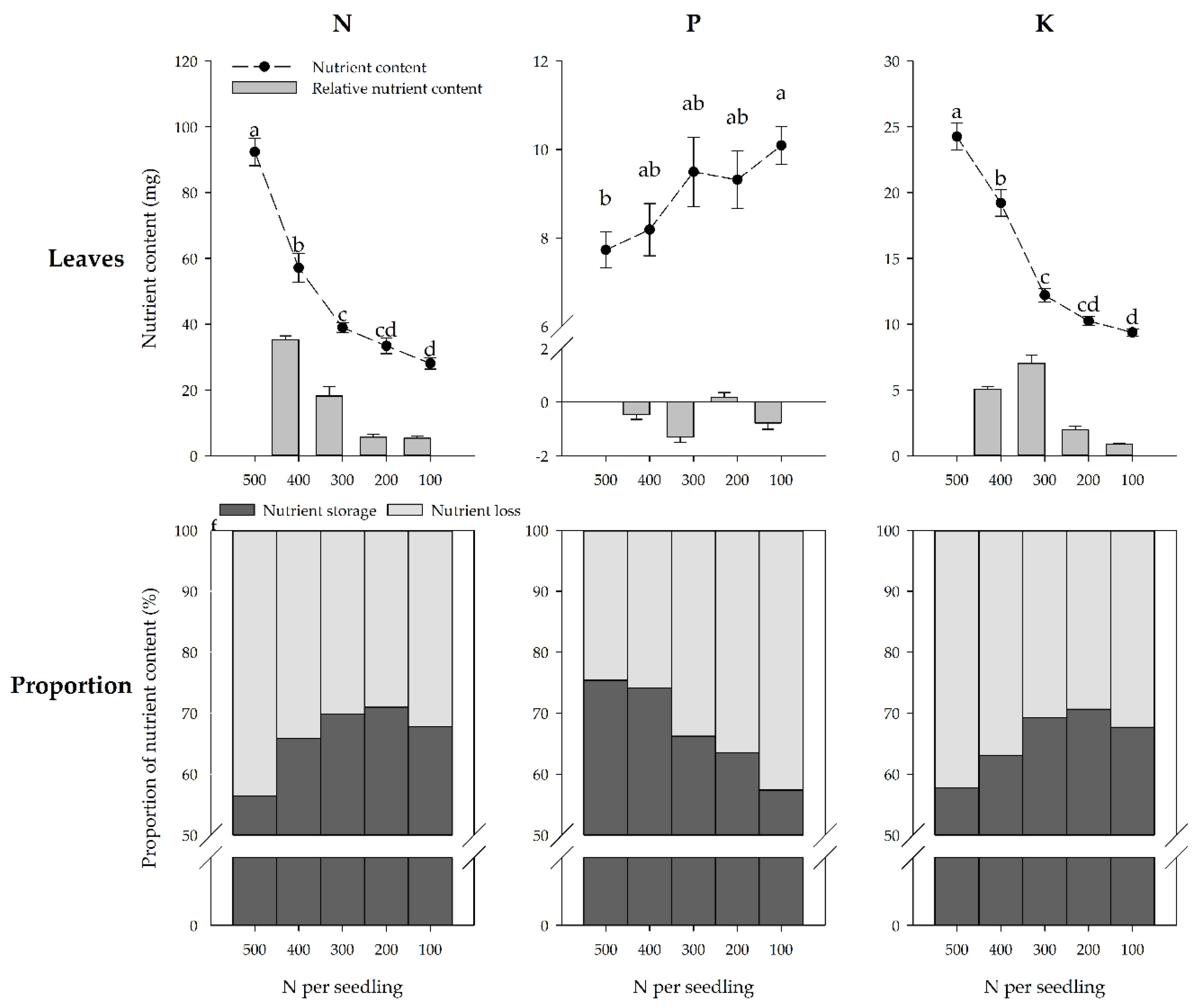
3.4. Nutrient Loss of the Seedlings
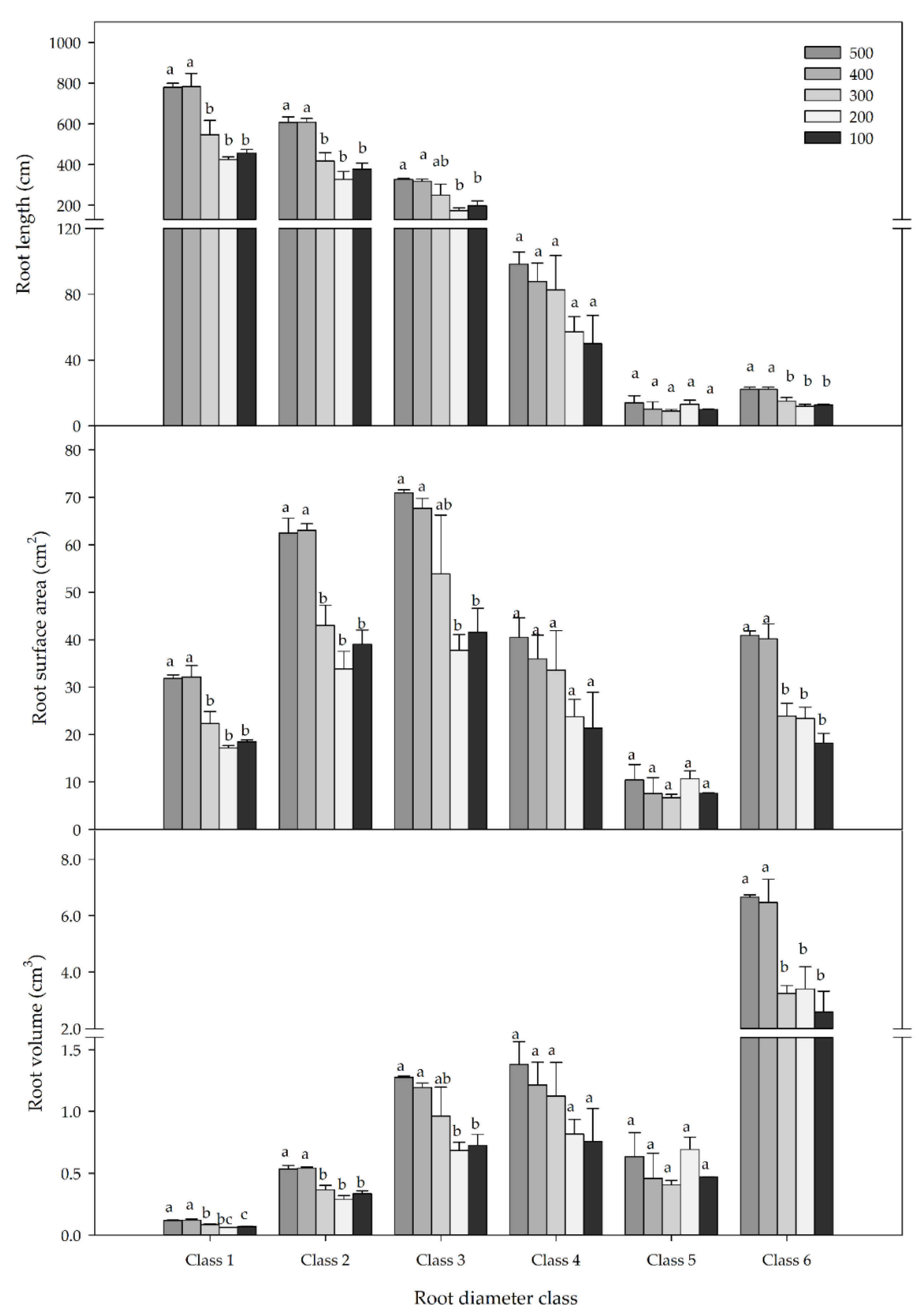
3.5. Root Morphology
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Taiz, L.; Zeiger, E. Plant Physiology, 4th ed.; Sinauer Associates: Sunderland, UK, 2006. [Google Scholar]
- Egilla, J.N.; Davies, F.T.; Drew, M.C. Effect of potassium on drought resistance of Hibiscus rosa-sinensis cv. Leprechaun: Plant growth, leaf macro- and micronutrient content and root longevity. Plant Soil 2001, 229, 213–224. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Uscola, M.; Jacobs, D.F. The role of stored carbohydrates and nitrogen in the growth and stress tolerance of planted forest trees. New For. 2015, 46, 813–839. [Google Scholar] [CrossRef]
- Wang, J.; Villar-Salvador, P.; Li, G.; Liu, Y. Moderate water stress does not inhibit nitrogen remobilization, allowing fast growth in high nitrogen content Quercus variabilis seedlings under dry conditions. Tree Physiol. 2018, 39, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Luoranen, J.; Rikala, R. Nutrient loading of Norway spruce seedlings hastens bud burst and enhances root growth after outplanting. Silva Fenn. 2011, 45, 319–329. [Google Scholar] [CrossRef]
- Adams, T.S.; Mccormack, M.L.; Eissenstat, D.M. Foraging strategies in trees of different root morphology: The role of root lifespan. Tree Physiol. 2013, 33, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Goebel, M.; Hobbie, S.E.; Bulaj, B.; Zadworny, M.; Archibald, D.D.; Oleksyn, J.; Reich, P.B.; Eissenstat, D.M. Decomposition of the finest root branching orders: Linking belowground dynamics to fine-root function and structure. Ecol. Monogr. 2011, 81, 89–102. [Google Scholar] [CrossRef]
- Doi, R.; Tanikawa, T.; Miyatani, K.; Hirano, Y. Intraspecific variation in morphological traits of root branch orders in Chamaecyparis obtusa. Plant Soil 2017, 416, 503–513. [Google Scholar] [CrossRef]
- Hishi, T.; Tateno, R.; Takeda, H. Anatomical characteristics of individual roots within the fine-root architecture of Chamaecyparis obtusa (Sieb. & Zucc.) in organic and mineral soil layers. Ecol. Res. 2006, 21, 754–758. [Google Scholar]
- Makita, N.; Hirano, Y.; Mizoguchi, T.; Kominami, Y.; Dannoura, M.; Ishii, H.; Finér, L.; Kanazawa, Y. Very fine roots respond to soil depth: Biomass allocation, morphology, and physiology in a broad-leaved temperate forest. Ecol. Res. 2011, 26, 95–104. [Google Scholar] [CrossRef]
- Volder, A.; Smart, D.R.; Bloom, A.J.; Eissenstat, D.M. Rapid decline in nitrate uptake and respiration with age in fine lateral roots of grape: Implications for root efficiency and competitive effectiveness. New Phytol. 2005, 165, 493–502. [Google Scholar] [CrossRef]
- Makita, N.; Hirano, Y.; Dannoura, M.; Kominami, Y.; Mizoguchi, T.; Ishii, H.; Kanazawa, Y. Fine root morphological traits determine variation in root respiration of Quercus serrata. Tree Physiol. 2009, 29, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Liu, F.; Xue, S. Nitrogen addition enhanced water uptake by affecting fine root morphology and coarse root anatomy of Chinese pine seedlings. Plant Soil 2017, 418, 177–189. [Google Scholar] [CrossRef]
- Nguyen, P.V.; Dickmann, D.I.; Pregitzer, K.S.; Hendrick, R. Late-season changes in allocation of starch and sugar to shoots, coarse roots, and fine roots in two hybrid poplar clones. Tree Physiol. 1990, 7, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.S.; Jacobs, D.F. Quantifying root system quality of nursery seedlings and relationship to outplanting performance. New For. 2005, 30, 295–311. [Google Scholar] [CrossRef]
- Markesteijn, L.; Poorter, L. Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought- and shade-tolerance. J. Ecol. 2009, 97, 311–325. [Google Scholar] [CrossRef]
- Mcinenly, L.E.; Merrill, E.; Cahill, J.F.; Juma, N.G. Festuca campestris alters root morphology and growth in response to simulated grazing and nitrogen form: Defoliation, N-form and fescue roots. Funct. Ecol. 2010, 24, 283–292. [Google Scholar] [CrossRef]
- Zangaro, W.; Nishidate, F.R.; Camargo, F.R.S.; Romagnoli, G.G.; Vandressen, J. Relationships among Arbuscular Mycorrhizas, Root morphology and seedling growth of tropical native woody species in southern Brazil. J. Trop. Ecol. 2005, 21, 529–540. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef]
- Cao, X.; Chen, C.; Zhang, D.; Shu, B.; Xiao, J.; Xia, R. Influence of nutrient deficiency on root architecture and root hair morphology of trifoliate orange (Poncirus trifoliata L. Raf.) seedlings under sand culture. Sci. Hortic. 2013, 162, 100–105. [Google Scholar] [CrossRef]
- Gao, Y.; Li, X.; Tian, Q.; Wang, B.; Zhang, B. Sulfur deficiency had different effects on Medicago truncatula ecotypes A17 and R108 in terms of growth, root morphology and nutrient contents. J. Plant Nutr. 2016, 39, 301–314. [Google Scholar] [CrossRef]
- Mollier, A.; Pellerin, S. Maize root system growth and development as influenced by phosphorus deficiency. J. Exp. Bot. 1999, 50, 487–497. [Google Scholar] [CrossRef]
- Wan, F.; Ross-Davis, A.L.; Shi, W.; Weston, C.; Song, X.; Chang, X.; Davis, A.S.; Liu, Y.; Teng, F. Subirrigation effects on larch seedling Growth, root morphology, and media chemistry. Forests 2019, 10, 38. [Google Scholar] [CrossRef]
- Schöll, L.V.; Hoffland, E.; Breemen, N.V. Organic acid exudation by ectomycorrhizal fungi and Pinus sylvestris in response to nutrient deficiencies. New Phytol. 2006, 170, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.W.; Dale, J.E. Nitrogen deficiency and fertilization effects on needle growth and photosynthesis in Sitka spruce (Picea sitchensis). Tree Physiol. 1995, 15, 813–817. [Google Scholar] [CrossRef]
- Ciompi, S.; Gentili, E.; Guidi, L.; Soldatini, G.F. The effect of nitrogen deficiency on leaf gas exchange and chlorophyll fluorescence parameters in sunflower. Plant Sci. 1996, 118, 177–184. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Shi, Z.; Balogh-Brunstad, Z.; Grant, M.; Harsh, J.; Gill, R.; Thomashow, L.; Dohnalkova, A.; Stacks, D.; Letourneau, M.; Keller, C.K. Cation uptake and allocation by red pine seedlings under cation-nutrient stress in a column growth experiment. Plant Soil 2014, 378, 83–98. [Google Scholar] [CrossRef]
- Lu, L.; Jiang, D.; Fu, J.; Zhuang, D.; Huang, Y.; Hao, M. Evaluating energy benefit of Pistacia chinensis based biodiesel in China. Renew. Sustain. Energy Rev. 2014, 35, 258–264. [Google Scholar] [CrossRef]
- Zhu, B.; Wang, Q.; Roge, E.F.; Nan, P.; Liu, Z.; Zhong, Y. Chemical variation in leaf oils of Pistacia chinensis from five locations in China. Chem. Nat. Compd. 2006, 42, 422–425. [Google Scholar] [CrossRef]
- Li, X.; He, X.; Li, Z.; Wang, Y.; Wang, C.; Shi, H.; Wang, F. Enzymatic production of biodiesel from Pistacia chinensis bge seed oil using immobilized lipase. Fuel 2012, 92, 89–93. [Google Scholar] [CrossRef]
- Noureen, F.; Khan, M.R.; Shah, N.A.; Khan, R.A.; Naz, K.; Sattar, S. Pistacia chinensis: Strong antioxidant and potent testicular toxicity amelioration agent. Asian Pac. J. Trop. Med. 2017, 10, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Yayeh, T.; Hong, M.; Jia, Q.; Lee, Y.C.; Kim, H.J.; Hyun, K.T.; Rhee, M.H. Pistacia chinensis inhibits NO production and upregulates HO-1 induction via PI-3K/Akt pathway in LPS stimulated macrophage cells. Am. J. Chin. Med. 2012, 40, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Trubat, R.; Cortina, J.; Vilagrosa, A. Plant morphology and root hydraulics are altered by nutrient deficiency in Pistacia lentiscus (L.). Trees 2006, 20, 334–339. [Google Scholar] [CrossRef]
- Lowther, J.R. Use of a single sulphuric acid-hydrogen peroxide digest for the analysis of Pinus radiata needles. Commun. Soil Sci. Plant Anal. 1980, 11, 175–188. [Google Scholar] [CrossRef]
- Salifu, K.F.; Timmer, V.R. Optimizing nitrogen loading of Picea mariana seedlings during nursery culture. Can. J. For. Res. 2003, 33, 1287–1294. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Peñuelas, J.L.; Jacobs, D.F. Nitrogen nutrition and drought hardening exert opposite effects on the stress tolerance of Pinus pinea L. seedlings. Tree Physiol. 2013, 33, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.Y.; Li, L.H.; Han, X.G.; Huang, J.H.; Wan, S.Q. Foliar nitrogen dynamics and nitrogen resorption of a sandy shrub Salix gordejevii in northern China. Plant Soil 2005, 278, 183–193. [Google Scholar] [CrossRef]
- Pallardy, S.G. Physiology of Woody Plants, 3rd ed.; Academic Press: Burlington, NJ, USA, 2008. [Google Scholar]
- Hodge, A. The plastic plant: Root responses to heterogeneous supplies of nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Fircks, Y.V.; Ericsson, T.; Sennerby-Forsse, L. Seasonal variation of macronutrients in leaves, stems and roots of Salix dasyclados Wimm. grown at two nutrient levels. Biomass Bioenergy 2001, 21, 321–334. [Google Scholar] [CrossRef]
- Kurdali, F. Seasonal nitrogen changes in Alnus orientalis and Populus nigra and N2 fixation by exotic alder species in Syria. Commun. Soil Sci. Plant Anal. 2000, 31, 2509–2522. [Google Scholar] [CrossRef]
- Chapin, F.S.; Kedrowski, R.A. Seasonal changes in nitrogen and phosphorus fractions and autumn retranslocation in evergreen and deciduous taiga trees. Ecology 1983, 64, 376–391. [Google Scholar] [CrossRef]
- Keyes, M.R.; Grier, C.C. Above- and below-ground net production in 40-year-old Douglas-fir stands on low and high productivity sites. Can. J. For. Res. 1981, 11, 599–605. [Google Scholar] [CrossRef]
- Bollard, E.G. Nitrogenous compounds in plant xylem sap. Nature 1956, 178, 1189–1190. [Google Scholar] [CrossRef]
- Cooke, J.E.; Weih, M. Nitrogen storage and seasonal nitrogen cycling in Populus: Bridging molecular physiology and ecophysiology. New Phytol. 2005, 167, 19–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wetzel, S.; Demmers, C.; Greenwood, J.S. Spherical organelles, analogous to seed protein bodies, fluctuate seasonally in parenchymatous cells of hardwoods. Can. J. Bot. 1989, 67, 3439–3445. [Google Scholar] [CrossRef]
- Brix, H. Effects of thinning and nitrogen fertilization on growth of Douglas-fir: Relative contribution of foliage quantity and efficiency (Pseudotsuga menziesii). Can. J. For. Res. 1983, 13, 167–175. [Google Scholar] [CrossRef]
- Sheriff, D.W. Gas Exchange of Field-Grown Pinus radiate—Relationships with Foliar Nutrition and Water Potential, and with Climatic Variables. Funct. Plant Biol. 1995, 22, 1015–1026. [Google Scholar] [CrossRef]
- Cakmak, I.; Hengeler, C.; Marschner, H. Changes in phloem export of sucrose in leaves in response to phosphorus, potassium and magnesium deficiency in bean plants. J. Exp. Bot. 1994, 45, 1251–1257. [Google Scholar] [CrossRef]
- Lott, J.N.A.; Ockenden, I.; Raboy, V.; Batten, G.D. Phytic acid and phosphorus in crop seeds and fruits: A global estimate. Seed Sci. Res. 2000, 10, 11–33. [Google Scholar] [CrossRef]
- Seufferheld, M.; Curzi, M.J. Recent discoveries on the roles of polyphosphates in plants. Plant Mol. Biol. Rep. 2010, 28, 549–559. [Google Scholar] [CrossRef]
- Shane, M.W.; Mccully, M.E.; Lambers, H. Tissue and cellular phosphorus storage during development of phosphorus toxicity in Hakea prostrata (Proteaceae). J. Exp. Bot. 2004, 55, 1033–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakie, T. Phosphorus fractions in tobacco plants as affected by phosphate application. Soil Sci. Plant Nutr. 1969, 15, 81–85. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | N (mg) | P (mg) | K (mg) |
|---|---|---|---|
| 500 | 500 | 204 | 386 |
| 400 | 400 | 163 | 309 |
| 300 | 300 | 122 | 231 |
| 200 | 200 | 81 | 154 |
| 100 | 100 | 41 | 77 |
| Organs | Treatments | N Concentration (mg/g) | P Concentration (mg/g) | K Concentration (mg/g) |
|---|---|---|---|---|
| 500 | 14.66 ± 0.74a | 2.87 ± 0.13a | 4.88 ± 0.26a | |
| 400 | 13.66 ± 0.76a | 2.81 ± 0.14a | 4.79 ± 0.15a | |
| Roots | 300 | 13.44 ± 0.44a | 2.66 ± 0.16a | 4.76 ± 0.22a |
| 200 | 14.20 ± 0.91a | 2.69 ± 0.13a | 4.75 ± 0.12a | |
| 100 | 10.59 ± 0.53b | 2.47 ± 0.14a | 3.81 ± 0.20b | |
| 500 | 13.15 ± 0.62a | 2.65 ± 0.14a | 2.63 ± 0.13a | |
| 400 | 12.10 ± 0.98a | 2.69 ± 0.10a | 2.68 ± 0.12a | |
| Stems | 300 | 12.08 ± 0.32a | 2.65 ± 0.12a | 2.69 ± 0.09a |
| 200 | 11.34 ± 0.49a | 2.48 ± 0.19ab | 2.60 ± 0.16a | |
| 100 | 9.43 ± 0.23b | 2.12 ± 0.16b | 2.57 ± 0.12a |
| Source | Root Length | Root Surface Area | Root Volume |
|---|---|---|---|
| Nutrient treatment | 39.4 (<0.001) | 28.6 (<0.001) | 14.2 (<0.001) |
| Root diameter | 464.2 (<0.001) | 83.2 (<0.001) | 168.3 (<0.001) |
| Nutrient treatment × Root diameter | 7.9 (<0.001) | 2.0 (0.019) | 7.2 (<0.001) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, X.; Wan, F.; Chang, X.; Zhang, J.; Sun, M.; Liu, Y. Effects of Nutrient Deficiency on Root Morphology and Nutrient Allocation in Pistacia chinensis Bunge Seedlings. Forests 2019, 10, 1035. https://doi.org/10.3390/f10111035
Song X, Wan F, Chang X, Zhang J, Sun M, Liu Y. Effects of Nutrient Deficiency on Root Morphology and Nutrient Allocation in Pistacia chinensis Bunge Seedlings. Forests. 2019; 10(11):1035. https://doi.org/10.3390/f10111035
Chicago/Turabian StyleSong, Xiehai, Fangfang Wan, Xiaochao Chang, Jin Zhang, Minghui Sun, and Yong Liu. 2019. "Effects of Nutrient Deficiency on Root Morphology and Nutrient Allocation in Pistacia chinensis Bunge Seedlings" Forests 10, no. 11: 1035. https://doi.org/10.3390/f10111035
APA StyleSong, X., Wan, F., Chang, X., Zhang, J., Sun, M., & Liu, Y. (2019). Effects of Nutrient Deficiency on Root Morphology and Nutrient Allocation in Pistacia chinensis Bunge Seedlings. Forests, 10(11), 1035. https://doi.org/10.3390/f10111035