Impact of Severe Drought during the Strong 2015/2016 El Nino on the Phenology and Survival of Secondary Dry Dipterocarp Species in Western Thailand

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
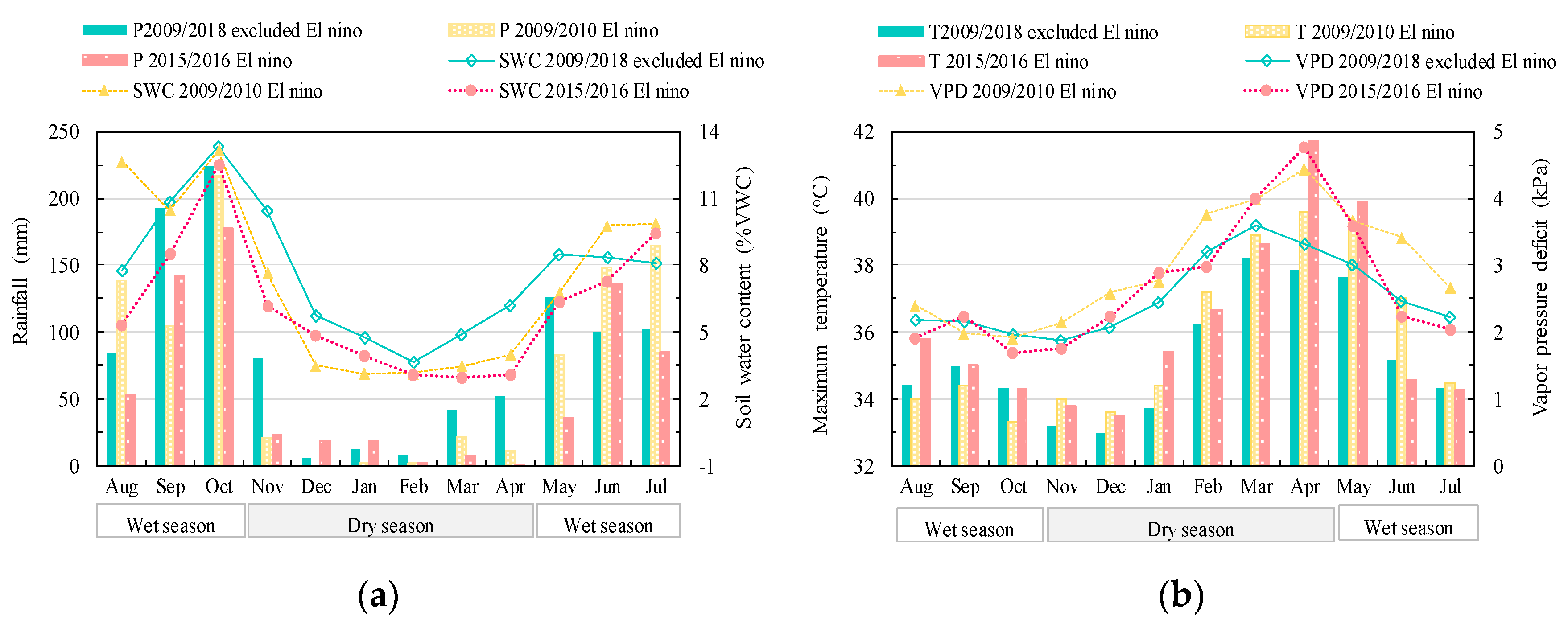
2.2. Microclimate Measurement
2.3. Sampling Design, Forest Structural Characteristic and Biodiversity
2.4. Phenological Indices
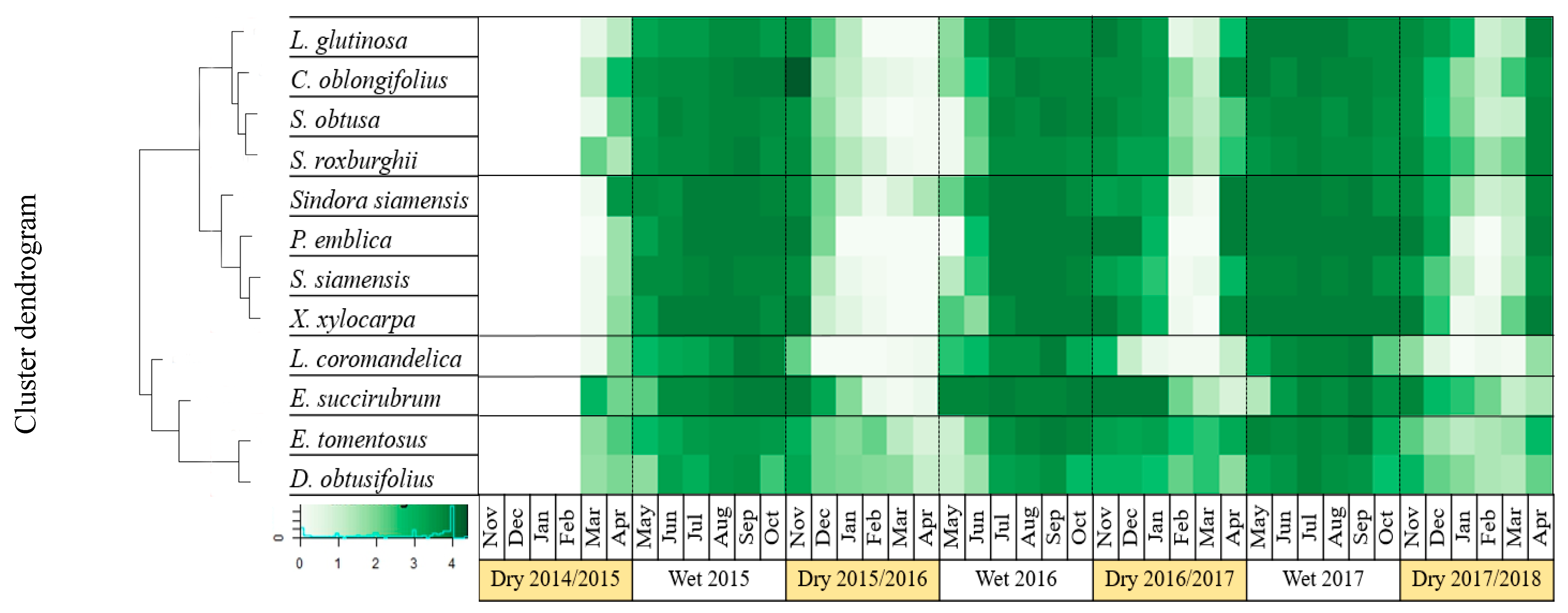
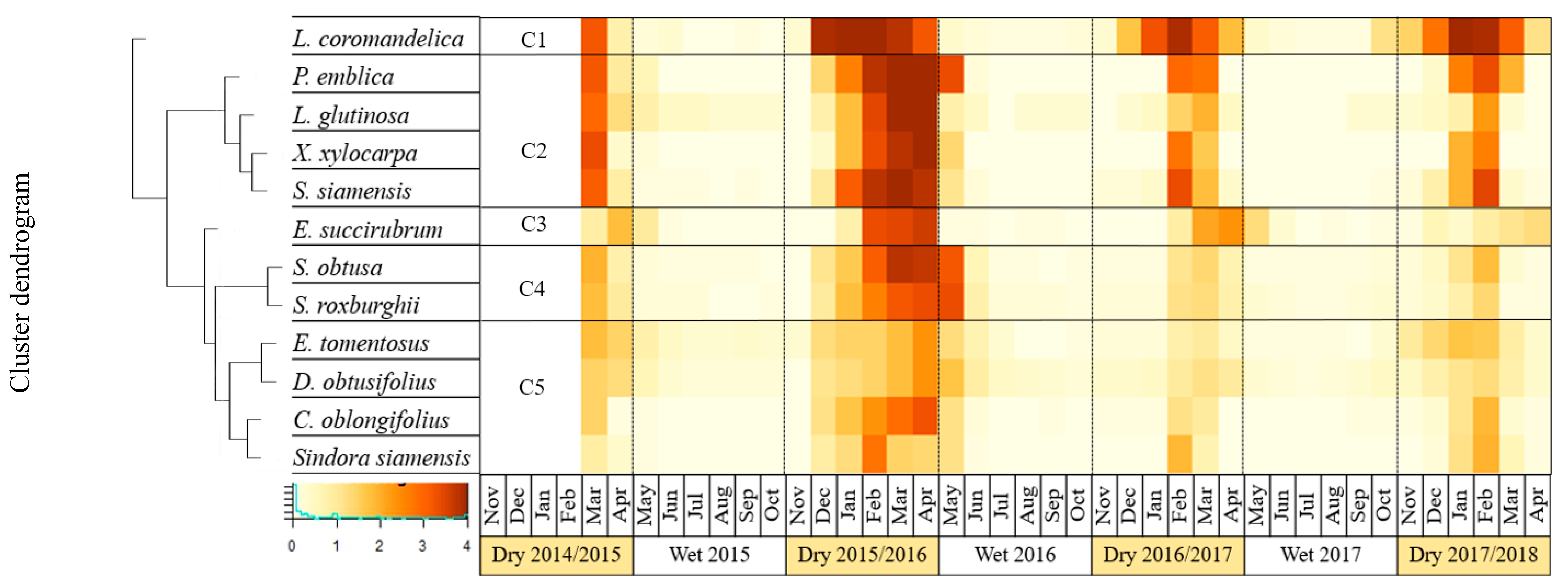
2.5. Clustering of Leaf Phenology
2.6. Wood Density
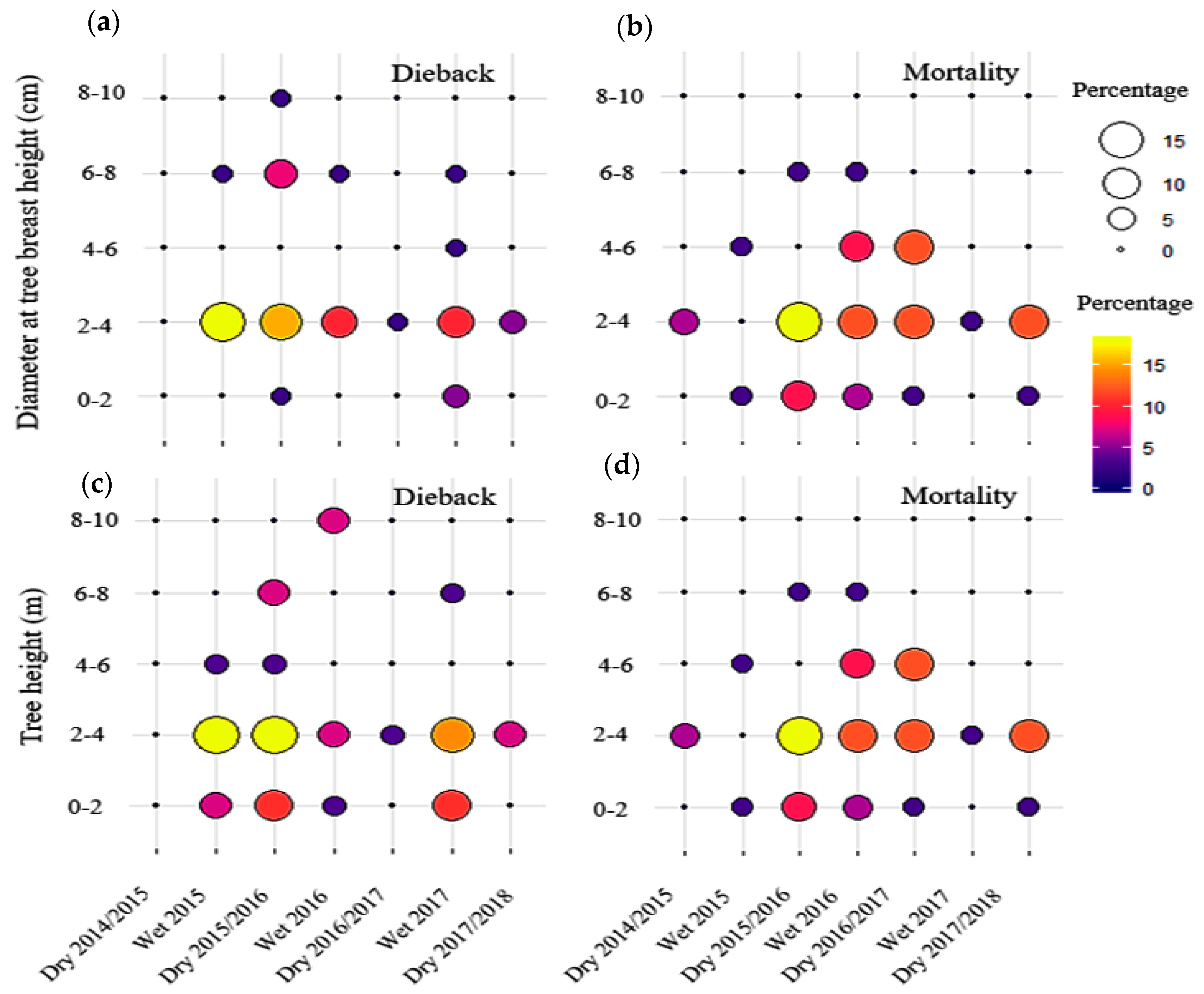
2.7. Die Back and Mortality
2.8. Data Analysis
3. Results
3.1. Impact of ENSO on the Microclimate of the Study DDF
3.2. Impact of El Niño on Leaf Phenology of the DDF
3.3. Relationship between Microclimate Conditions and the Degree of Deciduousness in the DDF
3.4. First Leaf Expansion and Leaf Litter Production of the Dominant Species
3.5. Wood Density
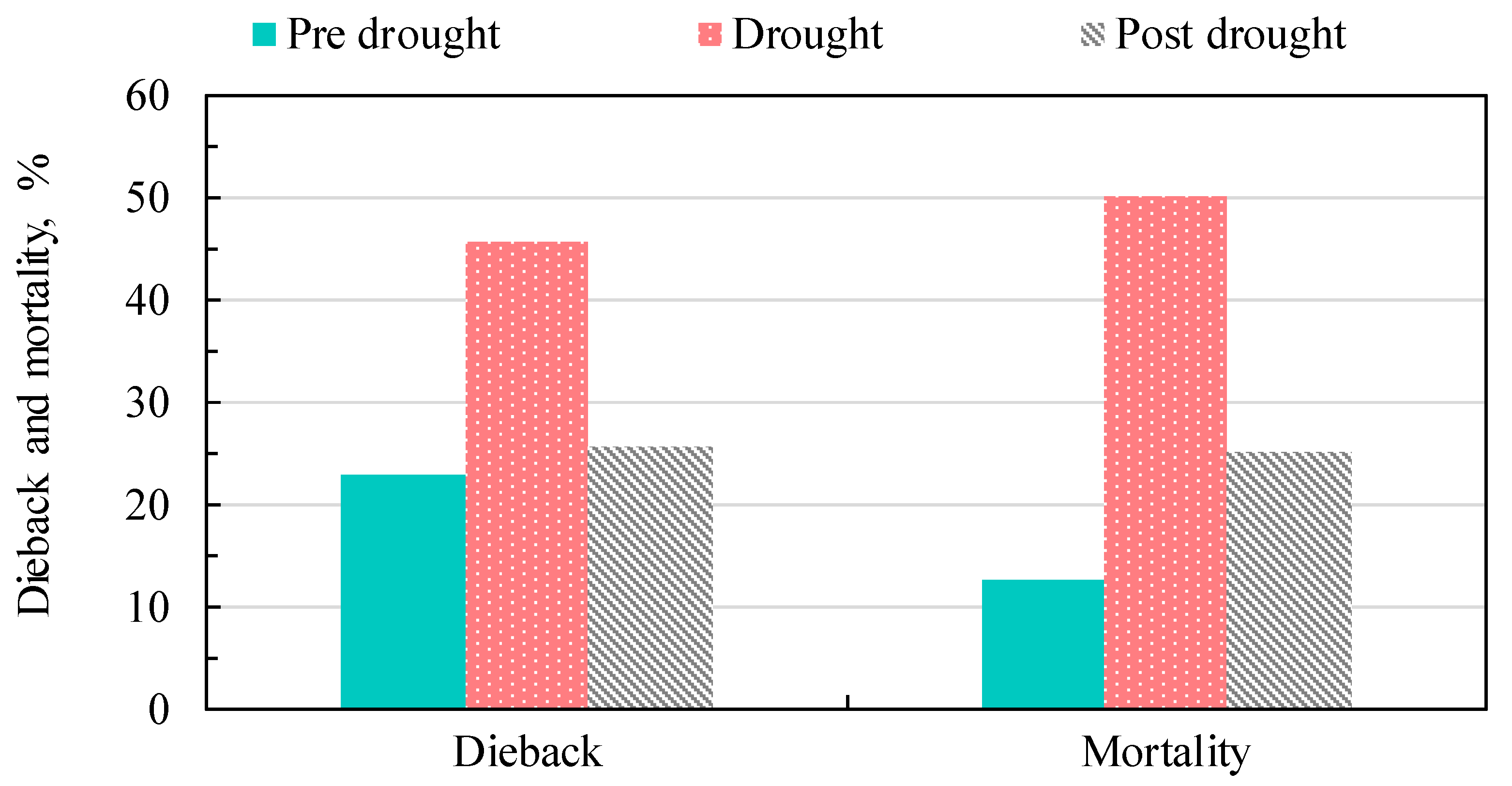
3.6. Dieback and Mortality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fu, Y.H.; Zhao, H.; Piao, S.; Peaucelle, M.; Peng, S.; Zhou, G.; Ciais, P.; Huang, M.; Menzel, A.; Peñuelas, J.; et al. Declining Global Warming Effects on the Phenology of Spring Leaf Unfolding. Res. Lett. 2015, 526, 104–106. [Google Scholar] [CrossRef] [PubMed]
- Vitasse, Y.; Signarbieux, C.; Fu, Y.H. Global Warming Leads to More Uniform Spring Phenology across Elevations. Proc. Natl. Acad. Sci. USA 2017, 115, 1004–1005. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.; Dupuy, M.; Gei, M.G.; Hulshof, C.; Medvigy, D.; Pizano, C.; Waring, B.G. Will Seasonally Dry Tropical Forests be Sensitive or Resistant to Future Changes in Rainfall Regimes? Environ. Res. Lett. 2017, 12, 023001. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate Change, Phenology, and Phenological Control of Vegetation Feedbacks to the Climate System. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Cleland, E.E.; Chuine, I.; Menzel, A.; Mooney, H.; Schwartz, M.D. Shifting Plant Phenology in Response to Global Change. Trends Ecol. Evol. 2007, 22, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Borchert, R. Induction of Hydration and Bud Break by Irrigation or Rain in Deciduous Trees of a Tropical Dry Forest in Costa Rica. Trees 1994, 8, 198–204. [Google Scholar] [CrossRef]
- Valdez-Hernández, M.; Andrade, J.L.; Jackson, P.C.; Rebolledo-Vieyra, M. Phenology of Five Tree Species of a Tropical Dry Forest in Yucatan, Mexico: Effects of Environmental and Physiological Factors. Plant Soil 2010, 329, 155–171. [Google Scholar] [CrossRef]
- Räsänen, T.A.; Lindgren, V.; Guillaume, J.H.A.; Buckley, B.M.; Kummu, M. On the Spatial and Temporal Variability of ENSO Precipitation and Drought Teleconnection in Mainland Southeast Asia. Clim. Past 2016, 12, 1889–1905. [Google Scholar] [CrossRef]
- Rahmat, R.; Archevarahuprok, B.; Kang, C.P. A Regional Climate Modelling Experiment for Southeast Asia: Using PRECIS Regional Climate Model and Selected CMIP3 Global Climate Models; Centre for Climate Research Singapore of the Meteorological Service Singapore: Singapore, 2014.
- Cai, W.; Borlance, S.; Lengaigne, M.; Van Rensch, P.; Collins, M.; Vecchi, G.; England, M.H. Increasing Frequency of Extreme El Niño Events due to Greenhouse Warming. Nat. Clim. Chang. 2014, 4, 111–116. [Google Scholar] [CrossRef]
- Loo, Y.Y.; Billa, L.; Singh, A. Effect of Climate Change on Seasonal Monsoon in Asia and its Impact on the Variability of Monsoon Rainfall in Southeast Asia. Geosci. Front. 2014, 6, 817–823. [Google Scholar] [CrossRef]
- Sooktawee, S.; Humphries, U.; Limsakul, A.; Wongwises, P. Possible Impacts of Northeast Monsoon on Precipitation and Temperature over the Indochina Peninsula. In Proceedings of the Second Environment Asia International Conference on Human Vulnerability and Global Environmental Change, Chonburi, Thailand, 15–17 May 2013. [Google Scholar]
- Murphy, P.G.; Lugo, A.E. Ecology of tropical dry forest. Annu. Rev. Ecol. Evol. Syst. 1986, 17, 67–88. [Google Scholar] [CrossRef]
- Borchert, R. Soil and Stem Water Storage Determine Phenology and Distribution of Tropical Dry Forest Trees. Ecology 1994, 75, 1437–1449. [Google Scholar] [CrossRef]
- Chen, X.; Wang, L.; Inouye, D. Delayed Response of Spring Phenology to Global Warming in Subtropics and Tropics. Agric. For. Meteorol. 2017, 234, 222–235. [Google Scholar] [CrossRef]
- Stan, K.; Sanchez-Azofeifa, A. Tropical Dry Forest Diversity, Climatic Response, and Resilience in a Changing Climate. Forests 2019, 10, 443. [Google Scholar] [CrossRef]
- Sakai, S.; Kitajima, K. Tropical Phenology: Recent Advances and perspectives. Ecol. Res. 2019, 34, 50–54. [Google Scholar] [CrossRef]
- Khamyong, N.; Wangpakapattanawong, P.; Chairuangsri, S.; Inta, A.; Tiansawat, P. Tree Species Composition and Height-Diameter Allometry of Three Forest Types in Northern Thailand. CMU J. Nat. Sci. 2018, 17, 289–306. [Google Scholar] [CrossRef]
- Wohlfart, C.; Wegmann, M.; Leimgruber, P. Mapping Threatened Dry Deciduous Dipterocarp Forest in Southeast Asia for Conservation Management. Trop. Conserv. Sci. 2014, 7, 597–613. [Google Scholar] [CrossRef]
- U.S. DOE (Department of Energy). Research Priorities for Tropical Ecosystems under Climate Change Workshop Report, DOE/SC-0153 U.S.; Department of Energy Office of Science: Germantown, MD, USA, 2012.
- Nguyen, T.T.; Baker, P.J. Structure and Composition of Deciduous Dipterocarp Forests in Central Vietnam: Patterns of Species’ Dominance and Regeneration Failure. Plant Ecol. Divers. 2016, 9, 589–601. [Google Scholar] [CrossRef]
- Rundel, P.; Boonpragob, K.; Patterson, M. Seasonal Water Relations and Leaf Temperature in a Deciduous Dipterocarp Forest in Northeastern Thailand. Forests 2017, 8, 368. [Google Scholar] [CrossRef]
- Estoque, R.C.; Gomi, K.; Togawa, T.; Ooba, M.; Hijioka, Y.; Akiyama, C.M.; Kuroda, K. The Future of Southeast Asia’s Forests. Nat. Commun. 2019, 692, 903–916. [Google Scholar] [CrossRef]
- Powers, J.S.; Marín-Spiotta, E. Ecosystem Processes and Biogeochemical Cycles in Secondary Tropical Forest Succession. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 497–519. [Google Scholar] [CrossRef]
- Borchert, R. Responses of Tropical Trees to Rainfall Seasonally and Its Long-Term Changes. Clim. Chang. 1998, 39, 381–393. [Google Scholar] [CrossRef]
- Lopez-Iglesias, B.; Villar, R.; Poorter, L. Functional Traits Predict Drought Performance and Distribution of Mediterranean Woody Species. Acta Oecol. 2014, 56, 10–18. [Google Scholar] [CrossRef]
- Worbes, M.; Blanchart, S.; Fichtler, E. Relations between Water Balance, Wood Traits and Phenological Behavior of Tree Species from a Tropical Dry Forest in Costa Rica-a Multifactorial Study. Tree Physiol. 2013, 33, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Markesteijn, L.; Poorter, L.; Paz, H.; Sack, L.; Bongers, F. Ecological Differentiation in Xylem Cavitation Resistance is Associated with Stem and Leaf Structural Traits. Plant Cell Environ. 2011, 34, 137–148. [Google Scholar] [CrossRef]
- Kenzo, T.; Sano, M.; Yoneda, R.; Chann, S. Comparison of Wood Density and Water Content between Dry Evergreen and Dry Deciduous Forest Trees in Central Cambodia. Jpn. Agric. Res. Q. 2017, 51, 363–374. [Google Scholar] [CrossRef]
- Itoh, A.; Nanami, S.; Harata, T.; Ohkubo, T.; Tan, S.; Chong, L.; Yamakura, T. The Effect of Habitat Association and Edaphic Conditions on Tree Mortality during El Niño-induced Drought in a Bornean Dipterocarp Forest. Biotropica 2012, 44, 606–617. [Google Scholar] [CrossRef]
- Nakagawa, M.; Tanaka, K.; Nakashizuka, T.; Ohkubo, T.; Kato, T.; Maeda, T.; Teo, S. Impact of Severe Drought Associated with the 1997–1998 El Niño in a Tropical Forest in Sarawak. J. Trop. Ecol. 2000, 16, 355–367. [Google Scholar] [CrossRef]
- Rodriguez-Zaccaro, F.D.; Groover, A. Wood and Water: How Trees Modify Wood Development to Cope with Drought. Plants People Planet 2019. [Google Scholar] [CrossRef]
- Álvarez-Yépiz, J.C.; Martínez-Yrízar, A.; Fredericksen, T.S. Special Issue: Resilience of Tropical Dry Forests to Extreme Disturbance Events. For. Ecol. Manag. 2018, 426, 1–6. [Google Scholar] [CrossRef]
- L’Heureux, M.L.; Takahashi, K.; Watkins, A.B.; Barnston, A.G.; Becker, E.J.; Di Liberto, T.E.; Mosquera-Vásquez, K. Observing and Predicting the 2015/16 El Niño. Bull. Am. Meteorol. Soc. 2017, 98, 1363–1382. [Google Scholar] [CrossRef]
- Hanpattanakit, P. Temporal Variations of Soil Respiration in a Dry Dipterocarp Forest. Ph.D. Thesis, King Mongkut’s University of Technology Thonburi, Bangkok, Thailand, 2013. [Google Scholar]
- Phillips, E.A. Methods of Vegetation Study; Henery Halt and Co. Inc.: New York, NY, USA, 1959; p. 107. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; Communication Research; The University of Illinois Press: Urbana, IL, USA, 1964; p. 125. [Google Scholar]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Hill, M.O. Diversity and Evenness: A Unify, Notation and its Consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef]
- Kuaraksa, C.; Elliott, S.; Hossaert-Mckey, M. The Phenology of Dioecious Ficus spp. Tree Species and its Importance for Forest Restoration Projects. For. Ecol. Manag. 2012, 265, 82–93. [Google Scholar] [CrossRef]
- Kassambara, A. Determining the Optimal Number of Clusters: 3 Must Know Methods. Available online: https://www.datanovia.com/en/lessons/determining-the-optimal-number-of-clusters-3-must-know-methods/ (accessed on 30 November 2018).
- Murtagh, F.; Legendre, P. Ward’s Hierarchical Agglomerative Clustering Method: Which Algorithms Implement Ward’s Criterion? J. Classif. 2014, 31, 275–295. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Pittermann, J. Drought Experience and Cavitation Resistance in Six Shrubs from the Great Basin, Utah. Basic Appl. Ecol. 2000, 1, 31–41. [Google Scholar] [CrossRef]
- NOAA (National Oceanic and Atmospheric Administration). Multivariate ENSO Index Version 2 (MEI.v2). Available online: https://www.esrl.noaa.gov/psd/enso/mei/ (accessed on 20 June 2019).
- R Core Team. A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2017. [Google Scholar]
- Tanaka, N.; Kume, T.; Yoshifuji, N.; Tanaka, K.; Takizawa, H.; Shiraki, K.; Suzuki, M. A Review of Evapotranspiration Estimates from Tropical Forests in Thailand and Adjacent Regions. Agric. For. Meteorol. 2008, 148, 807–819. [Google Scholar] [CrossRef]
- Kirtphaiboon, S.; Wongwises, P.; Limsakul, A.; Sooktawee, S.; Humphries, U. Rainfall Variability over Thailand Related to the El Nino-Southern Oscillation ENSO). J. Sustain. Energy Environ. 2014, 5, 1–6. [Google Scholar]
- NOAA (National Oceanic and Atmospheric Administration). Cold and Warm Episodes by Season. Available online: https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.php (accessed on 30 December 2018).
- Trisurat, Y.; Alkemade, R.; Arets, E. Projecting Forest Tree Distributions and Adaptation to Climate Change in Northern Thailand. J. Ecol. Nat. Environ. 2009, 1, 55–63. [Google Scholar]
- Elliott, S.; Baker, P.J.; Borchert, R. Leaf Flushing during the Dry season: The Paradox of Asian Monsoon Forests. Glob. Ecol. Biogeogr. 2006, 15, 248–257. [Google Scholar] [CrossRef]
- Limsakul, A.; Singhruck, P. Long-Term Trends and Variability of Total and Extreme Precipitation in Thailand. Atmos. Res. 2016, 169, 301–317. [Google Scholar] [CrossRef]
- Cleland, E.E.; Allen, J.M.; Crimmins, T.M.; Dunne, J.A.; Pau, S.; Travers, S.E.; Wolkovich, E.M. Phenological tracking enables positive species responses to climate change. Ecology 2012, 93, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Ishida, A.; Yamazaki, J.Y.; Harayama, H.; Yazaki, K.; Ladpala, P.; Nakano, T.; Maeda, T. Photoprotection of Evergreen and Drought-Deciduous Tree Leaves to Overcome the Dry Season in Monsoonal Tropical Dry Forests in Thailand. Tree Physiol. 2013, 43, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Ishida, A.; Diloksumpun, S.; Ladpala, P.; Staporn, D.; Panuthai, S.; Gamo, M.; Puangchit, L. Contrasting Seasonal Leaf Habits of Canopy Trees between Tropical Dry-Deciduous and Evergreen Forests in Thailand. Tree Physiol. 2006, 26, 643–656. [Google Scholar] [CrossRef] [PubMed]
- Milla, R.; Castro-Díez, P.; Maestro-Martínez, M.; Montserrat-Martí, G. Relationships between Phenology and the Remobilization of Nitrogen, Phosphorus and Potassium in Branches of Eight Mediterranean Evergreens. New Phytol. 2005, 168, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Estiarte, M.; Penuelas, J. Alteration of the Phenology of Leaf Senescence and Fall in Winter Deciduous Species by Climate Change: Effects on Nutrient Proficiency. Glob. Chang. Biol. 2015, 21, 1005–1017. [Google Scholar] [CrossRef]
- Gautam, T.P.; Mandal, T.N. Storage and Flux of Nutrients in Disturbed and Undisturbed Tropical Moist Forest of Eastern Nepal. Int. J. For. Res. 2018. [Google Scholar] [CrossRef]
- Lima, A.L.A.; Rodal, M.J.N. Phenology and Wood Density of Plants Growing in the Semi-arid Region of Northeastern Brazil. J. Arid Environ. 2010, 74, 1363–1373. [Google Scholar] [CrossRef]
- Méndez-Alonzo, R.; Paz, H.; Zuluaga, R.C.; Rosell, J.A.; Olson, M.E. Coordinated Evolution of Leaf and Stem Economics in Tropical Dry Forest Trees. Ecology 2012, 93, 2397–2406. [Google Scholar] [CrossRef]
- Graefe, S.; Fang, D.; Butz, P. Water Residence Times in Trees of a Neotropical Dry Forest. Trees 2019, 33, 1225–1231. [Google Scholar] [CrossRef]
- Rivera, G.; Elliott, S.; Caldas, L.S.; Nicolossi, G.; Coradin, V.T.R.; Borchert, R. Increasing Day-Length Induces Spring Flushing of Tropical Dry Forest Trees in the Absence of Rain. Trees 2002, 16, 445–456. [Google Scholar] [CrossRef]
- Hiromi, T.; Ichie, T.; Kenzo, T.; Ninomiya, I. Interspecific Variation in Leaf Water Use Associated with Drought Tolerance in Four Emergent Dipterocarp Species of a Tropical Rain Forest in Borneo. J. For. Res. 2012, 17, 369–377. [Google Scholar] [CrossRef]
- Condit, R.; Aguilar, S.; Hernandez, A.; Perez, R.; Lao, S.; Angehr, G.; Foster, R.B. Tropical Forest Dynamics across a Rainfall Gradient and the Impact of an El Niño Dry Season. J. Trop. Ecol. 2004, 20, 51–72. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Leaf Phenology | Clustering of Mature Leaf Stage | Wood Density (g cm−3) |
|---|---|---|---|---|
| Shorea obtusa | Dipterocapaceae | Deciduous | C1 | 0.77 ± 0.06 b,c |
| Shorea roxburghii | Dipterocapaceae | Deciduous | C1 | 0.68 ± 0.05 d |
| Shorea siamensis | Dipterocapaceae | Deciduous | C2 | 0.72 ± 0.05 c,d |
| Sindora siamensis | Leguminosae | Brevi-deciduous | C2 | 0.81 ± 0.03 b |
| Lannea coromandelica | Anacardiaceae | Stem succulent | C3 | 0.54 ± 0.02 e |
| Erythrophleum succirubrum. | Leguminosae-Caesalpinioideae | Brevi-deciduous | C4 | 0.90 ± 0.03 a |
| Dipterocarpus obtusifolius | Dipterocapaceae | Semi-deciduous * | C5 | 0.74 ± 0.05 b,c,d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaewthongrach, R.; Vitasse, Y.; Lamjiak, T.; Chidthaisong, A. Impact of Severe Drought during the Strong 2015/2016 El Nino on the Phenology and Survival of Secondary Dry Dipterocarp Species in Western Thailand. Forests 2019, 10, 967. https://doi.org/10.3390/f10110967
Kaewthongrach R, Vitasse Y, Lamjiak T, Chidthaisong A. Impact of Severe Drought during the Strong 2015/2016 El Nino on the Phenology and Survival of Secondary Dry Dipterocarp Species in Western Thailand. Forests. 2019; 10(11):967. https://doi.org/10.3390/f10110967
Chicago/Turabian StyleKaewthongrach, Rungnapa, Yann Vitasse, Taninnuch Lamjiak, and Amnat Chidthaisong. 2019. "Impact of Severe Drought during the Strong 2015/2016 El Nino on the Phenology and Survival of Secondary Dry Dipterocarp Species in Western Thailand" Forests 10, no. 11: 967. https://doi.org/10.3390/f10110967
APA StyleKaewthongrach, R., Vitasse, Y., Lamjiak, T., & Chidthaisong, A. (2019). Impact of Severe Drought during the Strong 2015/2016 El Nino on the Phenology and Survival of Secondary Dry Dipterocarp Species in Western Thailand. Forests, 10(11), 967. https://doi.org/10.3390/f10110967