Climate Associations with Headwater Streamflow in Managed Forests over 16 Years and Projections of Future Dry Headwater Stream Channels

Abstract
:1. Introduction
2. Methods
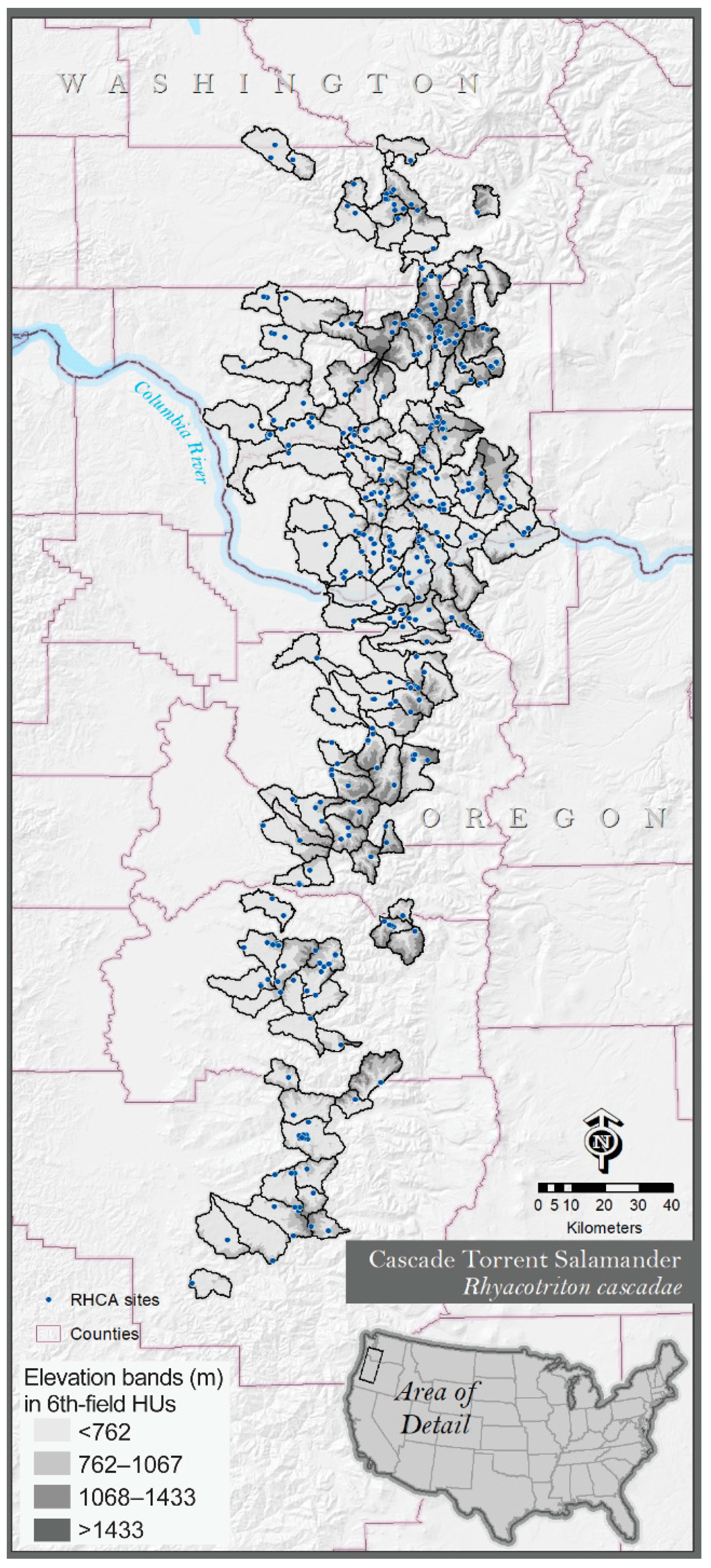
2.1. Study Area
2.2. Experimental Design
2.3. Field Methods
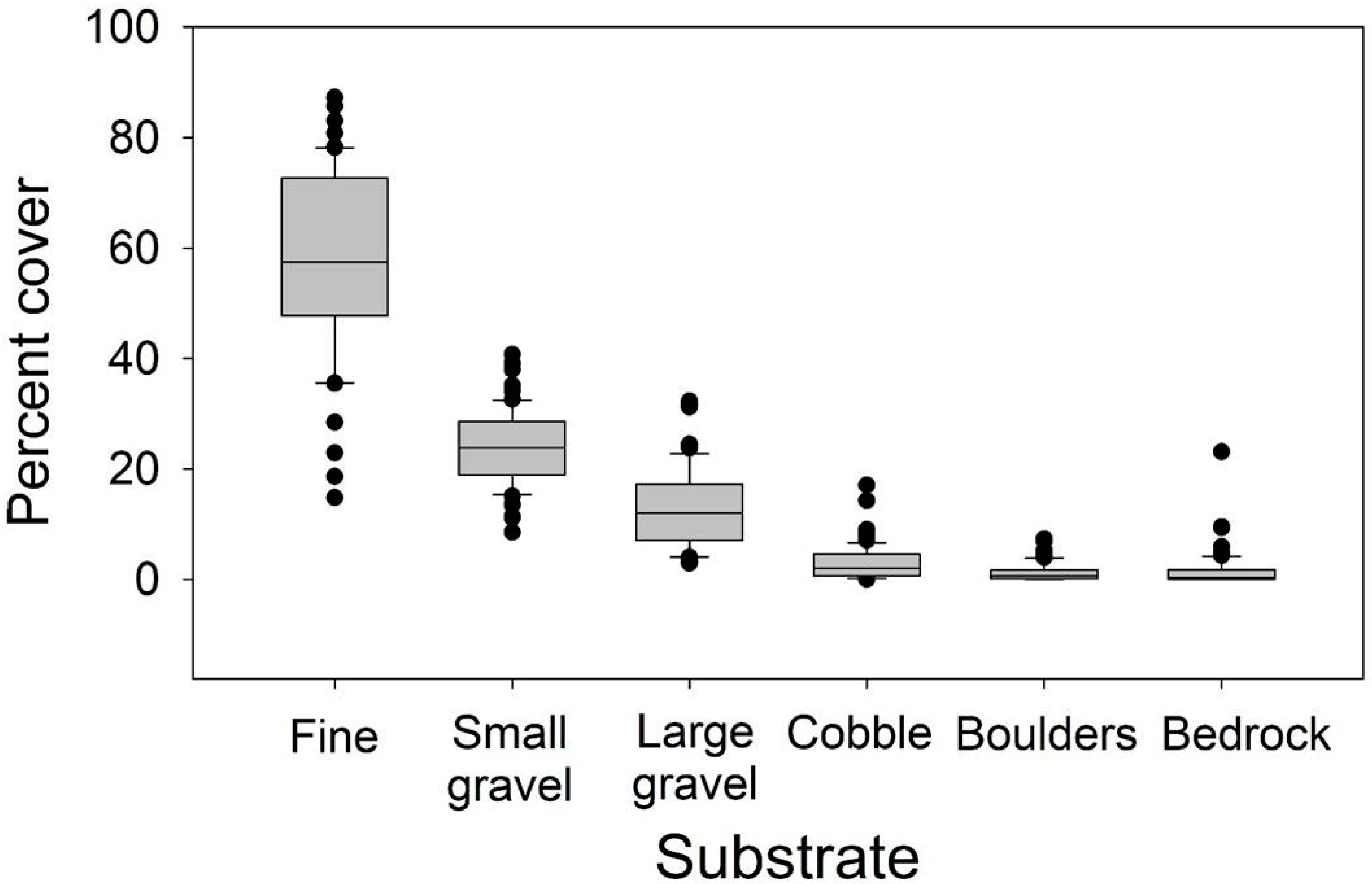
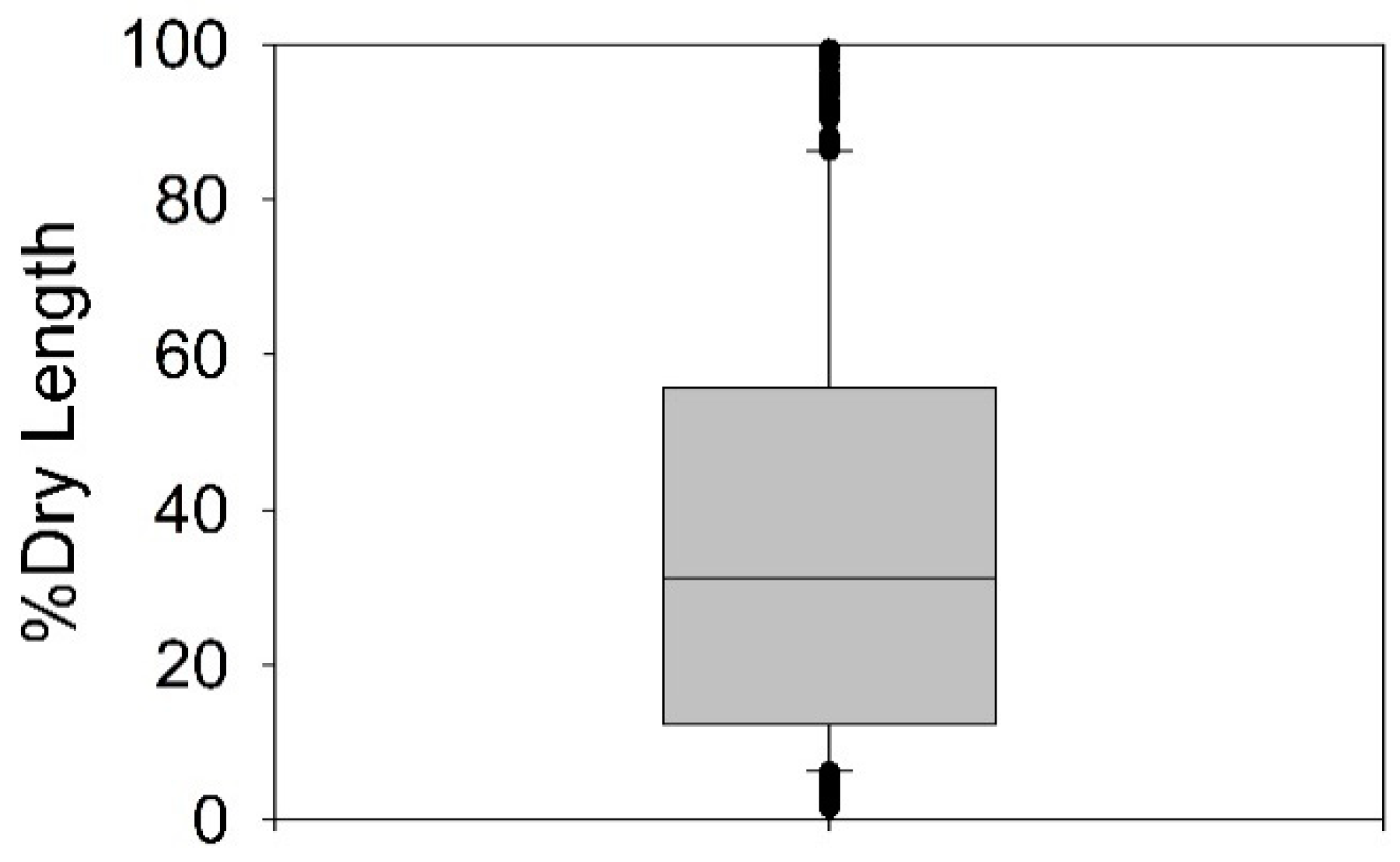
2.4. Stream Reach Characteristics
2.5. Climate Metrics and Future Scenarios
2.6. Statistical Analysis
2.7. Climate Change Projections
2.8. Landscape Projections
3. Results
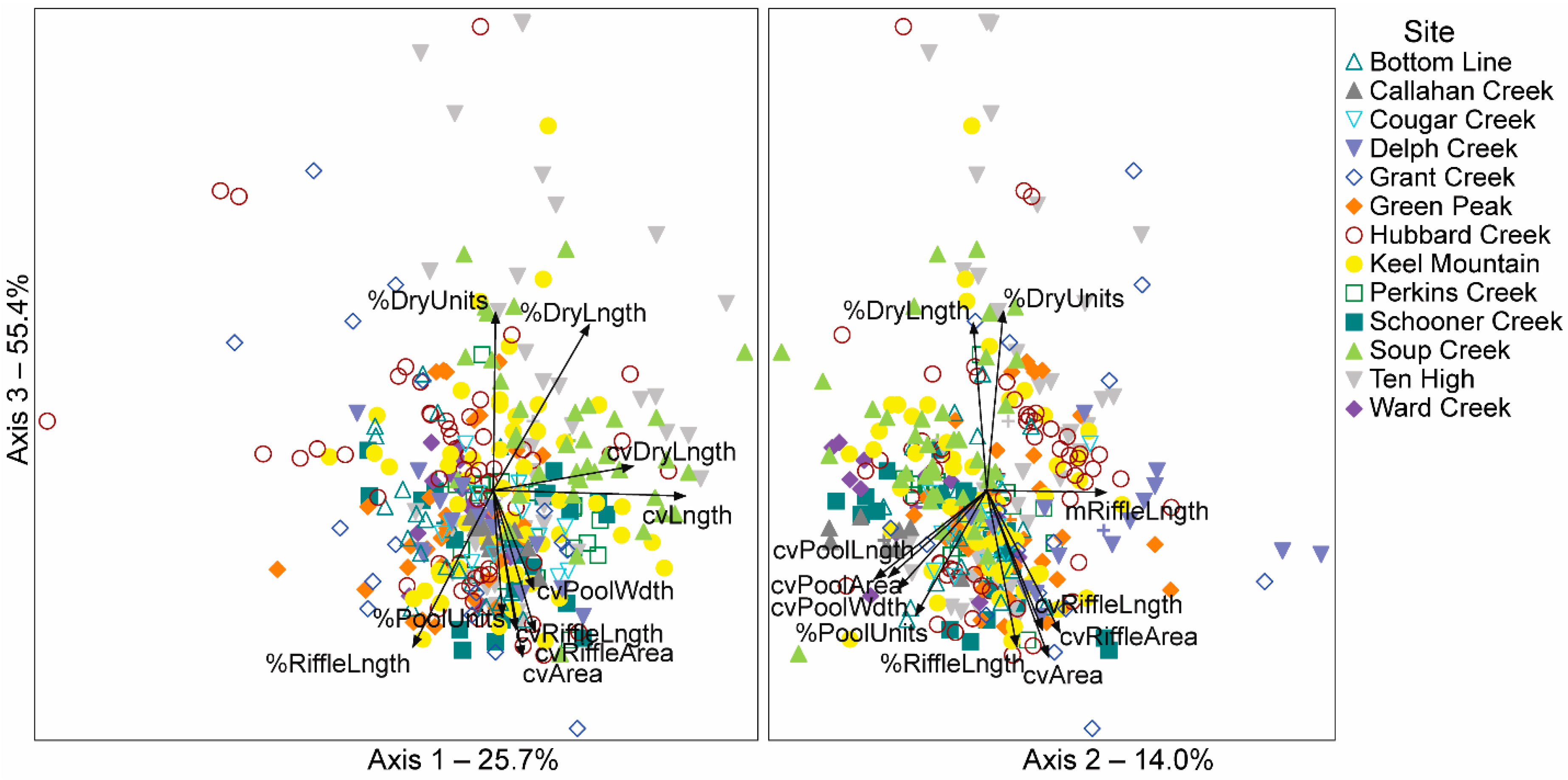
3.1. NMS Ordination
3.2. Relationships between Streamflow, Climate, Basin Area, and Buffer Treatments
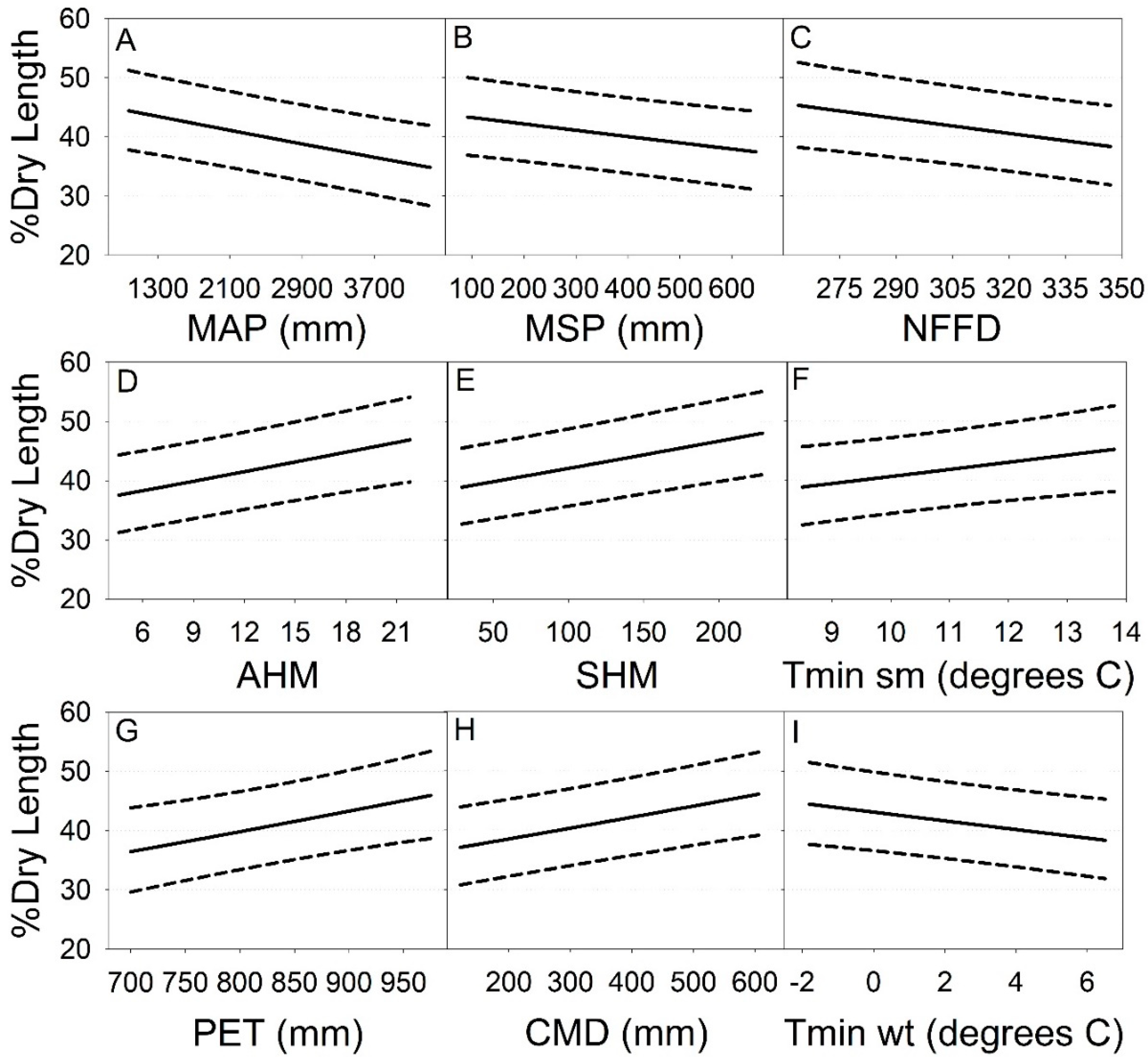
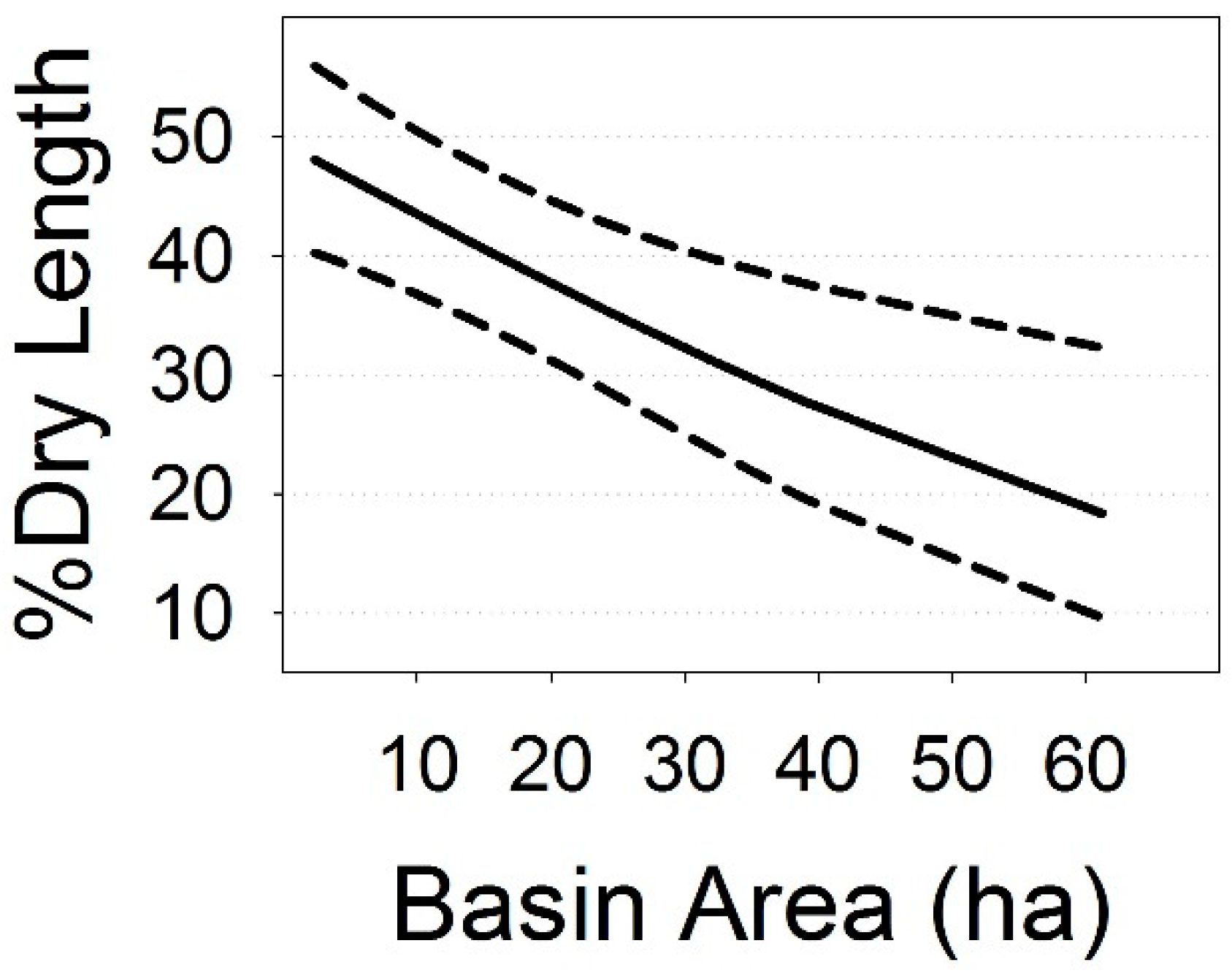
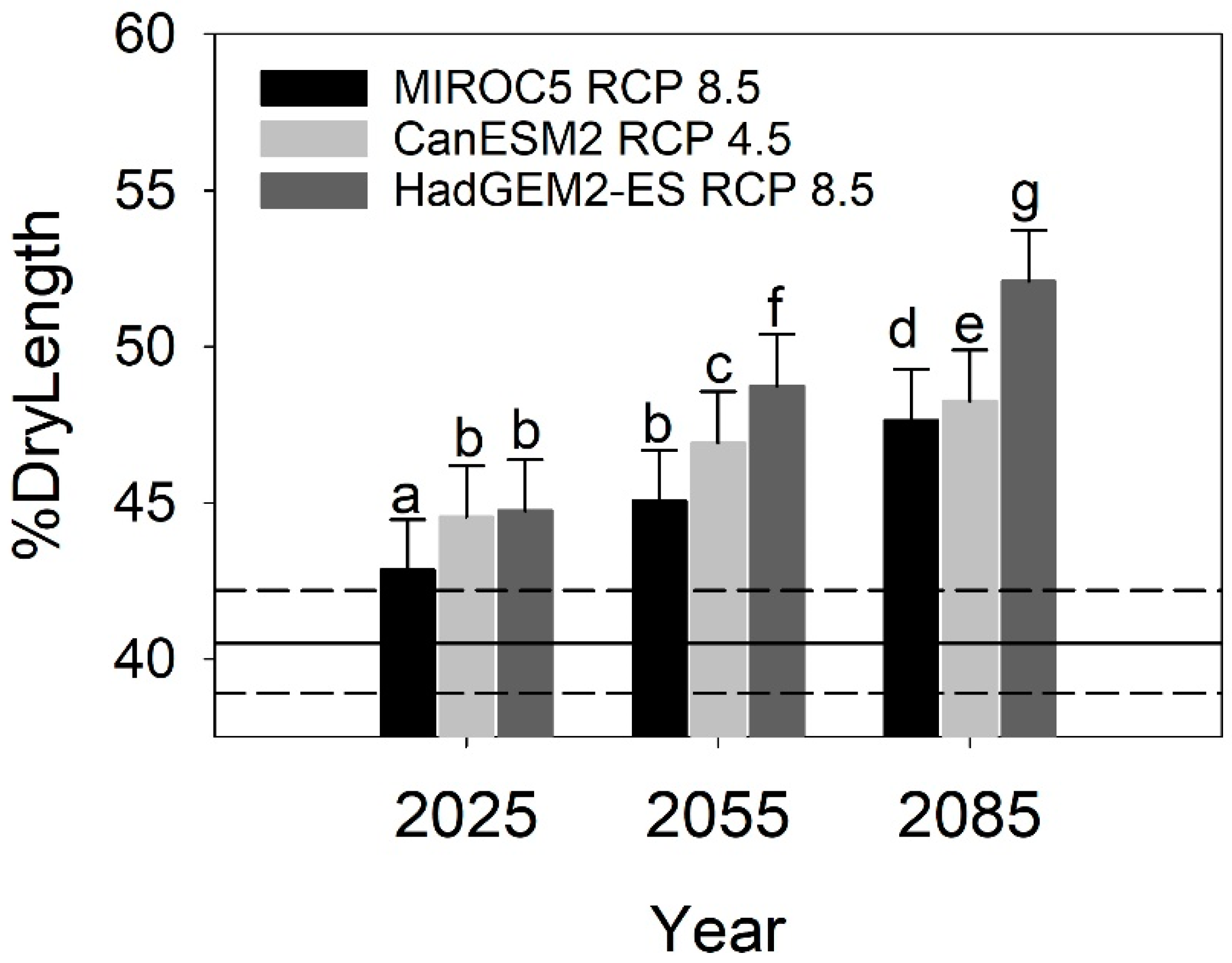
3.3. Climate Change Projections
3.4. Landscape Projections
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lindenmayer, D.B.; Franklin, J.F. Managing Forest Biodiversity: A Comprehensive Multiscaled Approach; Island Press: Washington, DC, USA, 2002; ISBN 1-55963-935-0. [Google Scholar]
- Olson, D.H.; van Horne, B. (Eds.) People, Forests and Change: Lessons from the Pacific Northwest; Island Press: Washington, DC, USA, 2017; ISBN 978-1-61091-766-7. [Google Scholar]
- Franklin, J.F.; Johnson, K.N.; Johnson, D.L. Ecological Forest Management; Waveland Press, Inc.: Long Grove, IL US, 2018. [Google Scholar]
- Franklin, J.F.; Spies, T.A.; Swanson, F.J. Setting the stage: Vegetation ecology and dynamics. In People, Forests and Change: Lessons from the Pacific Northwest; Olson, D.H., van Horne, B., Eds.; Island Press: Washington, DC, USA, 2017; pp. 16–32. [Google Scholar]
- Kim, J.B.; Marcot, B.G.; Olson, D.H.; Van Horne, B.; Vano, J.A.; Hand, M.S.; Salas, L.A.; Case, M.J.; Hennon, P.E.; D’Amore, D.V. Climate-smart approaches to managing forests. In People, Forests and Change: Lessons from the Pacific Northwest; Olson, D.H., van Horne, B., Eds.; Island Press: Washington, DC, USA, 2017; pp. 225–242. [Google Scholar]
- Reilly, M.J.; Spies, T.A.; Littell, J.; Butz, R.; Kim, J.B. Climate, disturbance, and vulnerability to vegetation change in the Northwest Forest Plan Area. In Synthesis of Science to Inform Land Management within the Northwest Forest Plan Area; General Technical Report PNW-GTR-996; Spies, T.A., Stine, P.A., Gravenmier, R., Long, J.W., Reilly, M.J., Eds.; US Department of Agriculture: Portland, OR, USA, 2018; Volume 1, pp. 29–92. [Google Scholar]
- Federal Register. 2012 Planning Rule. US Department of Agriculture, Forest Service. Available online: https://www.fs.usda.gov/detail/planningrule/home/?cid=stelprdb5359471 (accessed on 10 July 2019).
- Penaluna, B.E.; Olson, D.H.; Flitcroft, R.L.; Weber, M.; Bellmore, J.R.; Wondzell, S.M.; Dunham, J.B.; Johnson, S.L.; Reeves, G.H. Aquatic biodiversity in forests: A weak link in ecosystem services resilience. Biodivers. Conserv. 2017, 26, 3125–3155. [Google Scholar] [CrossRef]
- Reeves, G.H.; Spies, T.A. Watersheds and landscapes. In People, Forests and Change: Lessons from the Pacific Northwest; Olson, D.H., van Horne, B., Eds.; Island Press: Washington, DC, USA, 2017; pp. 207–222. [Google Scholar]
- Spies, T.A.; Hessburg, P.F.; Skinner, C.N.; Puettmann, K.J.; Reilly, M.J.; Davis, R.J.; Kertis, J.A.; Long, J.W.; Shaw, D.C. Old growth, disturbance, forest succession, and management in the area of the Northwest Fore Plan. In Synthesis of Science to Inform Land Management within the Northwest Forest Plan Area; Technical Coordinators; General Technical Report PNW-GTR-996; Spies, T.A., Stine, P.A., Gravenmier, R., Long, J.W., Reilly, M.J., Eds.; US Department of Agriculture: Portland, OR, USA, 2018; Volume 1, pp. 95–243. [Google Scholar]
- Molina, R.; Marcot, B.G.; Lesher, R. Protecting rare, old-growth, forest-associated species under the Survey and Manage program guidelines of the Northwest Forest Plan. Conserv. Biol. 2006, 20, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.H.; Penaluna, B.E.; Marcot, B.G.; Raphael, M.G.; Aubry, K.B. Biodiversity. In People, Forests and Change: Lessons from the Pacific Northwest; Olson, D.H., van Horne, B., Eds.; Island Press: Washington, DC, USA, 2017; pp. 174–190. [Google Scholar]
- Marcot, B.G.; Pope, K.L.; Slauson, K.; Welsh, H.H.; Wheeler, C.A.; Reilly, M.J.; Zielinski, W.J. Other species and biodiversity of older forests. In Synthesis of Science to Inform Land Management within the Northwest Forest Plan Area; Technical Coordinators; General Technical Report PNW-GTR-996; Spies, T.A., Stine, P.A., Gravenmier, R., Long, J.W., Reilly, M.J., Eds.; US Department of Agriculture: Portland, OR, USA, 2018; Volume 2, pp. 371–459. [Google Scholar]
- Olson, D.H.; Anderson, P.D.; Frissell, C.A.; Welsh, H.H., Jr.; Bradford, D.F. Biodiversity management approaches for stream riparian areas: Perspectives for Pacific Northwest headwater forests, microclimate and amphibians. For. Ecol. Manag. 2007, 246, 81–107. [Google Scholar] [CrossRef]
- Olson, D.H.; Burnett, K.M. Design and management of linkage areas across headwater drainages to conserve biodiversity in forest ecosystems. For. Ecol. Manag. 2009, 259S, S117–S126. [Google Scholar] [CrossRef]
- Olson, D.H.; Burnett, K.M. Geometry of forest landscape connectivity: Pathways for persistence. In Density Management for the 21st Century: West Side Story; General Technical Report PNW-GTR-880; Anderson, P.D., Ronnenberg, K.L., Eds.; US Department of Agriculture: Portland, OR, USA, 2013; pp. 220–238. [Google Scholar]
- Sidle, R.C.; Tsuboyama, Y.; Noguchi, S.; Hosoda, I.; Fujieda, M.; Shimizu, T. Streamflow generation in steep headwaters: A linked hydro-geomorphic paradigm. Hydrol. Process. 2000, 14, 369–385. [Google Scholar] [CrossRef]
- Meyer, J.L.; Wallace, J.B. Lost linkages and lotic ecology: Rediscovering small streams. In Ecology: Achievement and Challenge; Press, M.C., Huntly, N.J., Levin, S., Eds.; Blackwell Scientific: Oxford, UK, 2001; pp. 295–317. [Google Scholar]
- Gomi, T.; Sidle, R.C.; Richardson, J.S. Understanding processes and downstream linkages of headwater streams. BioScience 2002, 52, 905–916. [Google Scholar] [CrossRef]
- Datry, T.; Larned, S.T.; Tockner, K. Intermittent rivers: A challenge for freshwater ecology. BioScience 2014, 64, 229–235. [Google Scholar] [CrossRef]
- USA Department of Agriculture and Department of Interior. Record of Decision on Management of Habitat for Late-Successional and Old-Growth Forest Related Species within the Range of the Northern Spotted Owl; Northwest Forest Plan; US Department of Agriculture and Department of Interior: Portland, OR, USA, 1994.
- Pabst, R.J.; Spies, T.A. Distribution of herbs and shrubs in relation to landform and canopy cover in riparian forests of coastal Oregon. Can. J. Bot. 1998, 76, 298–315. [Google Scholar]
- Pabst, R.J.; Spies, T.A. Structure and composition of unmanaged riparian forests in the coastal mountains of Oregon, USA. Can. J. For. Res. 1999, 29, 1557–1573. [Google Scholar] [CrossRef]
- Progar, R.A.; Moldenke, A.R. Insect production from temporary and perennially flowing headwater streams in western Oregon. J. Freshw. Ecol. 2002, 17, 391–407. [Google Scholar] [CrossRef]
- Sheridan, C.D.; Olson, D.H. Amphibian assemblages in zero-order basins in the Oregon Coast Range. Can. J. For. Res. 2003, 33, 1452–1477. [Google Scholar] [CrossRef]
- Benda, L.; Poff, N.L.; Miller, D.; Dunne, T.; Reeves, G.; Pollock, M.; Pess, G. The network dynamics hypothesis: How river networks structure riverine habitats. BioScience 2004, 54, 413–427. [Google Scholar] [CrossRef]
- Ebersole, J.L.; Wigington, P.J., Jr.; Baker, J.P.; Cairns, M.A.; Church, M.R.; Compton, J.E.; Leibowitz, S.G.; Miller, B.; Hansen, B. Juvenile coho salmon growth and survival across stream network seasonal habitats. Trans. Am. Fish. Soc. 2006, 135, 1681–1697. [Google Scholar] [CrossRef]
- Ebersole, J.L.; Colvin, M.E.; Wigington, P.J., Jr.; Leibowitz, S.G.; Baker, J.P.; Church, M.R.; Compton, J.E.; Cairns, M.A. Hierarchical modeling of late-summer weight and summer abundance of juvenile coho salmon across a stream network. Trans. Am. Fish. Soc. 2009, 138, 1138–1156. [Google Scholar] [CrossRef]
- Hance, D.J.; Ganio, L.M.; Burnett, K.M.; Ebersole, J.L. Basin-scale variation in the spatial pattern of fall movement of juvenile Coho salmon in the West Fork Smith River, Oregon. Trans. Am. Fish. Soc. 2016, 145, 1018–1034. [Google Scholar] [CrossRef]
- Sheridan, C.D.; Spies, T.A. Vegetation-environment relationships in zero-order basins in coastal Oregon. Can. J. For. Res. 2005, 35, 340–355. [Google Scholar] [CrossRef]
- Danehy, R.J.; Bilby, R.E.; Langshaw, R.B.; Evans, D.M.; Turner, T.R.; Floyd, W.C.; Schoenholtz, S.H.; Duke, S.D. Biological and water quality responses to hydrologic disturbances in third-order forested streams. Ecohydrology 2012, 5, 90–98. [Google Scholar] [CrossRef]
- Reeves, G.H.; Burnett, K.M.; McGarry, E.V. Sources of large wood in a pristine watershed in coastal Oregon. Can. J. For. Res. 2003, 33, 1363–1370. [Google Scholar] [CrossRef]
- May, C.L.; Gresswell, R.E. Large wood recruitment and redistribution in headwater streams of the Oregon Coast Range, USA. Can. J. For. Res. 2003, 33, 1352–1362. [Google Scholar] [CrossRef]
- May, C.L.; Gresswell, R.E. Processes and rates of sediment and wood accumulation in headwater streams of the Oregon Coast Range, USA. Earth Surf. Process. Landf. 2004, 28, 409–424. [Google Scholar] [CrossRef]
- Benda, L.E.; Cundy, T.W. Predicting deposition of debris flows in mountain channels. Can. Geotech. J. 1990, 27, 409–417. [Google Scholar] [CrossRef]
- Benda, L.E.; Dunne, T. Stochastic forcing of sediment supply to channel networks from landsliding and debris flows. Water Resour. Res. 1997, 33, 2849–2863. [Google Scholar] [CrossRef]
- Benda, L.E.; Dunne, T. Stochastic forcing of sediment routing and storage in channel networks. Water Resour. Res. 1997, 33, 2865–2880. [Google Scholar] [CrossRef] [Green Version]
- Ebersole, J.L.; Wigington, P.J., Jr.; Leibowitz, S.G.; Comelio, R.L.; van Sickle, J. Predicting the occurrence of cold-water patches at intermittent and ephemeral tributary confluences with warm rivers. Freshw. Sci. 2015, 34, 111–124. [Google Scholar] [CrossRef]
- Reeves, G.H.; Olson, D.H.; Wondzell, S.M.; Bisson, P.A.; Gordon, S.; Miller, S.A.; Long, J.W.; Furniss, M.J. The aquatic conservation strategy of the Northwest Forest Plan—A review of the relevant science after 23 years. In Synthesis of Science to Inform Land Management within the Northwest Forest Plan Area; General Technical Report PNW-GTR-996; Spies, T.A., Stine, P.A., Gravenmier, R., Long, J.W., Reilly, M.J., Eds.; US Department of Agriculture: Portland, OR, USA, 2018; Volume 1, pp. 461–624. [Google Scholar]
- Wipfli, M.S.; Gregovich, D.P. Export of invertebrates and detritus from fishless headwater streams in southeast Alaska: Implications for downstream salmonid production. Freshw. Ecol. 2002, 47, 957–969. [Google Scholar] [CrossRef]
- Olson, D.H.; Kluber, M.R. Plethodontid salamander distributions in managed forest headwaters in western Oregon. Herpetol. Conserv. Biol. 2014, 9, 76–96. [Google Scholar]
- Emel, S.L.; Olson, D.H.; Knowles, L.L.; Storfer, A. Comparative landscape genetics of two endemic torrent salamander species, Rhyacotriton kezeri and R. variegatus: Implications for forest management and species conservation. Conserv. Genet. 2019, 20, 801–815. [Google Scholar] [CrossRef]
- Santiago, J.M.; Muñoz-Mas, R.; Solana, J.; de Jalón, D.G.; Alonso, C.; Martínez-Capel, F.; Pórtoles, J.; Monjo, R.; Ribalaygua, J. Waning habitats due to climate change: Effects of streamflow and temperature changes at the rear edge of the distribution of a cold-water fish. Hydrol. Earth Syst. Sci. 2017, 21, 4073–4101. [Google Scholar] [CrossRef]
- Latta, G.; Temesgen, H.; Adams, D.; Barrett, T. Analysis of potential impacts of climate change on forests of the United States Pacific Northwest. For. Ecol. Manag. 2010, 259, 720–729. [Google Scholar] [CrossRef]
- Rupp, D.E.; Abatzoglou, J.T.; Hegewisch, K.C.; Mote, P.W. Evaluation of CMIP5 20th century climate simulations for the Pacific Northwest USA. J. Geophys. Res. Atmos. 2013, 118, 10884–10906. [Google Scholar] [CrossRef]
- Isaak, D.J.; Wollrab, S.; Horan, D.; Chandler, G. Climate change effects on stream and river temperatures across the northwest US from 1980–2009 and implications for salmonid fishes. Clim. Chang. 2012, 113, 499–524. [Google Scholar] [CrossRef]
- Isaak, D.J.; Young, M.K.; Luce, C.H.; Hostetler, S.W.; Wenger, S.J.; Peterson, E.E.; ver Hoef, J.M.; Groce, M.C.; Horan, D.L.; Nagel, D.E. Slow climate velocities of mountain streams portend their role as refugia for cold-water biodiversity. Proc. Natl. Acad. Sci. USA 2016, 113, 4374–4379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tohver, I.M.; Hamlet, A.F.; Lee, S.-Y. Impacts of 21st-century climate change on hydrologic extremes in the Pacific Northwest region of North America. J. Am. Water Resour. Assoc. 2014, 50, 1461–1476. [Google Scholar] [CrossRef]
- Luce, C.H.; Lopez-Burgos, V.; Holden, Z. Sensitivity of snowpack storage to precipitation and temperature using spatial and temporal analog models. Water Resour. Res. 2014, 50, 9447–9462. [Google Scholar] [CrossRef]
- Tague, C.; Grant, G.E. Ground water dynamics mediate low-flow response to global warming in snow-dominated alpine regions. Water Resour. Res. 2009, 45, W07421. [Google Scholar] [CrossRef]
- Safeeq, M.; Grant, G.E.; Lewis, S.L.; Tague, C. Coupling snowpack and groundwater dynamics to interpret historical streamflow trends in the western United States. Hydrol. Process. 2013, 27, 655–668. [Google Scholar] [CrossRef]
- Safeeq, M.; Grant, G.E.; Lewis, S.L.; Kramer, M.G.; Staab, B. A hydrogeologic framework for characterizing summer streamflow sensitivity to climate warming in the Pacific Northwest, USA. Hydrol. Earth Syst. Sci. 2014, 18, 3693–3710. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, A.J.; Nolin, A.; Lewis, S.; Tague, C. Hydrogeologic controls on streamflow sensitivity to climatic variability. Hydrol. Process. 2008, 22, 4371–4385. [Google Scholar] [CrossRef]
- Hibbert, A.R. Forest treatment effects on water yield. In Forest Hydrology: Proceedings of a National Science Foundation Advanced Science Seminar; Sopper, W.E., Lull, H.W., Eds.; Pergamon Press: New York, NY, USA, 1967; pp. 527–543. [Google Scholar]
- Rothacher, J. Regimes of streamflow and their modification by logging. In Proceedings of the Symposium: Forest Land Uses and Stream Environment, Corvallis, OR, USA, 12–21 October 1970; pp. 40–54. [Google Scholar]
- Harr, R.D. Effects of streamflow in the rain-dominated portion of the Pacific Northwest. In Proceedings of the Workshop on Scheduling Timber Harvest for Hydrologic Concerns, Portland, OR, USA, 27–29 November 1979. [Google Scholar]
- Troendle, C.A.; King, R.M. The effect of timber harvest on the Fool Creek watershed, 30 years later. Water Resour. Res. 1985, 21, 1915–1922. [Google Scholar] [CrossRef]
- Campbell, I.C.; Doeg, T.J. Impact of timber harvesting and production on streams: A review. Aust. J. Mar. Freshw. Resour. 1989, 40, 519–539. [Google Scholar] [CrossRef] [Green Version]
- Bosch, J.M.; Hewlett, J.D. A review of catchment experiments to determine the effect of vegetation changes on water yield and evapotranspiration. J. Hydrol. 1982, 55, 3–23. [Google Scholar] [CrossRef]
- Andréassian, V. Waters and forests: From historical controversy to scientific debate. J. Hydrol. 2004, 291, 1–27. [Google Scholar] [CrossRef]
- Keppler, E.T.; Ziemer, R.R. Logging effects on streamflow: Water yield and summer low flows at Caspar Creek in northwestern California. Water Resour. Res. 1990, 26, 1669–1679. [Google Scholar] [CrossRef]
- Troendle, C.A.; Olsen, W.K. Potential effects of timber harvest and water management on streamflow dynamics and sediment transport. In Sustainable Ecological Systems: Implementing an Ecological Approach to Land Management; Technical Coordinators; General Technical Report GTR-RM-247; Covington, W.W., DeBano, L.F., Eds.; US Department of Agriculture: Fort Collins, CO, USA, 1994; pp. 34–41. [Google Scholar]
- Keppler, E.T. The summer flow and water yield response to timber harvest. In Technical Coordinator, Proceedings of the Conference on Coastal Watersheds: The Caspar Creek Story, Ukiah, CA, USA, 6 May 1998; Ziemer, R.R., Ed.; US Department of Agriculture: Albany, CA USA, 1998; pp. 35–43. [Google Scholar]
- Surfleet, C.G.; Skaugset, A.E. The effect of timber harvest on summer low flows, Hinkle Creek, Oregon. West. J. Appl. For. 2013, 28, 13–21. [Google Scholar] [CrossRef]
- Perry, T.D.; Jones, J.A. Summer streamflow deficits from regenerating Douglas-fir forest in the Pacific Northwest, USA. Ecohydrology 2017, 10, e1790. [Google Scholar] [CrossRef]
- Robles, M.D.; Turner, D.S.; Haney, J.A. A century of changing flows: Forest management changed flow magnitudes and warming advanced the timing of flow in a southwestern US river. PLoS ONE 2017, 12, e0187875. [Google Scholar] [CrossRef]
- Dwire, K.A.; Mellmann-Brown, S.; Gurrieri, J.T. Potential effects of climate change on riparian areas, wetlands, and groundwater-dependent ecosystems in the Blue Mountains, Oregon, USA. Clim. Serv. 2017, 10, 44–52. [Google Scholar] [CrossRef]
- Olson, D.H.; Weaver, G. Vertebrate assemblages associated with headwater hydrology in western Oregon managed forests. For. Sci. 2007, 53, 343–355. [Google Scholar]
- Olson, D.H.; Burton, J.H. Near-term effects of repeated-thinning with riparian buffers on headwater stream vertebrates and habitats in Oregon, USA. Forests 2014, 5, 2703–2729. [Google Scholar] [CrossRef]
- Cissel, J.H.; Anderson, P.D.; Olson, D.; Puettmann, K.J.; Berryman, S.D.; Chan, S.; Thompson, C. BLM Density Management and Riparian Buffer Study: Establishment Report and Study Plan; Scientific Investigations Report 2006-5087; US Geologial Survey: Reston, VA, USA, 2006.
- Franklin, J.F.; Dyrness, C.T. Natural Vegetation of Oregon and Washington; Oregon State University Press: Corvallis, OR, USA, 1988. [Google Scholar]
- Daly, C.; Gibson, W.P.; Taylor, G.H.; Johnson, G.L.; Pasteris, P. A knowledge-based approach to the statistical mapping of climate. Clim. Res. 2002, 22, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Hamann, A.; Spittlehouse, D.L.; Aitken, S.N. Development of scale-free climate data for western Canada for use in resource management. Int. J. Climatol. 2006, 26, 383–397. [Google Scholar] [CrossRef]
- Wang, T.; Hamann, A.; Spittlehouse, D.L.; Murdock, T.Q. ClimateWNA—High-resolution spatial climate data for Western North America. J. Appl. Meteorol. Climatol. 2012, 51, 16–29. [Google Scholar] [CrossRef]
- Olson, D.H.; Leirness, J.B.; Cunningham, P.G.; Steel, E.A. Riparian buffers and forest thinning: Effects on headwater vertebrates 10 years after thinning. For. Ecol. Manag. 2014, 321, 81–93. [Google Scholar] [CrossRef]
- Burton, J.I.; Olson, D.H.; Puettmann, K.J. Effects of riparian buffer width on wood loading in headwater streams after repeated forest thinning. For. Ecol. Manag. 2016, 372, 247–257. [Google Scholar] [CrossRef]
- Olson, D.H.; Rugger, C. Preliminary study of the effects of headwater riparian reserves with upslope thinning on stream habitats and amphibians in western Oregon. For. Sci. 2007, 53, 331–342. [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC). Fifth Assessment Report. 2014. Available online: www.ipcc.ch (accessed on 14 July 2019).
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Minchin, P.R. An evaluation of the relative robustness of techniques for ecological ordination. Vegetatio 1987, 69, 89–107. [Google Scholar] [CrossRef]
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data; Version 4.0; MjM Software: Gleneden Beach, OR, USA, 1999. [Google Scholar]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D. SAS(R) System for Mixed Models; SAS Institute Inc.: Cary, NC, USA, 1996. [Google Scholar]
- Paterson, S.; Lello, J. Mixed models: Getting the best use of parasitological data. Trends Parasitol. 2003, 19, 370–375. [Google Scholar] [CrossRef]
- Akaike, H. Maximum likelihood identification of Gaussian autoregressive moving average models. Biometrika 1973, 60, 255–265. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Howell, B.A.; Maggiulli, N.M. Conservation Assessment for the Cascade Torrent Salamander (Rhyacotriton cascadae); Version 1.0; US Department of Agriculture: Portland, OR, USA, 2011. Available online: https://www.fs.fed.us/r6/sfpnw/issssp/species-index/fauna-amphibians.shtml (accessed on 31 October 2019).
- Rundio, D.E.; Olson, D.H. Palatability of southern torrent salamander (Rhyacotriton variegatus) larvae to Pacific giant salamander (Dicamptodon tenebrosus) larvae. J. Herpetol. 2001, 35, 133–136. [Google Scholar] [CrossRef]
- Rundio, D.E.; Olson, D.H. Antipredator defenses of larval Pacific giant salamanders (Dicamptodon tenebrosus) against cutthroat trout (Oncorhynchus clarkii). Copeia 2003, 2, 392–397. [Google Scholar]
- Welsh, H.H., Jr.; Lind, A.J. Habitat correlates of the southern torrent salamander, Rhyacotriton variegatus (Caudata: Rhyacotritonidae), in northwestern California. J. Herpetol. 1996, 30, 385–398. [Google Scholar] [CrossRef]
- Leach, J.A.; Olson, D.H.; Anderson, P.D.; Eskelson, B.N.I. Spatial and seasonal variability of forested headwater stream temperatures in western Oregon, USA. Aquat. Sci. 2016, 79, 291–307. [Google Scholar] [CrossRef]
- Jones, L.L.C.; Leonard, W.P.; Olson, D.H. (Eds.) Amphibians of the Pacific Northwest; Seattle Audubon Society: Seattle, WA, USA, 2005. [Google Scholar]
- Vesely, D.G. Terrestrial Amphibian Abundance and Species Richness in Headwater Riparian Buffer Strips, Oregon Coast Range. Master’s Thesis, Oregon State University, Corvallis, OR USA, 1996. [Google Scholar]
- Gomez, D.M.; Anthony, R.G. Amphibian and reptile abundance in riparian and upslope areas of five forest types in western Oregon. Northwest Sci. 1996, 70, 109–119. [Google Scholar]
- Kluber, M.R.; Olson, D.H.; Puettmann, K.J. Thermal profiles of downed wood under different forest management regimes in the Oregon Coast Range and their potential impact on plethodontid salamander habitat. Northwest Sci. 2009, 83, 25–34. [Google Scholar] [CrossRef]
- Bormann, B.T.; Williams, B.K.; Minkova, T. Learning to learn: The best available science of adaptive management. In People, Forests and Change: Lessons from the Pacific Northwest; Olson, D.H., van Horne, B., Eds.; Island Press: Washington, DC, USA, 2017; pp. 102–115. [Google Scholar]
- Cissel, J.H.; Swanson, F.J.; Weisberg, P.J. Landscape management using historical fire regimes: Blue River, Oregon. Ecol. Appl. 1999, 9, 1217–1231. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | No. Streams | No. Reaches | Reach Treatments | Years Sampled | Post-Treatment Year Sampled |
|---|---|---|---|---|---|
| Bottom Line | 6 | 7 | 1, 2, 4 | 1998, ′99, 2002 | 1, 2 and 5 |
| Callahan Creek | 1 | 1 | 3 | 1996, ′98, ′99, 2002, ′08, ′09, ′10, ′11 | 0, 1, 2, 5 |
| Cougar Creek | 2 | 3 | 2, 3, 4 | 1996, ′99, 2000, ′03 | 0, 1, 2, 5 |
| Delph Creek | 2 | 2 | 2, 3 | 2001, ′02, ′05, ′08, ′09, ′10, ′11, one also in 1997 | 0, 1, 2, 5, 8, 9, 10, 11 |
| Grant Creek | 3 | 4 | 2, 3, 4 | 1996, ’99, 2000, ′03 | 0, 1, 2, 5 |
| Green Peak | 4 | 4 | 2, 3, 4 | 1998, ′99, 2000, ′01, ′04, ′08, ′09, ′10, ′11 | 0, 1, 2, 5, 9, 10, 11, 12 |
| O.M. Hubbard | 6 | 9 | 1, 2, 4 | Reaches 34 & 36: 1995, ′98, ′99, 2002, ′08, ′09, ′10, ′11; reaches 28, 29, 31 & 32: 1995, ′98, ′99, 2002 | 0, 1, 2, 5, 11, 12, 13, 14 |
| Keel Mountain | 11 | 12 | 1, 2, 3, 4 | Reaches 41, 44, 54 & 56: 1995, ′99, 2000; reaches 153, 154, 158, 159 & 160: 2008, ′09, ′10, ′11; reaches 49, 51, 53: 1995, ′99, 2000, ′03, ′08, ′09, ′10, ′11 | Reaches 49, 51, 54, 56: 0, 1, 2; reach 53: 0, 1, 2, 10, 11, 12, 13; reaches 41, 44, 54, 56: 0, 1, 2; other reaches 10–13 |
| Perkins Creek | 3 | 3 | 3, 4 | 1997, 2000, ′01, ′04, ′08, ′09, ′10, ′11 | 0, 1, 2, 5, 9, 10, 11, 12 |
| Schooner Creek (1 & 2) | 3 | 3 | 2, 3, 4 | 1997, ′03, ′04, ′05/’06 | 0, 3, 4, 5/6 |
| Schooner Creek (5 & 6) | 3 | 3 | 2, 3, 4 | 1997, ′01, ′02, ′05 | 0, 1, 2, 5 |
| North Soup | 5 | 5 | 1, 2, 3, 4 | 1996, ′99, 2000, ′03, ′08, ′09, ′10, ′11 | 0, 1, 2, 5, 10, 11, 12, 13 |
| Ten High | 6 | 6 | 2, 3, 4 | 1997, 2001, ′02, ′05, ′08, ′09, ′11 | 0, 1, 2, 5, 8, 9, 11 |
| North Ward | 2 | 3 | 2, 3, 4 | 1997, 2005, ′08, ′09, ′10, ′11 | 0, 2, 3, 4, 5 |
| Stream Reach and Flow Characteristics | Average | Range | |
|---|---|---|---|
| Mean ± SE | Minimum | Maximum | |
| General reach characteristics | |||
| Dry to Wet ratio (Dry: Wet) | 2.6 (0.6) | 0.01 | 187.4 |
| Average | |||
| Unit length (m) | 14.7 (0.8) | 2.6 | 153.9 |
| Unit width (m) | 0.4 (0.01) | 0.1 | 2.1 |
| Unit area (m2) | 9.7 (1.6) | 0.05 | 312.2 |
| Variability | |||
| CV Unit length (%) | 155.8 (2.5) | 65.6 | 359.8 |
| CV Unit Width (%) | 45.8 (0.9) | 0 | 130.9 |
| CV Unit Area (%) | 140.1 (2.5) | 0 | 367.0 |
| % Units by major flow class | |||
| Dry | 30.9 (0.7) | 2 | 66.7 |
| Riffle | 43.7 (0.4) | 0 | 66.7 |
| Pool | 24.3 (0.8) | 0 | 54.5 |
| % Reach length by major flow class | |||
| Dry | 36.2 (1.4) | 1.5 | 99.5 |
| Riffle | 55.9 (1.4) | 0 | 96.7 |
| Pool | 5.8 (0.4) | 0 | 35.8 |
| Surface flow characteristics | |||
| Average | |||
| Length of dry units (m) | 18.3 (1.7) | 1.4 | 306.5 |
| Length of riffle units (m) | 15.9 (0.9) | 0 | 96.7 |
| Length of pool units (m) | 1.3 (0.05) | 0 | 9.4 |
| Width of riffle units (m) | 0.4 (0.01) | 0 | 2.1 |
| Width of pool units (m) | 0.4 (0.01) | 0 | 1.9 |
| Area of riffle units (m2) | 11.2 (1.6) | 0 | 420 |
| Area of pool units (m2) | |||
| Variability | |||
| CV Dry unit length | 107.3 (2.8) | 0 | 280.4 |
| CV Riffle unit length | 100.3 (1.9) | 0 | 221.8 |
| CV Pool unit length | 32.7 (1.4) | 0 | 180.3 |
| CV Riffle unit width | 44.5 (1.1) | 0 | 142.6 |
| CV Pool unit width | 28.2 (1.1) | 0 | 114.3 |
| CV Riffle unit area | 115.5 (2.3) | 0 | 286 |
| CV Pool unit area | 47.6 (2.0) | 0 | 364.9 |
| Climate Metrics |
|---|
| Mean annual temperature (°C, MAT) |
| Mean warmest month temperature (°C, MWMT) |
| Mean coldest month temperature (°C, MCMT) |
| Temperature difference between MWMT and MCMT, or continentality (°C, TD) |
| Hargreaves potential evaporation (mm, PET) |
| Hargreaves reference climatic moisture deficit (mm, CMD) |
| Degree days below 0 °C, chilling degree days (DD < 0) |
| Degree days above 5 °C, growing degree days (DD > 5) |
| Degree days above 18 °C, heating degree days (DD > 18) |
| Degree days below 18 °C, chilling degree days (DD < 18) |
| Number of frost-free days |
| Precipitation as snow (August in previous year—July in current year, in mm, PAS) |
| Mean annual precipitation (mm, MAP) |
| Winter mean maximum temperature (°C, Tmax wt) |
| Spring mean maximum temperature (°C, Tmax sp) |
| Summer mean maximum temperature (°C, Tmax sm) |
| Winter mean minimum temperature (°C, Tmin wt) |
| Spring mean minimum temperature (°C, Tmin sp) |
| Summer mean minimum temperature (°C, Tmin sm) |
| Annual heat: Moisture index ((MAT + 10)/(MAP/1000)) |
| Summer heat:moisture index (MWMT/(Mean Summer Precipitation/1000)) |
| Stream Reach and Flow Characteristics | Axis | ||
|---|---|---|---|
| Axis 1 | Axis 2 | Axis 3 | |
| General reach characteristics | |||
| Dry: Wet ratio | 0.279 | −0.106 | 0.353 |
| Unit length (m) | −0.067 | 0.467 | 0.49 |
| Unit width (m) | −0.324 | 0.268 | −0.251 |
| Unit area (m2) | −0.299 | 0.528 | −0.007 |
| Variability | |||
| CV Unit length (%) | 0.739 | 0.068 | −0.13 |
| CV Unit Width (%) | 0.134 | 0.325 | −0.157 |
| CV Unit Area (%) | 0.289 | 0.42 | −0.688 |
| % Units by major flow class | |||
| Dry | 0.079 | 0.217 | 0.71 |
| Riffle | −0.361 | 0.411 | −0.268 |
| Pool | 0.162 | −0.449 | −0.597 |
| % Reach length by flow class | |||
| Dry | 0.522 | −0.197 | 0.686 |
| Riffle | −0.477 | 0.29 | −0.667 |
| Pool | 0.088 | −0.476 | −0.437 |
| Surface flow characteristics | |||
| Length of dry units (m) | 0.309 | 0.193 | 0.368 |
| Length of riffle units (m) | −0.535 | 0.582 | −0.081 |
| Length of pool units (m) | −0.002 | −0.249 | −0.356 |
| Width of riffle units (m) | −0.296 | 0.312 | −0.279 |
| Width of pool units (m) | −0.08 | −0.198 | −0.485 |
| Area of riffle units (m2) | −0.278 | 0.521 | −0.112 |
| Area of pool units (m2) | −0.068 | −0.133 | −0.191 |
| Variability | |||
| CV Dry unit length | 0.63 | −0.12 | 0.262 |
| CV Riffle unit length | 0.252 | 0.396 | −0.629 |
| CV Pool unit length | 0.326 | −0.528 | −0.496 |
| CV Riffle unit width | 0.175 | 0.335 | −0.315 |
| CV Pool unit width | 0.339 | −0.502 | −0.529 |
| CV Riffle unit area | 0.345 | 0.456 | −0.634 |
| CV Pool unit area | 0.332 | −0.565 | −0.503 |
| Variable | Estimate | SE | DF_Denom | F | p |
|---|---|---|---|---|---|
| MAT | −0.01134 | 0.02678 | 131 | 0.18 | 0.673 |
| MWMT | 0.03496 | 0.02047 | 215 | 2.92 | 0.089 |
| MCMT | −0.00817 | 0.01151 | 180 | 0.5 | 0.479 |
| TD | 0.01603 | 0.01041 | 249 | 2.37 | 0.125 |
| MAP | −0.00012 | 0.00003 | 128 | 13.24 | <0.001 |
| MSP | −0.00044 | 0.00016 | 101 | 8.16 | 0.005 |
| AHM | 0.02234 | 0.00594 | 109 | 14.15 | <0.001 |
| SHM | 0.00186 | 0.00036 | 57.5 | 26.57 | <0.001 |
| DD < 0 | 0.00082 | 0.00084 | 158 | 0.96 | 0.328 |
| DD > 5 | −0.00001 | 0.00009 | 132 | 0.02 | 0.881 |
| DD < 18 | 0.00004 | 0.00008 | 147 | 0.27 | 0.603 |
| DD > 18 | 0.00009 | 0.00038 | 136 | 0.06 | 0.813 |
| NFFD | −0.00344 | 0.00127 | 112 | 7.34 | 0.008 |
| PAS | −0.00164 | 0.00110 | 283 | 2.23 | 0.137 |
| Eref | 0.00143 | 0.00054 | 266 | 7.08 | 0.008 |
| CMD | 0.00077 | 0.00020 | 76.7 | 14.67 | <0.001 |
| Tmax wt | −0.02220 | 0.01445 | 75 | 2.36 | 0.129 |
| Tmax sp | 0.01336 | 0.01254 | 118 | 1.13 | 0.289 |
| Tmax sm | 0.01031 | 0.01668 | 200 | 0.38 | 0.537 |
| Tmin wt | −0.03033 | 0.01234 | 96.7 | 6.04 | 0.016 |
| Tmin sp | −0.01672 | 0.02089 | 161 | 0.64 | 0.425 |
| Tmin sm | 0.04943 | 0.02317 | 244 | 4.55 | 0.034 |
| Effect | Estimate | Standard Error | DF | t | Pr > |t| |
|---|---|---|---|---|---|
| Intercept | −0.812 | 0.209 | 147 | −2.80 | 0.0059 |
| Area (ha) | −0.024 | 0.007 | 38.7 | −3.26 | 0.0023 |
| SHM | 0.002 | <0.001 | 60.7 | 60.7 | <0.001 |
| Tmin sm | 0.058 | 0.022 | 250 | 2.63 | 0.0090 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olson, D.H.; Burton, J.I. Climate Associations with Headwater Streamflow in Managed Forests over 16 Years and Projections of Future Dry Headwater Stream Channels. Forests 2019, 10, 968. https://doi.org/10.3390/f10110968
Olson DH, Burton JI. Climate Associations with Headwater Streamflow in Managed Forests over 16 Years and Projections of Future Dry Headwater Stream Channels. Forests. 2019; 10(11):968. https://doi.org/10.3390/f10110968
Chicago/Turabian StyleOlson, Deanna H., and Julia I. Burton. 2019. "Climate Associations with Headwater Streamflow in Managed Forests over 16 Years and Projections of Future Dry Headwater Stream Channels" Forests 10, no. 11: 968. https://doi.org/10.3390/f10110968
APA StyleOlson, D. H., & Burton, J. I. (2019). Climate Associations with Headwater Streamflow in Managed Forests over 16 Years and Projections of Future Dry Headwater Stream Channels. Forests, 10(11), 968. https://doi.org/10.3390/f10110968