Seasonal Drought Effects on Intra-Annual Stem Growth of Taiwan Pine along an Elevational Gradient in Subtropical China


Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area and Tree Selection
2.2. Measurements of Stem Radial Variation
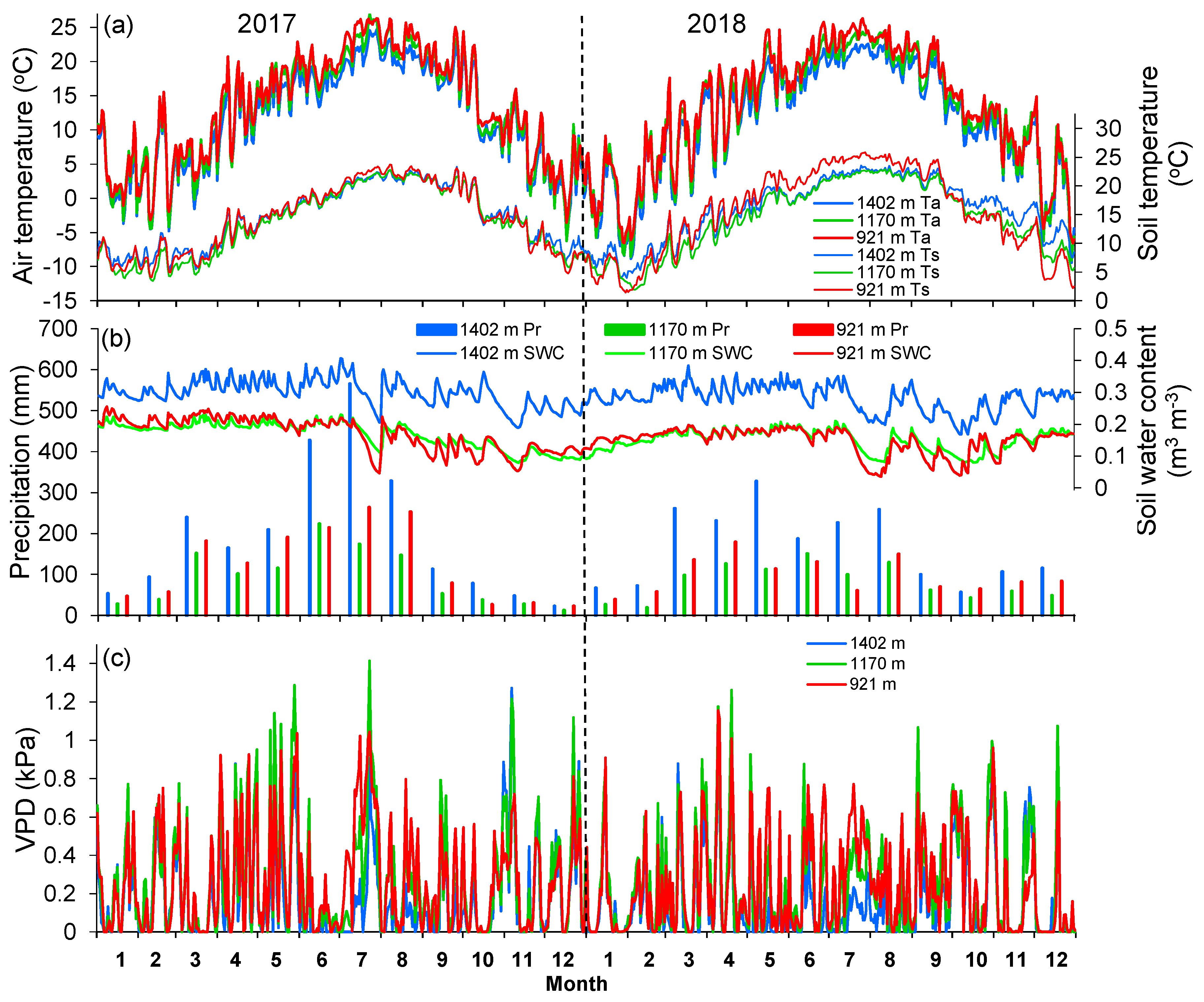
2.3. Microclimate Measurements
2.4. Modelling of Intra-Annual Stem Growth
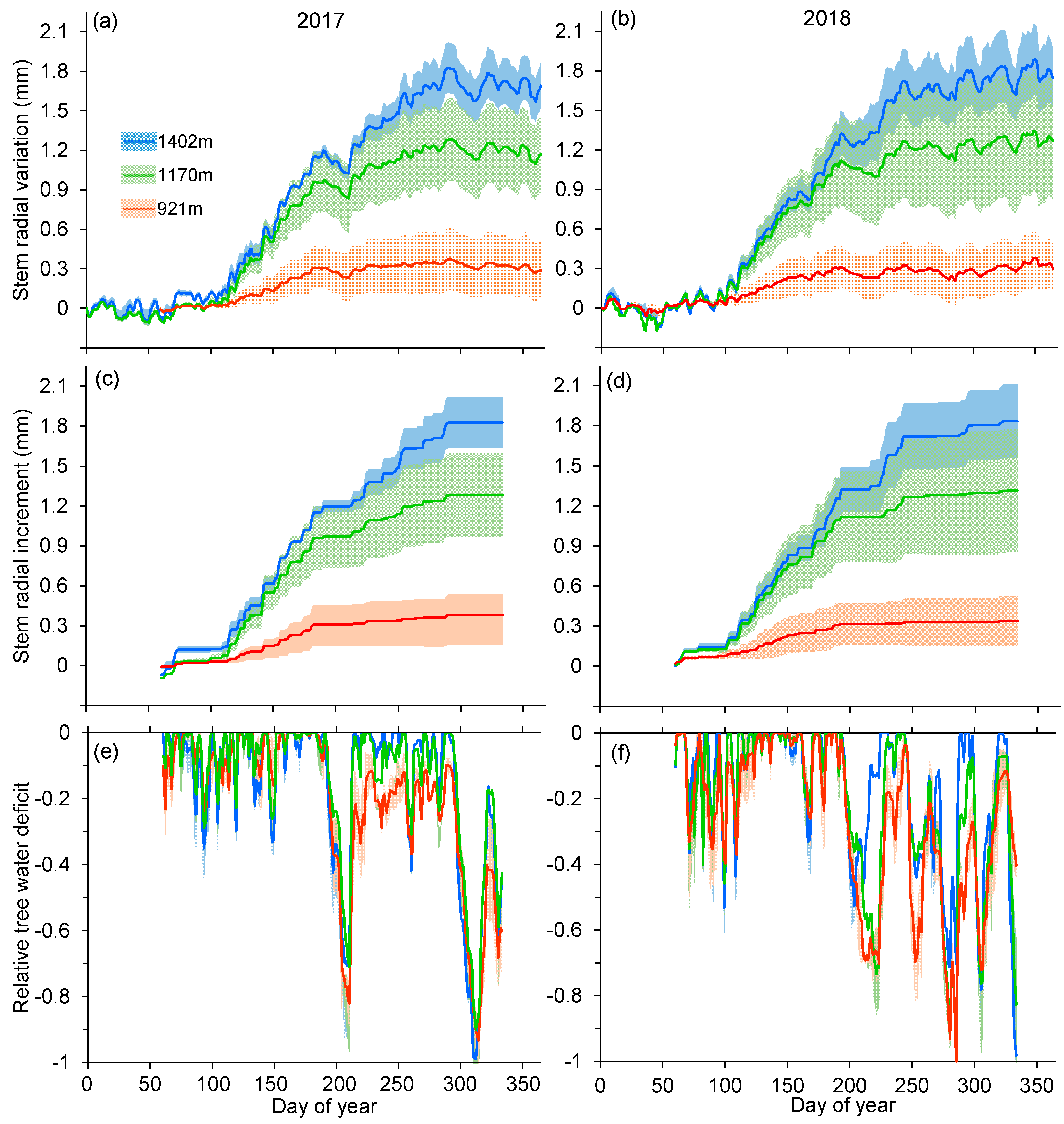
2.5. Extractions of Stem Radial Increment and Tree Water Deficit
2.6. Statistical Analysis
3. Results
3.1. Climate in 2017 and 2018
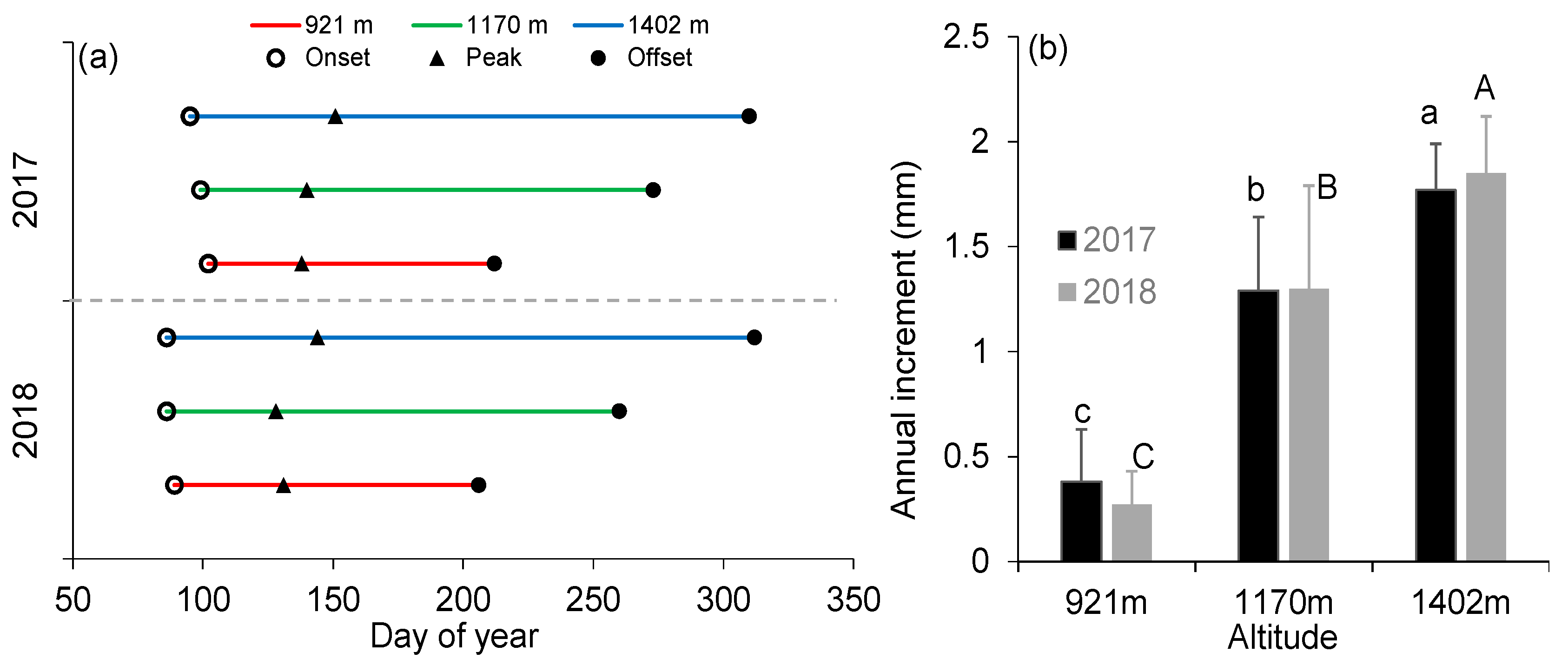
3.2. Elevational Variations in Intra-Annual Stem Growth Dynamics and Tree Water Deficit
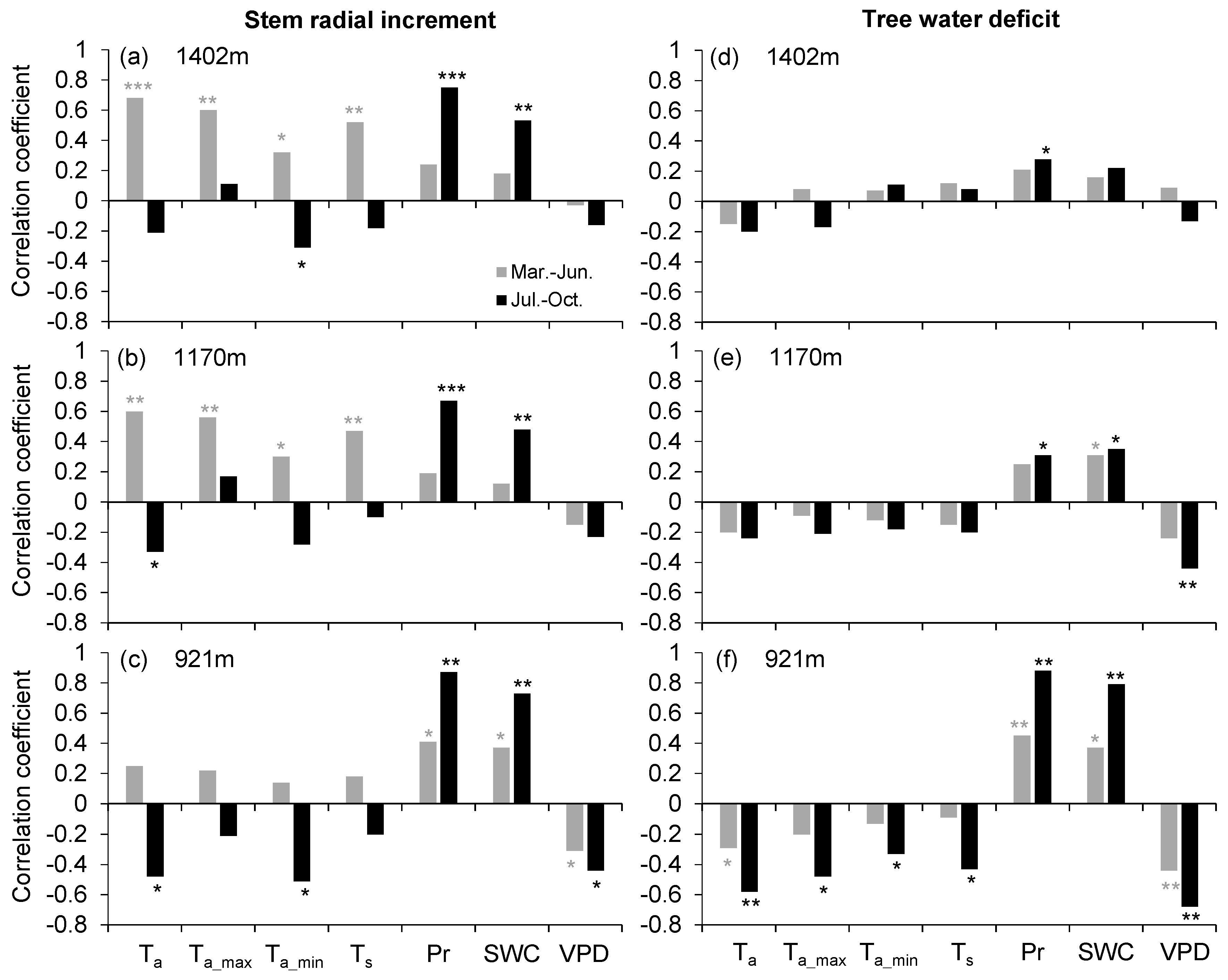
3.3. Correlations of Stem Radial Increment and Tree Water Deficit versus Microclimatic Variables
4. Discussion
4.1. Intra-Annual Stem Growth Dynamics along the Elevation Transect
4.2. Seasonal Drought Effects on Stem Radial Growth
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tee mortality reveals emerging climate change risks for forest. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Wei, X.; Wu, Y.; Liu, S.; Huang, Y.; Yan, J.; Zhang, D.; Zhang, Q.; Liu, J.; Meng, Z.; et al. Quantifying the hydrological responses to climate change in an intact forested small watershed in Southern China. Glob. Chang. Biol. 2011, 17, 3736–3746. [Google Scholar] [CrossRef]
- Shi, J.; Cook, E.R.; Lu, H.; Li, J.; Wright, W.E.; Li, S. Tree-ring based winter temperature reconstruction for the lower reaches of the Yangtze River in southeast China. Clim. Res. 2010, 41, 169–175. [Google Scholar] [CrossRef]
- Duan, J.; Zhang, Q.-B.; Lv, L.; Zhang, C. Regional-scale Winter-spring temperature variability and chilling damage dynamics over the past two centuries in southeastern China. Clim. Dyn. 2012, 39, 919–928. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, Y.; Shao, X. Temperature variability inferred from tree-ring widths in the Dabie Mountains of subtropical central China. Trees 2012, 26, 1887–1894. [Google Scholar] [CrossRef]
- Dang, H.; Zhang, Y.; Zhang, K.; Jiang, M.; Zhang, Q. Climate-growth relationships of subalpine fir (Abies fargesii) across the altitudinal range in the Shennongjia Mountains, central China. Clim. Chang. 2013, 117, 903–917. [Google Scholar] [CrossRef]
- Cai, Q.; Liu, Y. Two centuries temperature variations over subtropical southeast China inferred from Pinus taiwanensis Hayata tree-ring width. Clim. Dyn. 2017, 48, 1813–1825. [Google Scholar] [CrossRef]
- Cai, Q.; Liu, Y.; Duan, B.; Sun, C. Regional difference of the start time of the recent warming in Eastern China: Prompted by a 165-year temperature record deduced from tree rings in the Dabie Mountains. Clim. Dyn. 2018, 50, 2157–2168. [Google Scholar] [CrossRef]
- Chen, F.; Yuan, Y.J.; Wei, W.S.; Yu, S.L.; Zhang, T.W. Reconstructed temperature for Yong’an, Fujian, Southeast China: Linkages to the Pacific Ocean climate variability. Glob. Planet. Chang. 2012, 86, 11–19. [Google Scholar] [CrossRef]
- Li, Y.; Fang, K.; Cao, C.; Li, D.; Zhou, F.; Dong, Z.; Zhang, Y.; Gan, Z. A tree-ring chronology spanning 210 years in the coastal area of southeastern China, and its relationship with climate change. Clim. Res. 2016, 67, 209–220. [Google Scholar] [CrossRef]
- Cai, Q.; Liu, Y.; Liu, H.; Sun, C.; Wang, Y. Growing-season precipitation since 1872 in the coastal area of subtropical southeast China reconstructed from tree rings and its relationship with the East Asian summer monsoon system. Ecol. Indic. 2017, 82, 441–450. [Google Scholar] [CrossRef]
- Luo, D.; Huang, J.-G.; Jiang, X.; Ma, Q.; Liang, H.; Guo, X.; Zhang, S. Effect of climate and competition on radial growth of Pinus massoniana and Schima superba in China’s subtropical monsoon mixed forest. Dendrochronologia 2017, 46, 24–34. [Google Scholar] [CrossRef]
- Deslauriers, A.; Morin, H.; Urbinati, C.; Carrer, M. Daily weather response of balsam fir (Abies balsamea (L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Québec (Canada). Trees Struct. Funct. 2003, 17, 477–484. [Google Scholar] [CrossRef]
- Zweifel, R.; Haeni, M.; Buchmann, N.; Eugster, W. Are trees able to grow in periods of stem shrinkage? New Phytol. 2016, 211, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Mencuccini, M.; Salmon, Y.; Mitchell, P.; Hölttä, T.; Choat, B.; Meir, P.; O’Grady, A.; Tissue, D.; Zweifel, R.; Sevanto, S.; et al. An empirical method that separates irreversible stem radial growth from bark water content changes in trees: Theory and case studies. Plant Cell Environ. 2017, 40, 290–303. [Google Scholar] [CrossRef]
- Van der Maaten, E.; Pape, J.; Van der Maaten-Theunissen, M.; Scharnweber, T.; Smiljanić, M.; Cruz-García, R.; Wilmking, M. Distinct growth phenology but similar daily stem dynamics in three co-occurring broadleaved tree species. Tree Physiol. 2018, 38, 1820–1828. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Gričar, J.; Seo, J.-W.; Rathgeber, C.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- Camarero, J.J.; Olano, J.M.; Parras, A. Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol. 2010, 185, 471–480. [Google Scholar] [CrossRef]
- Dietrich, L.; Zweifel, R.; Kahmen, A. Daily stem diameter variations can predict the canopy water status of mature temperate trees. Tree Physiol. 2018, 38, 941–952. [Google Scholar] [CrossRef]
- Ziaco, E.; Biondi, F. Stem Circadian Phenology of Four Pine Species in Naturally Contrasting Climates from Sky-Island Forests of the Western USA. Forests 2018, 9, 396. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.S.; Nie, Y.Q.; Wen, F. Seasonal Dynamics of Stem Radial Increment of Pinus taiwanensis Hayata and Its Response to Environmental Factors in the Lushan Mountains, Southeastern China. Forests 2018, 9, 387. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.-G.; Guo, X.; Rossi, S.; Zhai, L.; Yu, B.; Zhang, S.; Zhang, M. Intra-annual wood formation of subtropical Chinese red pine shows better growth in dry season than wet season. Tree Physiol. 2018, 38, 1225–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Z.-X.; Bräuning, A.; Fu, P.-L.; Yang, R.-Q.; Qi, J.-H.; Grießinger, J.; Gebrekirstos, A. Intra-Annual Radial Growth of Pinus kesiya var. langbianensis Is Mainly Controlled by Moisture Availability in the Ailao Mountains, Southwestern China. Forests 2019, 10, 899. [Google Scholar] [CrossRef] [Green Version]
- Moser, L.; Fonti, P.; Büntgen, U.; Esper, J.; Luterbacher, J.; Franzen, J.; Frank, D. Timing and duration of European larch growing season along altitudinal gradients in the Swiss Alps. Tree Physiol. 2010, 30, 225–233. [Google Scholar] [CrossRef]
- Ziaco, E.; Biondi, F. Tree growth, cambial phenology, and wood anatomy of limber pine at a Great Basin (USA) mountain observatory. Trees Struct. Funct. 2016, 30, 1507–1521. [Google Scholar] [CrossRef] [Green Version]
- Saderi, S.; Rathgeber, C.B.K.; Rozenberg, P.; Fournier, M. Phenology of wood formation in larch (Larix decidua Mill.) trees growing along a 1000-m elevation gradient in the French Southern Alps. Ann. For. Sci. 2019, 76, 89. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, B.; Deslquriers, A.; Bräuning, A. Intra-annual stem radial increment response of Qilian juniper to temperature and precipitation along an altitudinal gradient in northwestern China. Trees Struct. Funct. 2015, 29, 25–34. [Google Scholar] [CrossRef]
- Zhang, J.; Gou, X.; Pederson, N.; Zhang, F.; Niu, H.; Zhao, S.; Wang, F. Cambial phenology in Juniperus przewalskii along different altitudinal gradients in a cold and arid region. Tree Physiol. 2018, 38, 840–852. [Google Scholar] [CrossRef]
- Gao, J.; Yang, B.; He, M.; Shishov, V. Intra-annual stem radial increment patterns of Chinese pine, Helan Mountains, Northern Central China. Trees Struct. Funct. 2019, 33, 751–763. [Google Scholar] [CrossRef]
- Ziaco, E.; Truettner, C.; Biondi, F.; Sarah, B. Moisture-driven xylogenesis in Pinus ponderosa from a Mojave Desert mountain reveals high phenological plasticity. Plant Cell Environ. 2018, 41, 823–836. [Google Scholar] [CrossRef]
- Ren, P.; Rossi, S.; Camarero, J.J.; Ellison, A.M.; Liang, E.; Peñuelas, J. Critical temperature and precipitation thresholds for the onset of xylogenesis of Juniperus przewalskii in a semi-arid area of the north-eastern Tibetan Plateau. Ann. Bot. 2018, 121, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Obojes, N.; Meurer, A.; Newsely, C.; Tasser, E.; Oberhuber, W.; Mayr, S.; Tappeiner, U. Water stress limits transpiration and growth of European larch up to the lower subalpine belt in an inner-alpine dry valley. New Phytol. 2018, 220, 460–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhuo, Z. On the Positional Problem of the Pinus Taiwanensis Forest in the Vegetation Altitude Belts of Mt. Lushan. Chin. J. Plan. Ecol. 1989, 13, 28–35. (In Chinese) [Google Scholar]
- Rathgeber, C.B.K.; Rossi, S.; Bontemps, J.D. Cambial activity related to tree size in a mature silver-fir plantation. Ann. Bot. 2011, 108, 429–438. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Zhang, Y.; Guo, Y.; Kang, M.; Wang, M.; Wang, B. Intra-Annual Xylem Growth of Larix principis-rupprechtii at Its Upper and Lower Distribution Limits on the Luyashan Mountain in North-Central China. Forests 2015, 6, 3809–3827. [Google Scholar] [CrossRef] [Green Version]
- Delpierre, N.; Lireux, S.; Hartig, F.; Camarero, J.J.; Cheaib, A.; Čufar, K.; Cuny, H.; Deslauriers, A.; Fonti, P.; Gričar, J.; et al. Chilling and forcing temperatures interact to predict the onset of wood formation in Northern Hemisphere conifers. Glob. Chang. Biol. 2019, 25, 1089–1105. [Google Scholar] [CrossRef]
- Eilmann, B.; Zweifel, R.; Buchmann, N.; Pannatier, E.G.; Rigling, A. Drought alters timing, quantity, and quality of wood formation in Scots pine. J. Exp. Bot. 2011, 62, 2763–2771. [Google Scholar] [CrossRef] [Green Version]
- Gruber, A.; Strobl, A.; Veit, B.; Oberhuber, W. Impact of drought on the temporal dynamics of wood formation in Pinus sylvestris. Tree Physiol. 2010, 30, 490–501. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Fang, K.; Li, Y.; Dong, Z.; Zhang, Y.; Zhou, F. Response of Pinus taiwanensis growth to climate changes at its southern limit of Daiyun Mountain, mainland China Fujian Province. Sci. China Earth Sci. 2016, 59, 328–336. [Google Scholar] [CrossRef]
- Cai, Q.; Liu, Y.; Qian, H.; Liu, R. Inverse effects of recent warming on trees growing at the low and high altitudes of the Dabie Mountains, subtropical China. Dendrochronologia 2019. [Google Scholar] [CrossRef]
- Deslauriers, A.; Huang, J.-G.; Balducci, L.; Beaulieu, M.; Rossi, S. The contribution of carbon and water in modulating wood formation in black spruce saplings. Plant Physiol. 2016, 170, 2072–2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagedorn, F.; Joseph, J.; Peter, M.; Luster, J.; Pritsch, K.; Geppert, U.; Kerner, R.; Molinier, V.; Egli, S.; Schaub, M. Recovery of trees from drought depends on belowground sink control. Nat. Plants 2016, 2, 16111. [Google Scholar] [CrossRef] [PubMed]
- Hacket-Pain, A.J.; Ascoli, D.; Vacchiano, G.; Biondi, F.; Cavin, L.; Conedera, M.; Drobyshev, I.; Linan, I.D.; Friend, A.D.; Grabner, M. Climatically controlled reproduction drives interannual growth variability in a temperate tree species. Ecol. Lett. 2018, 21, 1833–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauder, J.D.; Moran, E.V.; Hart, S.C. Fight or flight? Potential tradeoffs between drought defense and reproduction in conifers. Tree Physiol. 2019, 39, 1071–1085. [Google Scholar] [CrossRef]
- De Luis, M.; Gričar, J.; Čufar, K.; Raventós, J. Seasonal dynamics of wood formation in Pinus halepensis from dry and semi-arid ecosystems in Spain. IAWA J. 2007, 28, 389–404. [Google Scholar] [CrossRef] [Green Version]
- Vieira, J.; Campelo, F.; Rossi, S.; Carvalho, A.; Freitas, H.; Nabais, C. Adjustment Capacity of Maritime Pine Cambial Activity in Drought-Prone Environments. PLoS ONE 2015, 10, e0126223. [Google Scholar] [CrossRef] [Green Version]
- Fajstavr, M.; Bednářová, E.; Nezval, O.; Giagli, K.; Gryc, V.; Vavrćík, H.; Horáček, P.; Urban, J. How needle phenology indicates the changes of xylem cell formation during drought stress in Pinus sylvestris L. Dendrochronologia 2019, 56, 125600. [Google Scholar] [CrossRef]
- Ren, P.; Ziaco, E.; Rossi, S.; Biondi, F.; Prislan, P.; Liang, E. Growth rate rather than growing season length determines wood biomass in dry environments. Agric. For. Meteorol. 2019, 271, 46–53. [Google Scholar] [CrossRef]
- King, G.M.; Gugerli, F.; Fonti, P.; Frank, D.C. Tree growth response along an elevational gradient: Climate or genetics? Oecologia 2013, 173, 1587–1600. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Gou, X.; Deng, Y.; Zhang, R.; Liu, W.; Zhang, F.; Lu, M.; Chen, Y.; Zheng, W. Tree growth response of Fokienia hodginsii to recent climate warming and drought in southwest China. Int. J. Biometeorol. 2017, 61, 2085–2096. [Google Scholar] [CrossRef]
- Zhou, G.; Peng, C.; Li, Y.; Liu, S.; Zhang, Q.; Tang, X.; Liu, J.; Yan, J.; Zhang, D.; Chu, G. A climate change-induced threat to the ecological resilience of a subtropical monsoon evergreen broad-leaved forest in Southern China. Glob. Chang. Biol. 2013, 19, 1197–1210. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elevations (m) | Latitude | Longitude | Number of Trees (n) | DBH (cm) | Age (year) | Height (m) |
|---|---|---|---|---|---|---|
| 921 | 29°33′26″ | 115°57′53″ | 3 | 19.3 ± 1.8 | 32 ± 3 | 7.7 ± 0.9 |
| 1170 | 29°33′36″ | 115°58′39″ | 5 | 19.7 ± 5.1 | 33 ± 8 | 5.9 ± 0.4 |
| 1402 | 29°33′40″ | 115°59′15″ | 3 | 20.1 ± 2.4 | 33 ± 4 | 5.9 ± 0.8 |
| Months | Year | 921 m | 1170 m | 1402 m | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ta | Pr | SWC | VPD | Ta | Pr | SWC | VPD | Ta | Pr | SWC | VPD | ||
| January–December | 2017 | 13.3 | 1493 | 0.17 | 0.26 | 12.6 | 1111 | 0.17 | 0.26 | 11.5 | 2343 | 0.30 | 0.19 |
| 2018 | 13.1 | 1167 | 0.15 | 0.24 | 12.4 | 974 | 0.15 | 0.23 | 11.2 | 2008 | 0.28 | 0.16 | |
| March–June | 2017 | 14.1 | 713 | 0.21 | 0.26 | 13.4 | 591 | 0.20 | 0.27 | 12.2 | 1040 | 0.33 | 0.19 |
| 2018 | 15.9 | 559 | 0.18 | 0.27 | 15.0 | 487 | 0.18 | 0.25 | 13.7 | 1006 | 0.31 | 0.16 | |
| July–September | 2017 | 22.5 | 595 | 0.15 | 0.33 | 21.6 | 374 | 0.17 | 0.26 | 20.1 | 1006 | 0.31 | 0.13 |
| 2018 | 22.1 | 280 | 0.11 | 0.30 | 21.1 | 291 | 0.14 | 0.24 | 19.6 | 584 | 0.26 | 0.11 | |
| Year | Sites | A | β | κ (10−2) | y0 (10−2) | R2 | rmax (µm d−1) | rmean (µm d−1) |
|---|---|---|---|---|---|---|---|---|
| 2017 | 921 m | 0.38 ± 0.25 | 6.20 ± 3.44 | 4.56 ± 2.71 | 0.47 ± 1.46 | 0.98 | 4.79 ± 1.76 b | 2.93 ± 1.08 b |
| 1170 m | 1.26 ± 0.33 | 3.85 ± 0.96 | 2.77 ± 0.81 | −3.69 ± 2.42 | 0.99 | 12.43 ± 3.52 a | 7.60 ± 2.15 a | |
| 1402 m | 1.77 ± 0.21 | 2.92 ± 0.23 | 1.93 ± 0.18 | −0.36 ± 3.40 | 0.99 | 12.55 ± 1.50 a | 7.67 ± 0.92 a | |
| 2018 | 921 m | 0.28 ± 0.18 | 4.33 ± 2.86 | 3.59 ± 2.08 | 1.07 ± 2.19 | 0.97 | 3.60 ± 2.11 B | 2.20 ± 1.29 B |
| 1170 m | 1.28 ± 0.49 | 3.38 ± 0.55 | 2.66 ± 0.53 | −2.04 ± 1.64 | 0.99 | 11.99 ± 3.53 A | 7.33 ± 2.16 A | |
| 1402 m | 1.84 ± 0.27 | 2.71 ± 0.09 | 1.88 ± 0.04 | −1.54 ± 0.64 | 0.99 | 12.73 ± 1.92 A | 7.79 ± 1.17 A |
| Year | Sites | Onset (DOY) | Peak (DOY) | Offset (DOY) | Ta at Onset | Ta_min at Onset | Ta at Offset | Pr at Offset | SWC at Offset |
|---|---|---|---|---|---|---|---|---|---|
| 2017 | 921 m | 102 ± 10 a | 138 ± 5 b | 212 ± 15 c | 12.4 | 8.6 | 25.8 | 0 | 0.06 |
| 1170 m | 99 ± 5 a | 140 ± 8 ab | 273 ± 15 b | 9.3 | 5.7 | 20.1 | 23.6 | 0.15 | |
| 1402 m | 95 ± 7 a | 151 ± 8 a | 310 ± 11 a | 7.7 | 4.5 | 9.6 | 1.4 | 0.23 | |
| 2018 | 921 m | 89 ± 9 A | 131 ± 11 B | 206 ± 14 C | 12.7 | 8.9 | 24.8 | 0 | 0.07 |
| 1170 m | 86 ± 3 A | 128 ± 6 B | 260 ± 8 B | 9.5 | 6.7 | 18.1 | 11 | 0.12 | |
| 1402 m | 86 ± 6 A | 144 ± 6 A | 312 ± 12 A | 7.8 | 4.4 | 10.2 | 27.6 | 0.22 |
| Independent Variables | 921 m | 1170 m | 1402 m | All |
|---|---|---|---|---|
| Timing of growth onset (DOY) | −0.64 | −0.54 | −0.35 | −0.22 |
| Timing of growth offset (DOY) | 0.97 * | 0.83 * | 0.53 | 0.75 *** |
| Mean growth rate (μm d−1) | 0.78 | 0.91 ** | 0.85 * | 0.87 *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Wang, C.; Zhao, J. Seasonal Drought Effects on Intra-Annual Stem Growth of Taiwan Pine along an Elevational Gradient in Subtropical China. Forests 2019, 10, 1128. https://doi.org/10.3390/f10121128
Liu X, Wang C, Zhao J. Seasonal Drought Effects on Intra-Annual Stem Growth of Taiwan Pine along an Elevational Gradient in Subtropical China. Forests. 2019; 10(12):1128. https://doi.org/10.3390/f10121128
Chicago/Turabian StyleLiu, Xinsheng, Chunsong Wang, and Junkai Zhao. 2019. "Seasonal Drought Effects on Intra-Annual Stem Growth of Taiwan Pine along an Elevational Gradient in Subtropical China" Forests 10, no. 12: 1128. https://doi.org/10.3390/f10121128
APA StyleLiu, X., Wang, C., & Zhao, J. (2019). Seasonal Drought Effects on Intra-Annual Stem Growth of Taiwan Pine along an Elevational Gradient in Subtropical China. Forests, 10(12), 1128. https://doi.org/10.3390/f10121128