Hormones and Heterosis in Hybrid Balsam Poplar (Populus balsamifera L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Families and Progeny
2.2. Greenhouse Propagation
2.3. Measurements, Harvest and Selection for Hormone Analysis
2.4. Analysis of GAs, IAA, and ABA
2.5. Data Analysis
3. Results
3.1. Hybrid Vigour in Intra-Specific Hybrids
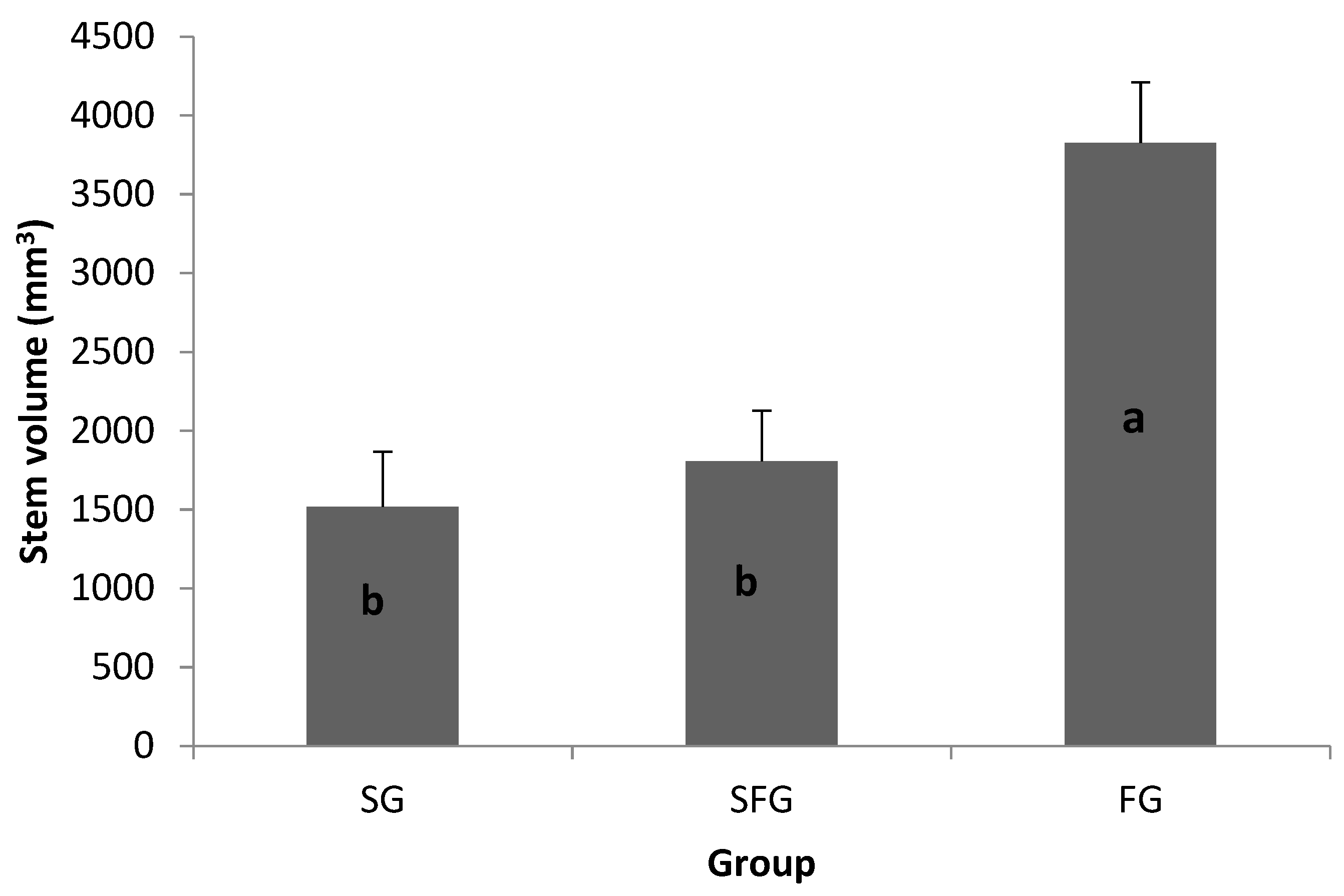
3.2. Greenhouse Growth at Two Months (34 Days after Transplanting)
3.3. Greenhouse Gas Exchange
3.4. Comparisons of Greenhouse Growth at Day 34 and Field Growth Performance at Age Eight Years
3.5. Hormone Analysis
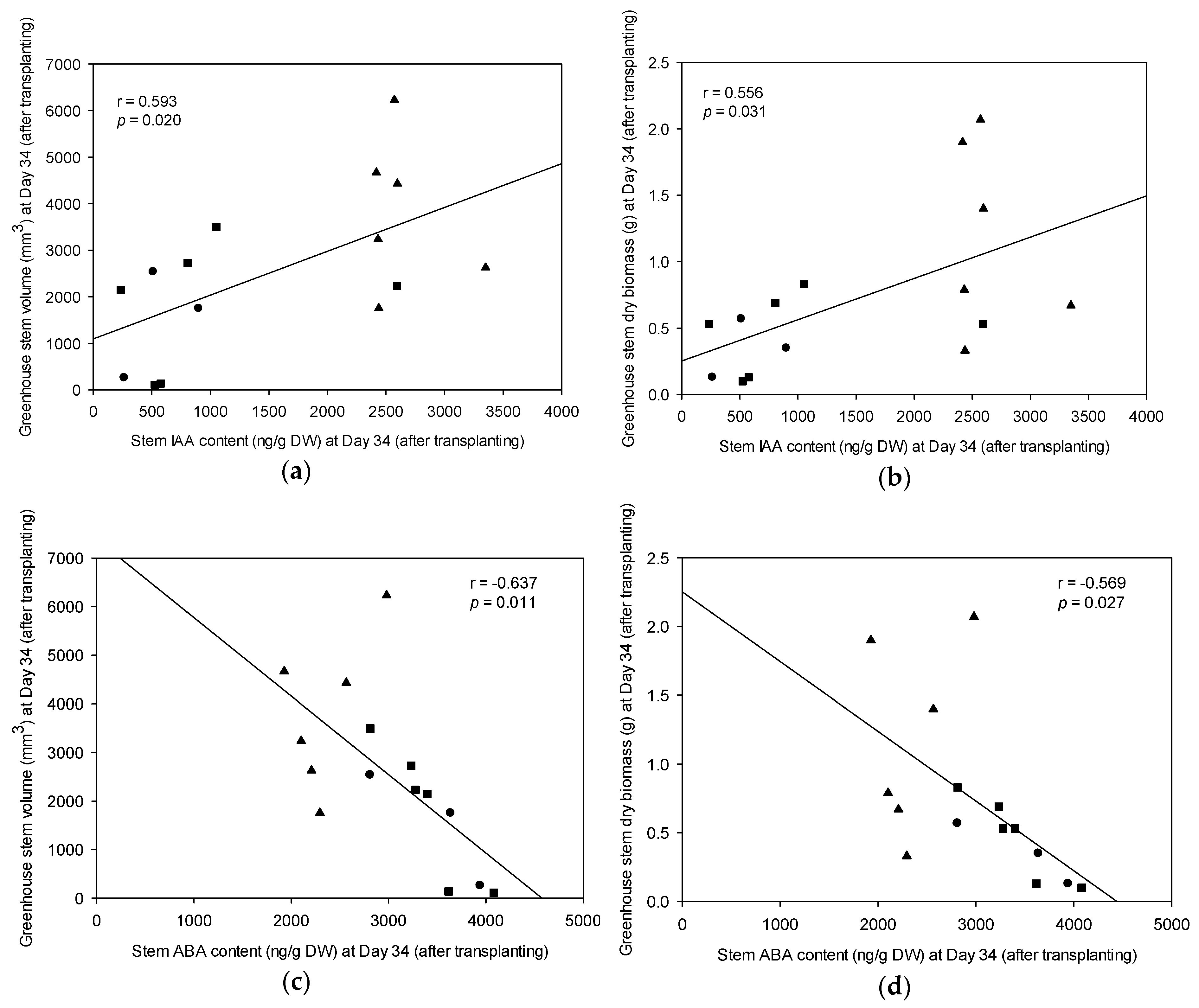
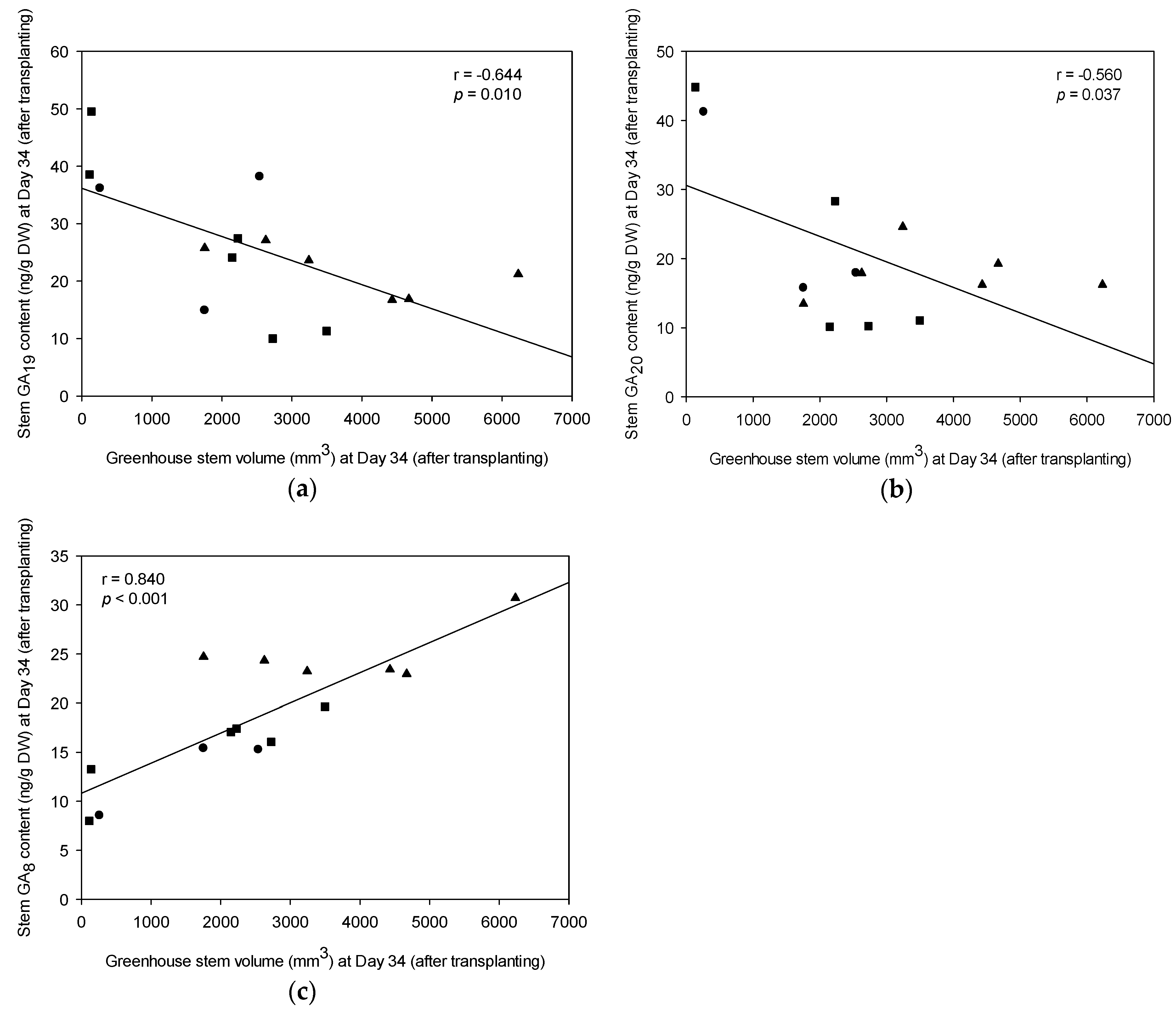
3.6. Endogenous Plant Growth Hormone in Greenhouse-Grown Stecklings versus Field Growth Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sinha, S.K.; Khanna, R. Physiological, biochemical, and genetic basis of heterosis. Adv. Agron. 1975, 27, 123–174. [Google Scholar]
- Wehrhahn, C.; Allard, R.W. The detection and measurement of the effects of individual genes involved in the inheritance of a quantitative character in wheat. Genetics 1965, 51, 109–119. [Google Scholar] [PubMed]
- Stuber, C.W. Heterosis in plant breeding. Plant Breed Rev. 1994, 12, 227–251. [Google Scholar]
- Baranwal, V.K.; Mikkilineni, V.; Zehr, U.B.; Tyagi, A.K.; Kapoor, S. Heterosis: Emerging ideas about hybrid vigour. EXBOTJ 2012, 63, 6309–6314. [Google Scholar] [CrossRef] [PubMed]
- Stettler, R.F.; Zsuffa, L.; Wu, R. The role of hybridization in the genetic manipulation of Populus. In Biology of Populus and its Implications for Management and Conservation; Part I, Chapter 4; Stettler, R., Bradshaw, T., Heilman, P., Hinckley, T., Eds.; NRC (National Research Council of Canada) Research Press: Ottawa, ON, Canada, 1996; pp. 87–112. [Google Scholar]
- Zasada, J.C.; Phipps, H.M. Populus balsamifera L. Balsam Poplar. In Silvics of North America Volume 2. Hardwoods; Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990; Volume 2, p. 877. [Google Scholar]
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Ray, N. Surfing during population expansions promotes genetic revolutions and structuration. Trends Ecol. Evol. 2008, 23, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.L.; Cornman, R.S.; Martin, N.H. Hybridization, hybrid fitness and the evolution of adaptations. Plant Biosyst. 2008, 142, 166–171. [Google Scholar] [CrossRef]
- Korn, M.; Gärtner, T.; Erban, A.; Kopka, J.; Selbig, J.; Hincha, D.K. Predicting Arabidopsis Freezing Tolerance and Heterosis in Freezing Tolerance from Metabolite Composition. Molecular Plant 2010, 3, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Hedden, P.; Zhang, Q. Heterosis in Rice Seedlings: Its Relationship to Gibberellin Content and Expression of Gibberellin Metabolism and Signaling Genes. Plant Physiol. 2011, 156, 1905–1920. [Google Scholar] [CrossRef] [PubMed]
- Olszewski, N.; Sun, T.-P.; Gubler, F. Gibberellin signaling: biosynthesis, catabolism, and response pathways. Plant Cell 2002, 14, S61–S80. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.G.; Hedden, P. Gibberellin biosynthesis and its regulation. Biochem. J. 2012, 444, 11–25. [Google Scholar]
- Kurepin, L.V.; Janzen, L.; Park, E.-J.; Lee, W.-Y.; Zhang, R.; Pharis, R.P. Plant hormone-assisted early family selection in Pinus densiflora via a retrospective approach. Tree Physiol. 2014, 35, 86–94. [Google Scholar]
- Ueguchi-Tanaka, M.; Fujisawa, Y.; Kobayashi, M.; Ashikari, M.; Iwasaki, Y.; Kitano, H.; Matsuoka, M. Rice dwarf mutant d1, which is defective in the alpha subunit of the heterotrimeric G protein, affects gibberellin signal transduction. Proc. Natl. Acad. Sci. USA 2000, 97, 11638–11643. [Google Scholar] [CrossRef] [PubMed]
- Rieu, I.; Eriksson, S.; Powers, S.J.; Gong, F.; Griffiths, J.; Woolley, L.; Benlloch, R.; Nilsson, O.; Thomas, S.G.; Hedden, P.; et al. Genetic analysis reveals that C19-GA 2-oxidation is a major gibberellin inactivation pathway in Arabidopsis. Plant Cell. 2008, 20, 2420–2436. [Google Scholar] [CrossRef] [PubMed]
- Rood, S.B.; Buzzell, R.I.; Mander, L.N.; Pearce, D.; Pharis, R.P. Gibberellins: a phytohormonal basis for heterosis in maize. Science 1988, 241, 1216–1218. [Google Scholar] [CrossRef] [PubMed]
- Uggla, C.; Moritz, T.; Sandberg, G.; Sundberg, B. Auxin as a positional signal in pattern formation in plants. Proc. Natl. Acad. Sci. USA 1996, 93, 9282–9286. [Google Scholar] [CrossRef] [PubMed]
- Tuominen, H.; Puech, L.; Fink, S.; Sundberg, B. A radial concentration gradient of indole-3-acetic acid is related to secondary xylem development in hybrid aspen. Plant Physiol. 1997, 115, 577–585. [Google Scholar] [CrossRef]
- Uggla, C. Indole-3-Acetic Acid Controls Cambial Growth in Scots Pine by Positional Signaling. Plant Physiol. 1998, 117, 113–121. [Google Scholar] [CrossRef]
- Fu, S.-F.; Wei, J.-Y.; Chen, H.-W.; Liu, Y.-Y.; Lu, H.-Y.; Chou, J.-Y. Indole-3-acetic acid: A widespread physiological code in interactions of fungi with other organisms. Plant Signal. Behav. 2015, 10, e1048052. [Google Scholar] [CrossRef]
- Zeevaart, J.A.D.; A Creelman, R. Metabolism and physiology of abscisic acid. Annu. Rev. Plant Biol. 1988, 39, 439–473. [Google Scholar] [CrossRef]
- Lim, C.W.; Baek, W.; Jung, J.; Kim, J.-H.; Lee, S.C. Function of ABA in stomatal defense against biotic and drought stresses. IJMS 2015, 16, 15251–15270. [Google Scholar] [CrossRef] [PubMed]
- Climate Edmonton. Available online: https://en.climate-data.org/north-america/canada/alberta/edmonton-610/ (accessed on 16 January 2019).
- Climate Trécesson. Available online: https://en.climate-data.org/north-america/canada/quebec/trecesson-719862/ (accessed on 16 January 2019).
- DesRochers, A.; Thomas, B.R. A comparison of pre-planting treatments on hardwood cuttings of four hybrid poplar clones. New Forests 2003, 26, 17–32. [Google Scholar] [CrossRef]
- Drewes, E.C.; Soolanayakanahally, R.Y.; Guy, R.D.; Silim, S.N.; Schroeder, W.R. Enhanced assimilation rate and water use efficiency with latitude through increased photosynthetic capacity and internal conductance in balsam poplar (Populus balsamifera L.). Plant Cell Environ. 2009, 32, 1821–1832. [Google Scholar]
- Kurepin, L.V.; Emery, R.J.N.; Pharis, R.P.; Reid, D.M. Uncoupling light quality from light irradiance effects in Helianthus annuus shoots: putative roles for plant hormones in leaf and internode growth. EXBOTJ 2007, 58, 2145–2157. [Google Scholar] [CrossRef]
- SAS Institute Inc. Base SAS® 9.4 Procedures Guide: Statistical Procedures, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Zawaski, C.; Busov, V.B. Roles of gibberellin catabolism and signaling in growth and physiological response to drought and short-day photoperiods in Populus trees. PLoS ONE 2014, 9, e86217. [Google Scholar] [CrossRef]
- Ford, K.R.; Harrington, C.A.; Clair, J.B.S. Photoperiod cues and patterns of genetic variation limit phenological responses to climate change in warm parts of species’ range: modeling diameter-growth cessation in coast Douglas-fir. Glob. Change Biol. 2017, 23, 3348–3362. [Google Scholar] [CrossRef]
- Rohde, A.; Bastien, C.; Boerjan, W.; Thomas, S. Temperature signals contribute to the timing of photoperiodic growth cessation and bud set in poplar. Tree Physiol. 2011, 31, 472–482. [Google Scholar] [CrossRef]
- Schmidtling, R.C.; Nelson, C.D. Interprovenance crosses in loblolly pine using selected parents. Forest Genet. 1996, 3, 53–66. [Google Scholar]
- Harfouche, A.; Bahrman, N.; Baradat, P.; Guyon, J.P.; Petit, R.J.; Kremer, A. Provenance hybridization in a diallel mating scheme of maritime pine (Pinus pinaster). II. Heterosis. Can. J. For. Res. 2000, 30, 10–16. [Google Scholar] [CrossRef]
- Kirk, H.; Vrieling, K.; Klinkhamer, P.G.L. Maternal effects and heterosis influence the fitness of plant hybrids. New Phytol. 2005, 166, 685–694. [Google Scholar] [CrossRef]
- Bräutigam, K.; Soolanayakanahally, R.; Champigny, M.; Mansfield, S.; Douglas, C.; Campbell, M.M.; Cronk, Q. Sexual epigenetics: gender-specific methylation of a gene in the sex determining region of Populus balsamifera. Sci. Rep. 2017, 7, 45388. [Google Scholar] [CrossRef] [PubMed]
- Pharis, R.P.; Yeh, F.C.; Dancik, B.P. Superior growth potential in trees: What is its basis, and can it be tested for at an early age? Can. J. For. Res. 1991, 21, 368–374. [Google Scholar] [CrossRef]
- Williams, D.J.; Dancik, B.P.; Pharis, R.P. Early progeny testing and evaluation of controlled crosses of black spruce. Can. J. For. Res. 1987, 17, 1442–1450. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Körner, C.; Farquhar, G.D.; Wong, S.C. Carbon isotope discrimination by plants follows latitudinal and altitudinal trends. Oecologia 1991, 88, 30–40. [Google Scholar] [CrossRef]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; DeLay, D.; Petit, J.-M.; Barbaroux, C.; Le Thiec, D.; Bréchet, C.; Brignolas, F. Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides × Populus nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef]
- Chamaillard, S.; Fichot, R.; Vincent-Barbaroux, C.; Bastien, C.; Depierreux, C.; Dreyer, E.; Villar, M.; Brignolas, F. Variations in bulk leaf carbon isotope discrimination, growth and related leaf traits among three Populus nigra L. populations. Tree Physiol. 2011, 31, 1076–1087. [Google Scholar] [CrossRef]
- Guy, R.D.; Holowachuk, D.L. Population differences in stable carbon isotope ratio of Pinus contorta Dougl. ex Loud.: relationship to environment, climate of origin, and growth potential. Can. J. Bot. 2001, 79, 274–283. [Google Scholar] [CrossRef]
- Diefendorf, A.F.; Mueller, K.E.; Wing, S.L.; Koch, P.L.; Freeman, K.H. Global patterns in leaf 13C discrimination and implications for studies of past and future climate. Proc. Natl. Acad. Sci. USA 2010, 107, 5738–5743. [Google Scholar] [CrossRef]
- Eilmann, B.; de Vries, S.M.G.; den Ouden, J.; Mohren, G.M.J.; Sauren, P.; Sass-Klaassen, U. Origin matters! Difference in drought tolerance and productivity of coastal Douglas-fir (Pseudotsuga menziesii (Mirb.)) provenances. Forest Ecol. Manag. 2013, 302, 133–143. [Google Scholar] [CrossRef]
- Wolbang, C.M.; Ross, J.J. Auxin promotes gibberellin biosynthesis in decapitated tobacco plants. Planta 2001, 214, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Ozga, J.A.; Yu, J.; Reinecke, D.M. Pollination-, development-, and auxin-specific regulation of gibberellins 3β-hydroxylase gene expression in pea fruit and seeds. Plant Physiol. 2003, 131, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- Nemhauser, J.L.; Hong, F.; Chory, J. Different plant hormones regulate similar processes through largely non-overlapping transcriptional responses. Cell 2006, 126, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, M.A.; Liu, J.; Enders, T.A.; Bartel, B.; Strader, L.C. A gain-of-function mutation in IAA16 confers reduced responses to auxin and abscisic acid and impedes plant growth and fertility. Plant Mol. Biol. 2012, 79, 359–373. [Google Scholar] [CrossRef] [PubMed]
- Kurepin, L.V.; Walton, L.J.; Pharis, R.P.; Emery, R.J.N.; Reid, D.M. Interactions of temperature and light quality on phytohormone-mediated elongation of Helianthus annuus hypocotyls. Plant Growth Regul. 2010, 64, 147–154. [Google Scholar] [CrossRef]
- Hedden, P.; Phillips, A.L. Gibberellin metabolism: new insights revealed by the genes. Trends Plant Sci. 2000, 5, 523–530. [Google Scholar] [CrossRef]
- Eriksson, M.E.; Israelsson, M.; Olsson, O.; Moritz, T. Increased gibberellin biosynthesis in transgenic trees promotes growth, biomass production and xylem fiber length. Nat. Biotechnol. 2000, 18, 784–788. [Google Scholar] [CrossRef] [PubMed]
- Busov, V.B.; Meilan, R.; Pearce, D.W.; Ma, C.; Rood, S.B.; Strauss, S.H. Activation tagging of a dominant gibberellin catabolism gene (GA 2-oxidase) from Poplar that regulates tree stature. Plant physiol. 2003, 132, 1283–1291. [Google Scholar] [CrossRef]
- Reinecke, D.M.; Wickramarathna, A.D.; Ozga, J.; Kurepin, L.V.; Jin, A.L.; Good, A.G.; Pharis, R.P. Gibberellin 3-oxidase gene expression patterns influence gibberellin biosynthesis, growth, and development in Pea. Plant physiol. 2013, 163, 929–945. [Google Scholar] [CrossRef]
- Yao, Y.; Nie, X.; Zhang, Y.; Ni, Z.; Sun, Q. Gibberellins and heterosis of plant height in wheat (Triticum aestivum L.). BMC Genet. 2007, 8, 40. [Google Scholar]
- Park, E.J.; Kim, H.T.; Choi, Y.I.; Lee, C.H.; Nguyen, V.P.; Jeon, H.W.; Cho, J.S.; Pharis, R.P.; Kurepin, L.V.; et al. Overexpression of gibberellin 20-oxidase1 from Pinus densiflora results in enhanced wood formation with gelatinous fiber development in a trangenic hybrid poplar. Tree Physiol. 2015, 35, 1264–1277. [Google Scholar] [PubMed]
- Ko, J.-H.; Kim, H.-T.; Han, K.-H.; Ko, J.; Kim, H.; Hwang, I.; Han, K. Tissue-type-specific transcriptome analysis identifies developing xylem-specific promoters in poplar. Plant Biotechnol. J. 2012, 10, 587–596. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Fast-Growing Progeny | Group (Fast-Growing (FG)) | Slow-Growing Progeny | Group (Slow-Growing in Fast- Growing Group (SFG) or Slow-Growing (SG) Group) |
|---|---|---|---|---|
| AP5396 | 147071 | FG | 147043 | SFG |
| 147083 | FG | 147051 | SFG | |
| 147072 | FG | 147041 | SFG | |
| AP5401 | 178092 | SG | ||
| 178073 | SG | |||
| 178104 | SG | |||
| AP5402 | 180071 | FG | 180093 | SFG |
| 180103 | FG | 180063 | SFG | |
| 180081 | FG | 180094 | SFG | |
| AP5411 | 255081 | SG | ||
| 255011 | SG | |||
| 255102 | SG | |||
| AP5414 | 270051 | SG | ||
| 270101 | SG | |||
| 270061 | SG | |||
| AP5416 | 272084 | FG | 272024 | SFG |
| 272102 | FG | 272071 | SFG | |
| 272091 | FG | 272023 | SFG |
| Cross-Types | Height (m) | DBH (mm) | Stem Volume (cm3) |
|---|---|---|---|
| AB♀ × AB♂ | 3.67 ± 0.05 c | 33.70 ± 0.76 b | 1375.94 ± 65.88 b |
| AB♀ × QC♂ | 4.06 ± 0.04 a | 37.77 ± 0.54 a | 1786.78 ± 59.48 a |
| QC♀ × AB♂ | 3.78 ± 0.03 bc | 34.73 ± 0.40 b | 1389.78 ± 35.27 b |
| QC♀ × QC♂ | 3.80 ± 0.05 b | 34.52 ± 0.63 b | 1433.09 ± 55.63 b |
| Height (cm) | Caliper (mm) | Stem Volume (mm3) | A | gs | iWUE | δ13Cleaf (‰) | |
|---|---|---|---|---|---|---|---|
| Height (cm) | 1 | 0.933 ** | 0.861 ** | 0.751 * | 0.532 * | 0.606 * | −0.545 ** |
| Caliper (mm) | 1 | 0.890 ** | 0.743 * | 0.614 * | 0.517 * | −0.502 | |
| Stem volume (mm3) | 1 | 0.556 * | 0.327 | 0.344 | −0.495 | ||
| A | 1 | 0.596 * | 0.796 ** | −0.461 | |||
| gs | 1 | 0.205 | −0.563 * | ||||
| iWUE | 1 | −0.505 * |
| Field AB1 | ||
| Stem parameters of 8-year-old trees at FieldAB1 | Day 34 stem volume (mm3) | Day 34 stem dry weight (g) |
| Height (cm) | 0.550 *1 | 0.516 * |
| DBH (mm) | 0.608 * | 0.584 * |
| Stem volume (cm3) | 0.629 * | 0.601 * |
| Field AB2 | ||
| Stem parameters of 8-year-old trees at | Day 34 stem volume (mm3) | Day 34 stem dry weight (g) |
| Height (cm) | 0.654 ** | 0.686 ** |
| DBH (mm) | 0.537 * | 0.604 * |
| Stem volume (cm3) | 0.619 * | 0.671 ** |
| Field QC1 | ||
| Stem parameters of 6-year-old trees at | Day 34 stem volume (mm3) | Day 34 stem dry weight (g) |
| Height (cm) | 0.758 ** | 0.739 ** |
| DBH (mm) | 0.666 ** | 0.705 ** |
| Stem volume (cm3) | 0.588 * | 0.684 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Thomas, B.R. Hormones and Heterosis in Hybrid Balsam Poplar (Populus balsamifera L.). Forests 2019, 10, 143. https://doi.org/10.3390/f10020143
Hu Y, Thomas BR. Hormones and Heterosis in Hybrid Balsam Poplar (Populus balsamifera L.). Forests. 2019; 10(2):143. https://doi.org/10.3390/f10020143
Chicago/Turabian StyleHu, Yue, and Barb R. Thomas. 2019. "Hormones and Heterosis in Hybrid Balsam Poplar (Populus balsamifera L.)" Forests 10, no. 2: 143. https://doi.org/10.3390/f10020143
APA StyleHu, Y., & Thomas, B. R. (2019). Hormones and Heterosis in Hybrid Balsam Poplar (Populus balsamifera L.). Forests, 10(2), 143. https://doi.org/10.3390/f10020143