Host Phylogenetic Relatedness and Soil Nutrients Shape Ectomycorrhizal Community Composition in Native and Exotic Pine Plantations

 , and
, and
Abstract
:1. Introduction
- (1)
- Do closely related exotic and native host species support similar ECM communities?
- (2)
- To what extent do neutral (dispersal) versus niche (abiotic factors) shape ECM assemblages in exotic and native pines?
2. Materials and Methods
2.1. Study Sites
2.2. Sampling Methods
2.3. Soil Physicochemical Analysis
2.4. Molecular Identification of Ectomycorrhizal Fungal Community
2.5. Statistical Analyses
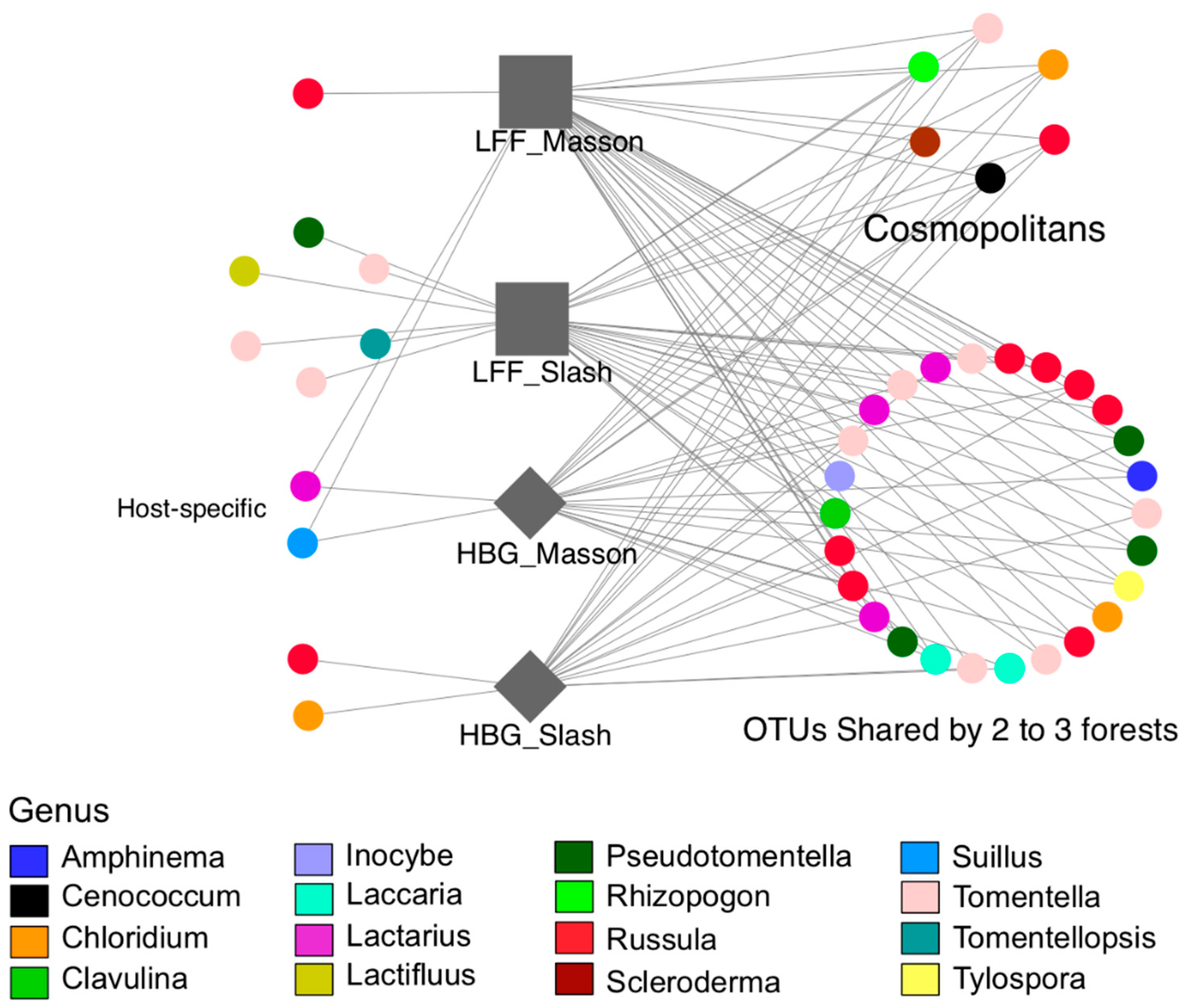
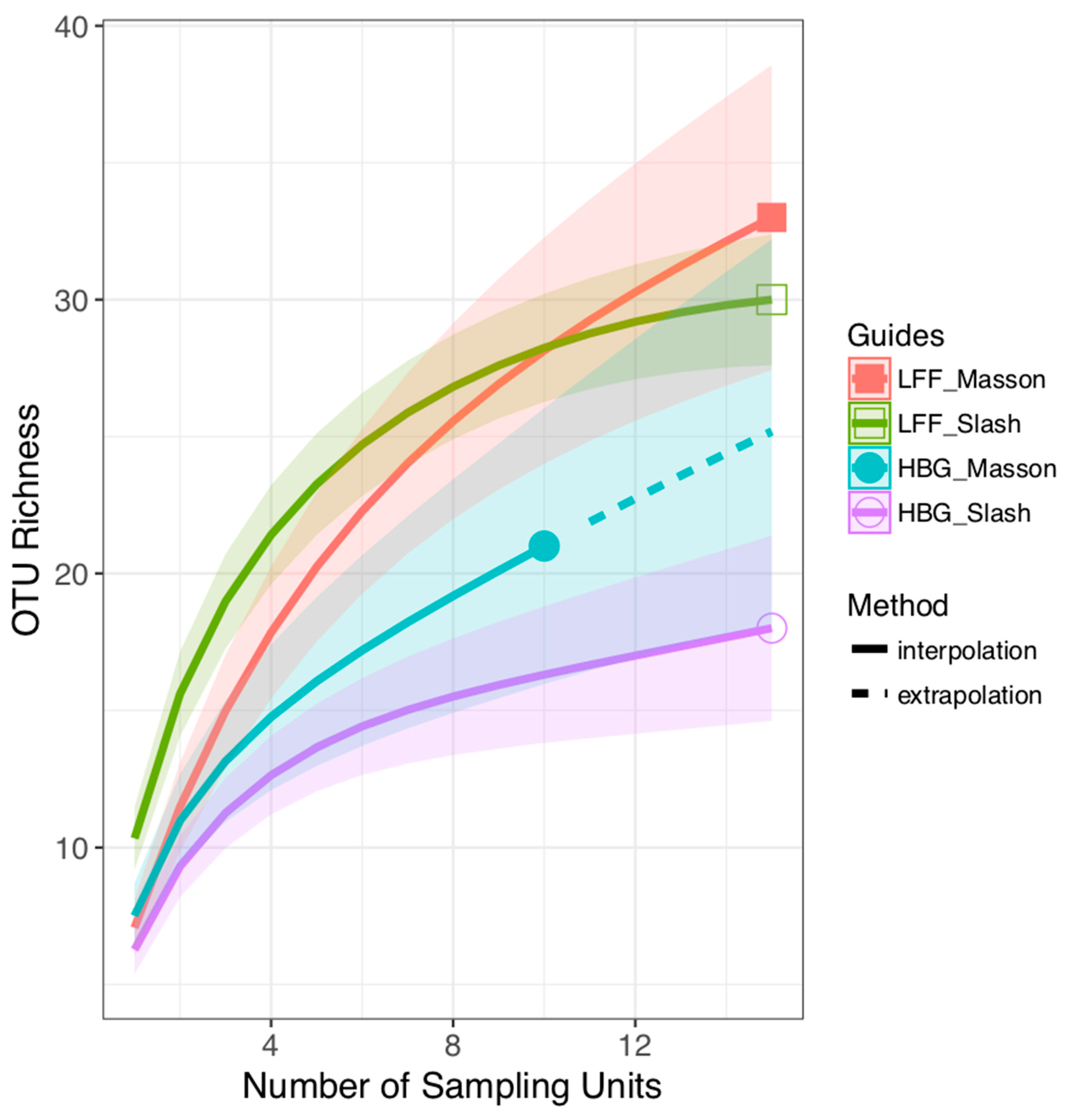
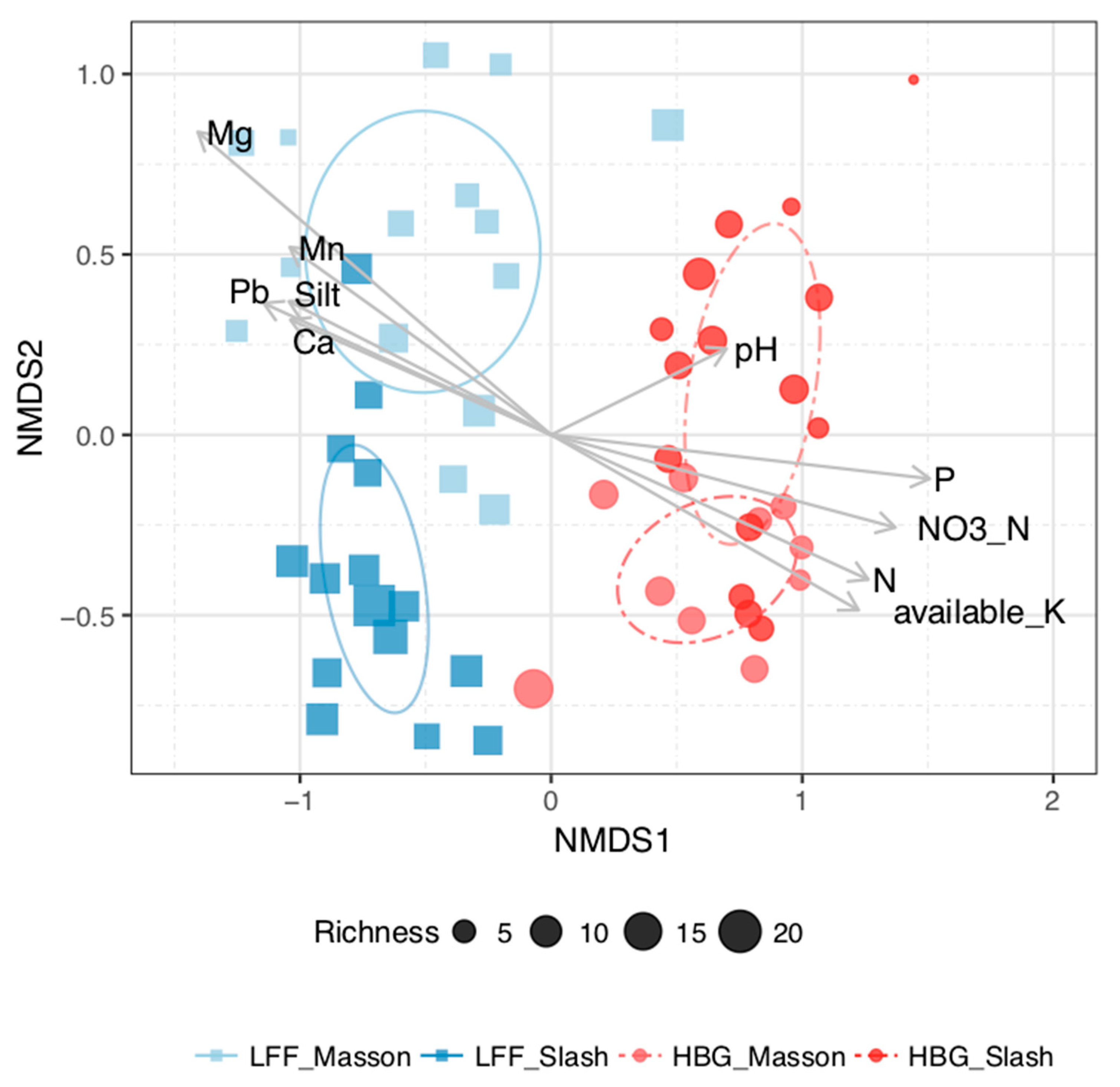
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Richardson, D.M.; Higgins, S.I. Ecology and biogeography of Pinus. In Pines as Invaders in the Southern Hemisphere; Richardson, D.M., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 243–266. [Google Scholar]
- Wardle, D.A. Communities and Ecosystems: Linking the Aboveground and Belowground Components; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Van der Putten, W.H.; Klironomos, J.N.; Wardle, D.A. Microbial ecology of biological invasions. ISME J. 2007, 1, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuñez, M.A.; Horton, T.R.; Simberloff, D. Lack of below- ground mutualisms hinders Pinaceae invasions. Ecology 2009, 90, 2352–2359. [Google Scholar] [CrossRef] [PubMed]
- Vellinga, E.C.; Wolfe, B.E.; Pringle, A. Global patterns of ectomycorrhizal introductions. New Phytol. 2009, 181, 960–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikola, P. Mycorrhizal inoculation in afforestation. Int. Rev. For. Res. 1970, 3, 123–196. [Google Scholar]
- Richardson, D.M.; Allsopp, N.; D’Antonio, C.M.; Milton, S.J.; Rejmanek, M. Plant invasions—The role ofmutualisms. Biol. Rev. 2000, 75, 65–93. [Google Scholar] [CrossRef] [PubMed]
- Trocha, L.K.; Kalucka, I.; Stasińska, M.; Nowak, W.; Dabert, M.; Leski, T.; Rudawska, M.; Oleksyn, J. Ectomycorrhizal fungal communities of native and non-native Pinus and Quercus species in a common garden of 35-year-old trees. Mycorrhiza 2012, 22, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Dickie, I.A.; Bolstridge, N.; Cooper, J.A.; Pelzer, D.A. Co-invasion by Pinus and its mycorrhizal fungi. New Phytol. 2010, 187, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Kohout, P.; Sykorova, Z.; Bahram, M.; Hadincova, V.; Alberchtova, J.; Tedersoo, L.; Vohnik, M. Ericaceous dwarf shrub affect ectomycorrhizal fungal community of the invasive Pinus strobus and native Pinus sylvestris in a pot experiment. Mycorrhiza 2011, 21, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Hayward, J.; Horton, T.R.; Pauchard, A.; Nuñez, M.A. A single ectomycorrhizal fungal species can enable a Pinus invasion. Ecology 2015, 96, 497–506. [Google Scholar] [CrossRef]
- Hynson, N.A.; Merckx, V.S.F.T.; Perry, B.A.; Tresder, K.K. Identities and distributions of the co-invading ectomycorrhizal fungal symbionts of exotic pines in the Hawaiian Islands Nicole. Biol. Invasions 2013, 15, 2373–2385. [Google Scholar] [CrossRef]
- Nuñez, M.A.; Dickie, I.A. Invasive belowground mutualists of woody plants. Biol. Invasions 2014, 16, 645–661. [Google Scholar] [CrossRef]
- Bahram, M.; Köljalg, U.; Courty, P.E.; Diedhiou, A.G.; Kjøller, R.; Põlme, S.; Ryberg, M.; Veldre, V.; Tedersoo, L. The distance decay of similarity in communities of ectomycorrhizal fungi in different ecosystems and scales. J. Ecol. 2013, 101, 1335–1344. [Google Scholar] [CrossRef] [Green Version]
- Talbot, J.M.; Bruns, T.D.; Taylor, J.W.; Smith, D.P.; Branco, S.; Glassman, S.I.; Erlandson, S.; Vigalys, R.; Liao, H.L.; Smith, M.E.; et al. Endemism and functional convergence across the North American soil mycobiome. Proc. Natl. Acad. Sci. USA 2014, 111, 6341–6346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glassman, S.I.; Peay, K.G.; Talbot, J.M.; Smith, D.P.; Chung, J.A.; Taylor, J.W.; Vigalys, R.; Bruns, T.D. A continental view of pine-associated ectomycorrhizal fungal spore banks: A quiescent functional guild with a strong biogeographic pattern. New Phytol. 2015, 205, 1619–1631. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, S.; Mori, A.S.; Kawaguchi, E.; Hobara, S.; Osono, T. Disentangling the relative importance of host tree community, abiotic environment and spatial factors on ectomycorrhizal fungal assemblages along an elevation gradient. FEMS Microbiol. Ecol. 2016, 92, fiw044. [Google Scholar] [CrossRef] [PubMed]
- Nara, K. Spores of ectomycorrhizal fungi: Ecological strategies for germination and dormancy. New Phytol. 2009, 181, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Beiler, K.J.; Durall, D.M.; Simard, S.W.; Maxwell, S.A.; Kretzer, A.M. Architecture of the wood-wide web: Rhizopogon spp. genets link multiple Douglas-fir cohorts. New Phytol. 2010, 185, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Pickles, B.J.; Genney, D.R.; Potts, J.M.; Lennon, J.J.; Anderson, I.C.; Alexander, I.J. Spatial and temporal ecology of scots pine ectomycorrhizas. New Phytol. 2010, 186, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Lang, C.; Finkeldey, R.; Polle, A. Spatial patterns of ectomycorrhizal assemblages in a monospecific forest in relation to host tree genotype. Front. Plant Sci. 2013, 4, 103. [Google Scholar] [CrossRef] [PubMed]
- Boeraeve, M.; Honnay, O.; Mullens, N.; Vandekerkhove, K.; Keersmaeker, L.D.; Thomaes, A.; Jacquemyn, H. The impact of spatial isolation and local habitat conditions on colonization of recent forest stands by ectomycorrhizal fungi. For. Ecol. Manag. 2018, 429, 84–92. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Izzo, A.; Robinson, R. Changes in ectomycorrhizal community structure on two containerized oak hosts across an experimental hydrologic gradient. Mycorrhiza 2009, 19, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Roy-Bolduc, A.; Laliberté, E.; Hijri, M. High richness of ectomycorrhizal fungi and low host specificity in a coastal sand dune ecosystem revealed by network analysis. Ecol. Evol. 2016, 6, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Williams, L.J.; Vincent, J.B.; Stefanskyi, A.; Cavender-Bares, J.; Messier, C.; Paquette, A.; Gravel, D.; Reich, P.B.; Kennedy, P.G. Ectomycorrhizal fungal diversity and saprotrophic fungal diversity are linked to different tree community attributes in a field-based tree experiment. Mol. Ecol. 2016, 25, 4032–4066. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.O.; Smith, J.E.; Luoma, D.L.; Jones, M.D. Ectomycorrhizal communities of ponderosa pine and lodgepole pine in the south-central Oregon pumice zone. Mycorrhiza 2016, 26, 275–286. [Google Scholar] [CrossRef] [PubMed]
- O’Hanlon, R.; Harrington, T.J. Diversity and distribution of mushroom forming fungi (Agaricomycetes) in Ireland. Biol. Environ. 2011, 111B, 117–133. [Google Scholar] [CrossRef]
- O’Hanlon, R.; Harrington, T.J. Similar taxonomic richness but different communities of ectomycorrhizas in native forests and non-native plantation forests. Mycorrhiza 2012, 22, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Cullings, K.W.; Vogler, D.R.; Parker, V.T.; Finley, S.K. Ectomycorrhizal specificity patterns in a mixed Pinus contorta and Picea engelmannii forest in Yellowstone National Park. Appl. Environ. Microbiol. 2000, 66, 4988–4991. [Google Scholar] [CrossRef] [PubMed]
- Bahram, M.; Köljalg, U.; Kohout, P.; Mirshahvaladi, S.; Tedersoo, L. Ectomycorrhizal fungi of exotic pine plantations in relation to native host trees in Iran: Evidence of host range expansion by local symbionts to distantly related host taxa. Mycorrhiza 2013, 23, 11–19. [Google Scholar] [CrossRef] [PubMed]
- SFA (State Forestry Administration P.R. China). National Forest Resources Statistics (2009–2013), NFI 8th; State Forestry Administration: Beijing, China, 2014.
- Chen, Q.J.; Shu, Z.Z.; Pan, W.; Long, Y.Z.; Wei, B.S. The cultivation zoning and base layout of slash pine in China. For. Sci. Technol. Guangdong Prov. 1995, 11, 1–6, (In Chinese with English Abstract). [Google Scholar]
- Zhao, T.; Zhao, W.; Gao, R.; Zhang, Q.; Li, G.; Liu, X. Induced outbreaks of indigenous insect species by exotic tree species. Acta Ecol. Sin. 2007, 50, 826–833, (In Chinese with English Abstract). [Google Scholar]
- Huang, P.M.; Li, Y.C.; Summer, M.E. Handbook of Soil Sciences: Properties and Processes, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Wallander, H.; Nilsson, L.O.; Hagerberg, D.; Baath, E. Estimation of the biomass and seasonal growth of external mycelium of ectomycorrhizal fungi in the field. New Phytol. 2001, 151, 753–760. [Google Scholar] [CrossRef] [Green Version]
- Branco, S.; Bruns, T.D.; Singleton, I. Fungi at a small scale: Spatial zonation of fungal assemblages around single trees. PLoS ONE 2013, 8, e78295. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.P.; Peay, K.G. Sequence depth, not PCR replication, improves ecological inference from next generation DNA sequencing. PLoS ONE 2014, 9, e90234. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, R.H.; Veldre, V.; Hartmann, M.; Unterseher, M.; Amend, A.; Bergsten, J.; Kristiansson, E.; Ryberg, M.; Jumpponen, A.; Abarenkov, K. An open source software package for automated extraction of ITS1 and ITS2 from fungal ITS sequences for use in high-throughput community assays and molecular ecology. Fungal Ecol. 2010, 3, 284–287. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedersoo, L.; Nilsson, R.H.; Abarenkov, K.; Jairus, T.; Sadam, A.; Saar, I.; Bahram, M.; Bechem, E.; Chuyong, G.; Kõljalg, U. 454 pyrosequencing and Sanger sequencing of tropical mycorrhizal fungi provide similar results but reveal substantial methodological biases. New Phytol. 2010, 188, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Ihrmark, K.; Bödeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubatova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandström-Durling, M.; Clemmensen, C.E.; et al. New primers to amplify the fungal ITS2 region—Evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2015, 24, 241–248. [Google Scholar] [CrossRef]
- Tedersoo, L.; Smith, M.E. Lineages of ectomycorrhizal fungi revisited: Foraging strategies and novel lineages revealed by sequences from belowground. Fungal Biol. Rev. 2013, 27, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.H.; Smith, D.; Peay, K.G.; Kennedy, P.G. Parsing ecological signal from noise in next generation amplicon sequencing. New Phytol. 2015, 205, 1389–1393. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, D.B.; Nilsson, R.H.; Tedersoo, L.; Abarenkov, K.; Carlsen, T.; Kjøller, R.; Kõljalg, U.; Pennanen, T.; Rosendahl, S.; Stenlid, J.; et al. Fungal community analysis by high-throughput sequencing of amplified markers—A user’s guide. New Phytol. 2013, 199, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Paulson, J.N.; Stine, O.C.; Bravo, H.C.; Pop, M. Differential abundance analysis for microbial marker-gene surveys. Nat. Methods 2013, 10, 1200–1202. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Waste not, want not: Why rarefying microbiome data is inadmissable. PLoS Comput. Biol. 2014, 10, e1003531. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.; Xu, Z.Z.; Peddada, S.; Amir, A.; Bittinger, K.; Gonzalez, A.; Lozupone, C.; Zaneveld, J.R.; Vazquer-Baeza, Y.; Birminghan, A.; et al. Normalization and microbial differential abundance strategies depend upon data characteristics. Microbiome 2017, 5, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Caceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Cline, M.S.; Smoot, M.; Cerami, E.; Kuchinsky, A.; Landys, N.; Workman, C.; Christmas, R.; Avila-Campilo, I.; Creech, M.; Gross, B.; et al. Integration of biological networks and gene expression data using Cytoscape. Nat. Protoc. 2007, 2, 2366–2382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallander, H.; Ekblad, A.; Godbold, D.L.; Johnson, D.; Bahr, A.; Baldrian, P.; Bjork, R.G.; Kieliszewska-Rokicka, B.; Kjoller, R.; Kraigher, H.; et al. Evaluation of methods to estimate production, biomass and turnover of ectomycorrhizal mycelium in forests soils—A review. Soil Biol. Biochem. 2013, 57, 1034–1047. [Google Scholar] [CrossRef]
- Tedersoo, L.; Mett, M.; Ishida, T.A.; Bahram, M. Phylogenetic relationships among host plants explain differences in fungal species richness and community composition in ectomycorrhizal symbiosis. New Phytol. 2013, 199, 822–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, T.A.; Nara, K.; Hogestu, T. Host effects on ectomycorrhizal fungal communities: Insight from eight host species in mixed conifer-broadleaf forests. New Phytol. 2007, 174, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Karst, J.; Burns, C.; Cale, J.A.; Antunes, P.M.; Woods, M.; Lamit, L.J.; Hoeksema, J.D.; Zabinski, C.; Gehring, C.A.; La Flèche, M.; et al. Tree species with limited geographical ranges show extreme responses to ectomycorrhizas. Glob. Ecol. Biogeogr. 2018, 27, 839–848. [Google Scholar] [CrossRef]
- Tedersoo, L.; Nara, K. General latitudinal gradient of biodiversity is reversed in ectomycorrhizal fungi. New Phytol. 2010, 185, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Bruns, T.D.; Bidartondo, M.I.; Taylor, D.L. Host specificity in ectomycorrhizal communities: What do the exceptions tell us? Integr. Comp. Biol. 2002, 42, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.G.; Izzo, A.D.; Bruns, T.D. There is high potential for the formation of common mycorrhizal networks between understory and canopy trees in a mixed evergreen forest. J. Ecol. 2003, 91, 1071–1080. [Google Scholar] [CrossRef]
- Motiejūnatie, J.; Kačergius, A.; Kasparavičius, J. Preliminary studies on genetic diversity of ectomycorrhizal fungus Suillus bovinus Lithuania. Botnaica Lithuanica 2013, 19, 91–98. [Google Scholar] [CrossRef]
- Kretzer, A.; Li, Y.; Szaro, T.; Bruns, T.D. Internal transcribed spacer sequences from 38 recognized species of Suillus sensu lato: Phylogenetic and taxonomic implications. Mycologia 1996, 88, 776–785. [Google Scholar] [CrossRef]
- Molina, R.; Massicotte, H.; Trappe, J.M. Specificity phenomena in mycorrhizal symbioses: Community-ecological consequences and practical implications. In Mycorrhizal Functioning; Allen, M.F., Ed.; Chapman and Hall: New York, NY, USA, 1992; pp. 357–423. [Google Scholar]
- Nara, K.; Nakaya, H.; Wu, B.; Zhou, Z.; Hogetsu, T. Underground primary succession of ectomycorrhizal fungi in a volcanic desert on Mount Fuji. New Phytol. 2003, 159, 743–756. [Google Scholar] [CrossRef] [Green Version]
- Rusca, T.A.; Kennedy, P.G.; Bruns, T.D. The effect of different pine hosts on the sampling of Rhizopogon spore banks in five Eastern Sierra Nevada forests. New Phytol. 2006, 170, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Hynson, N.A.; Bruns, T.D. Stayin’ alive: Survival of mycorrhizal fungal propagules from a 6-yr-old forest soil. Fungal Ecol. 2012, 5, 741–746. [Google Scholar] [CrossRef]
- Peay, K.G.; Garbelotto, M.; Bruns, T.D. Evidence of dispersal limitation in soil microorganisms: Isolation reduces species richness on mycorrhizal tree islands. Ecology 2010, 91, 3631–3640. [Google Scholar] [CrossRef] [PubMed]
- Bahram, M.; Põlme, S.; Kõljalg, U.; Zarre, S.; Tedersoo, L. Regional and local patterns of ectomycorrhizal fungal diversity and community structure along an altitudinal gradient in the Hyrcanian forests of northern Iran. New Phytol. 2012, 193, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Sujia, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [PubMed]
- Avis, P.G.; Mueller, G.M.; Lussenhop, J. Ectomycorrhizal fungal communities in two North American oak forests respond to nitrogen addition. New Phytol. 2008, 179, 472–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, F.; Barsoum, N.; Lilleskovm, E.; Bidartondo, M.I. Nitrogen availability is a primary determinant of conifer mycorrhizas across complex environmental gradients. Ecol. Lett. 2010, 13, 1103–1113. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.D.; Phillips, L.A.; Treu, R.; Ward, V.; Berch, S.M. Functional responses of ectomycorrhizal fungal communities to long-term fertilization of lodgepole pine (Pinus contorta Dougl. ex loud. var. latifoliaengelm.) stands in central British Columbia. Appl. Soil Ecol. 2012, 60, 29–40. [Google Scholar] [CrossRef]
- Erlandson, S.R.; Savage, J.A.; Cavender-Bares, J.M.; Peay, K.G. Soil moisture and chemistry influence diversity of ectomycorrhizal fungal communities associating with willow along an hydrologic gradient. FEMS Microbiol. Ecol. 2016, 92, fiv148. [Google Scholar] [CrossRef] [PubMed]
- Corrales, A.; Turner, B.L.; Tedersoo, L.; Anslan, S.; Dalling, J.W. Nitrogen addition alters ectomycorrhizal fungal communities and soil enzyme activities in a tropical montane forest. Fungal Ecol. 2017, 27, 14–23. [Google Scholar] [CrossRef]
- Jentschke, G.; Godbold, D. Metal toxicity and ectomycorrhizas. Physiol. Plant. 2000, 109, 107–116. [Google Scholar] [CrossRef]
- Rosenstock, N.P.; Berner, C.; Smits, M.M.; Kram, P.; Wallander, H. The role of phosphorus, magnesium and potassium availability in soil fungal exploration of mineral nutrient sources in Norway spruce forests. New Phytol. 2016, 211, 542–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrino-Kyker, S.R.; Kluber, L.A.; Petersen, S.M.; Coyle, K.P.; Hewins, C.R.; de Forest, J.L.; Smemo, K.A.; Burke, D.J. Mycorrhizal fungal communities respond to experimental elevation of soil pH and P availability in temperate hardwood forests. FEMS Microbiol. Ecol. 2016, 92, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Peay, K.G.; Kennedy, P.G.; Bruns, T.D. Rethinking ectomycorrhizal succession: Are root density and hyphal exploration types drivers of spatial and temporal zonation? Fungal Ecol. 2011, 4, 233–240. [Google Scholar] [CrossRef]
- Peay, K.G.; Bruns, T.D.; Kennedy, P.G.; Bergemann, S.E.; Garbelotto, M. A strong species-area relationship for eukaryotic soil microbes: Island size matters for ectomycorrhizal fungi. Ecol. Lett. 2007, 10, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Lilleskov, E.A.; Bruns, T.D.; Horton, T.R.; Taylor, D.L.; Grogan, P. Detection of forest stand-level spatial structure in ectomycorrhizal fungal communities. FEMS Microbiol. Ecol. 2004, 49, 319–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, S.; Sato, H.; Tanabe, A.S.; Hidaka, A.; Kadowaki, K.; Toju, H. Spatial segregation and aggregation of ectomycorrhizal and root-endophytic fungi in the seedlings of two Quercus species. PLoS ONE 2014, 9, e96363. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.E.; Henkel, T.W.; Williams, G.C.; Aime, M.C.; Fremier, A.K.; Vilgalys, R. Investigating niche partitioning of ectomycorrhizal fungi in specialized rooting zones of the monodominant leguminous tree Dicymbe corymbosa. New Phytol. 2017, 215, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Moeller, H.V.; Peay, K.G. Competition-function tradeoffs in ectomycorrhizal fungi. PeerJ 2016, 4, e2270. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
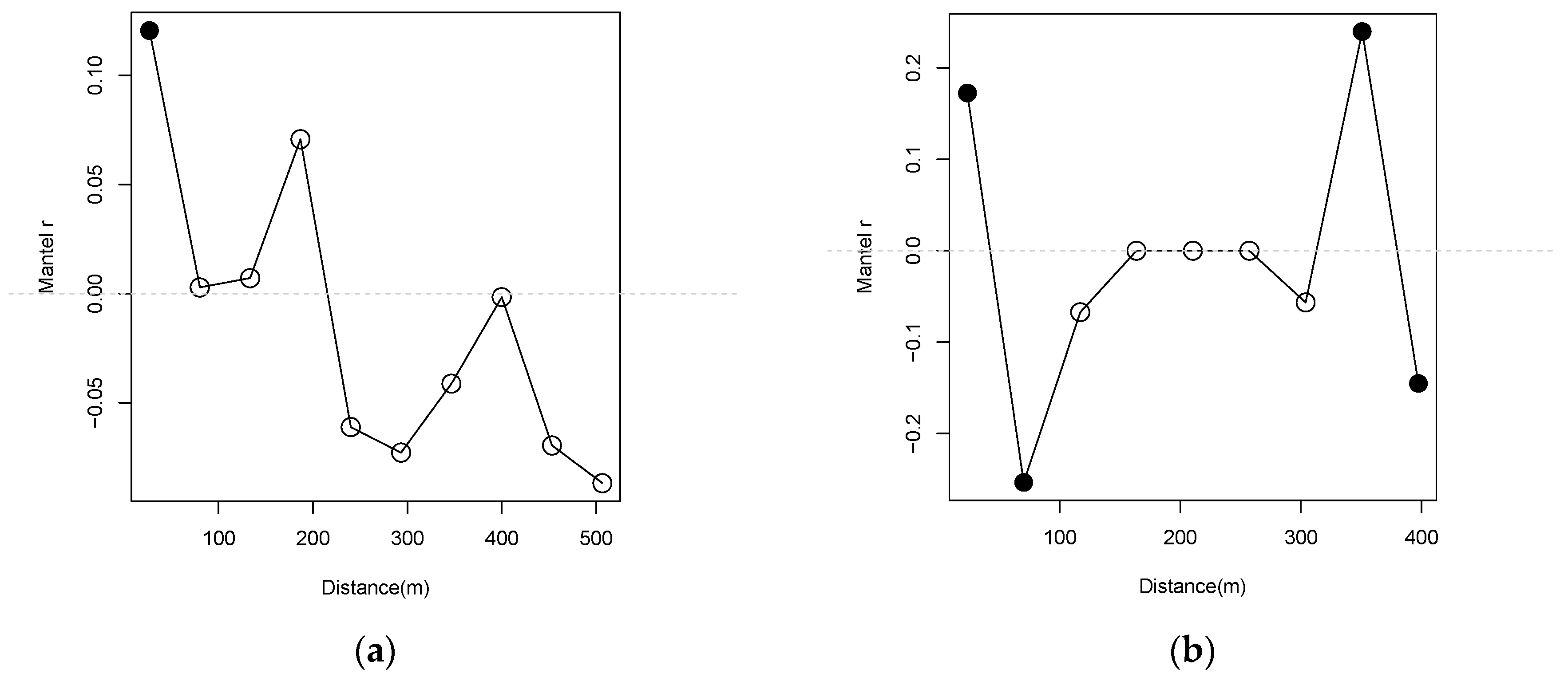{kind=link}
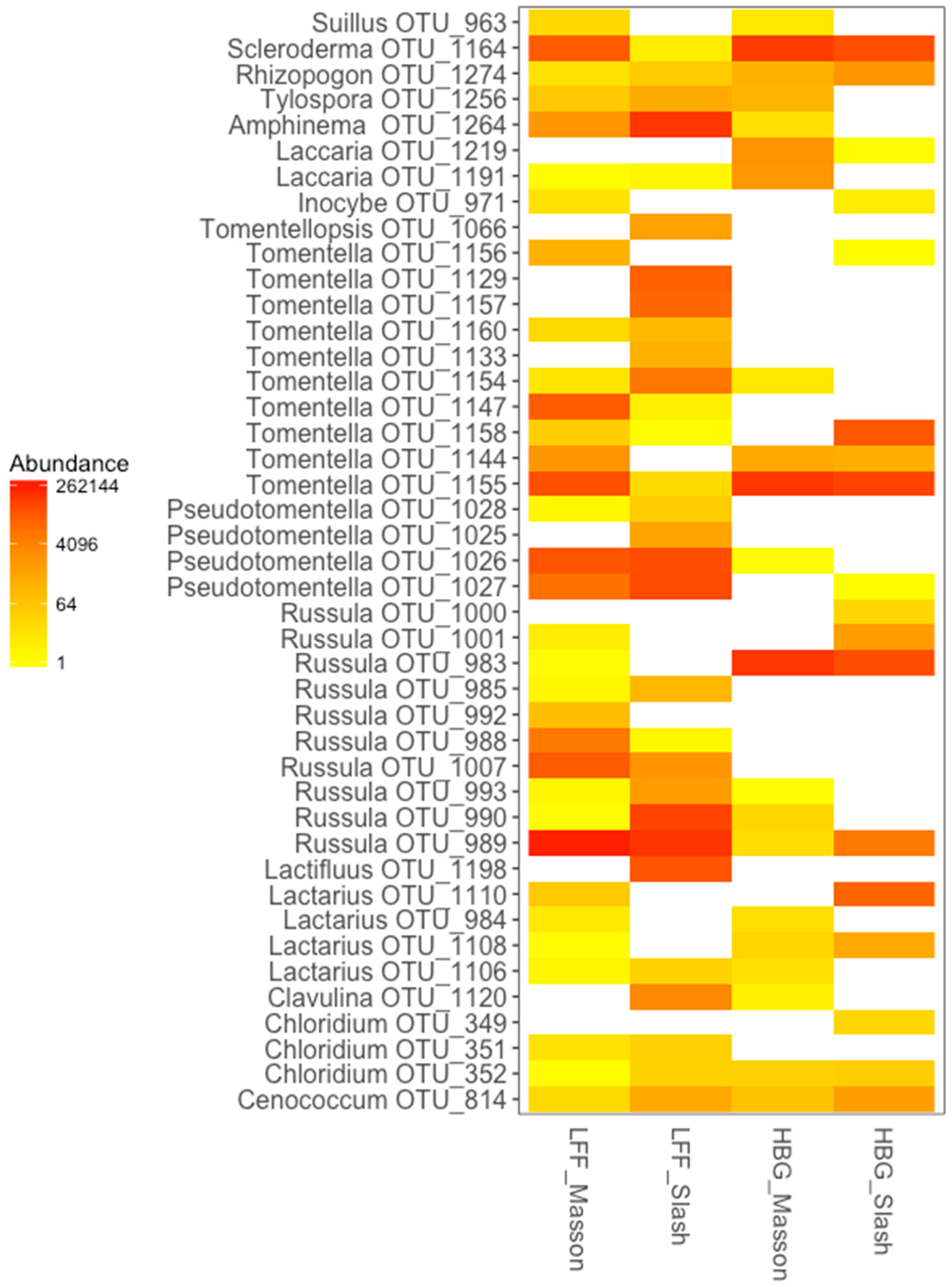
| Site | Species Name | OTU No. | Indicate Value | p-Value | Closest BLAST Match in Genbank | Closest UNITE Species Hypothesis Match | Distribution 1 | Host of Pinus |
|---|---|---|---|---|---|---|---|---|
| Longli Forest Farm, LFF | ||||||||
| MA | Russula virescens | OTU_988 | 0.683 | 0.003 | KU552087 (100) | SH179772.07FU | China | not indicated |
| Russula violeipes | OTU_1007 | 0.619 | 0.012 | LT201954 (100) | SH191296.07FU | Yunnan (China) | P. yunnanensis | |
| Russula.sp1 | OTU_992 | 0.447 | 0.05 | LT602950 (97) | SH218430.07FU | Yunnan (China) | P. yunnanensis | |
| SL | Amphiema.sp | OTU_1264 | 0.961 | 0.001 | LC176645 (100) | SH193510.07FU | China | P. massoniana |
| Tomentella stuposa | OTU_1154 | 0.856 | 0.001 | UDB024437 (100) | SH529807.07FU | Laos | not indicated | |
| Clavulina corralloides | OTU_1120 | 0.816 | 0.001 | KF359593 (94) | SH220215.07FU | North America | P. banksiana | |
| Russula sp.2 | OTU_990 | 0.774 | 0.001 | AB636419 (93) | SH218466.07FU | Korea | P. koraiensis | |
| Russula sp.3 | OTU_993 | 0.682 | 0.001 | AB211253 (95) | SH186553.07FU | Japan | P. densiflora | |
| Tomentella sp.1 | OTU_1157 | 0.632 | 0.001 | UDB018462 (97) | SH010050.07FU | China | not indicated | |
| Tomentella sp.2 | OTU_1129 | 0.632 | 0.002 | UDB018462 (99) | SH010050.07FU | China | not indicated | |
| Pseudotomentella sp1 | OTU_1027 | 0.628 | 0.011 | AB839386 (100) | SH223400.07FU | East Europe | P. sylvestris | |
| Pseudotomentella sp2 | OTU_1025 | 0.577 | 0.011 | AB587791 (100) | SH189639.07FU | China | P. massoniana | |
| Tomentella.sp.3 | OTU_1133 | 0.516 | 0.011 | AB587791 (100) | SH189639.07FU | Japan, Korea | P. thunbergii | |
| Site | Russula sp.4 | OTU_989 | 0.941 | 0.001 | AB211253 (99) | SH186553.07FU | Japan | not indicated |
| Pseudotomentella sp.3 | OTU_1026 | 0.913 | 0.001 | AB839386 (100) | SH223400.07FU | East Europe | P. sylvestris | |
| Hunan Botanic Garden, HBG | ||||||||
| MA | Russula sp.5 | OTU_983 | 0.785 | 0.001 | AB839393 (100) | SH201481.07FU | South China | P. massoniana |
| Laccaria amethystina | OTU_1219 | 0.548 | 0.015 | KF692988 (100) | SH220964.07FU | China, Japan | P. densiflora | |
| Laccaria aurantia | OTU_1191 | 0.547 | 0.03 | KU685645 (99) | SH179274.07FU | Yunnan (China) | P. yunnanensis | |
| SL | Russula sp.6 | OTU_1000 | 0.577 | 0.001 | UDB032527 (100) | SH189355.07FU | Laos | not indicated |
| Tomentella.sp.4 | OTU_1158 | 0.577 | 0.044 | AB587783 (100) | SH189355.07FU | a.w. | multiple | |
| Russula sp.7 | OTU_1001 | 0.576 | 0.006 | KP866130 (99) | SH199912.07FU | Hunan (China) | P. elliottii | |
| Site | Tomentella sp.5 | OTU_1155 | 0.891 | 0.003 | JX556209 (100) | SH189353.07FU | South China | not indicated |
| Scleroderma sp.1 | OTU_1164 | 0.772 | 0.006 | KP866131 (100) | SH189277.07FU | Hunan (China) | P. elliottii | |
| Analysis of Variance (ANOVA) Test | Mean (Standard Error) of Index | ||||||
|---|---|---|---|---|---|---|---|
| Longli Forest Farm (LFF) | Hunan Botanic Garden (HBG) | ||||||
| Masson Pine (n = 15) | Slash Pine (n = 15) | Masson Pine (n = 10) | Slash Pine (n = 15) | ||||
| Observed Richness | F | P | |||||
| Host | 2.11 | 0.152 | 7.07 (0.62) b | 10.33 (0.8) a | 7.50 (1) ab | 6.27 (0.53) b | |
| Site | 7.95 | 0.007 | |||||
| Interaction | 9.31 | 0.004 | |||||
| Chao1 | F | P | |||||
| Host | 0.43 | 0.513 | 8.13 (0.88) ab | 11.32 (0.8) a | 8.85 (1.6) ab | 6.77 (0.71) b | |
| Site | 5.36 | 0.025 | |||||
| Interaction | 7.51 | 0.008 | |||||
| Shannon | F | P | 0.47 (0.09) b | 0.72 (0.1) a | 0.30 (0.09) b | 0.58 (0.12) a | |
| Host | 5.49 | 0.023 | |||||
| Site | 2.93 | 0.174 | |||||
| Interaction | 0.02 | 0.899 | |||||
| PERMANOVA Test | PERMDISP Pair-Wise Test | |||||||
|---|---|---|---|---|---|---|---|---|
| Longli Forest Farm (LFF) | Hunan Botanic Garden (HBG) | |||||||
| Masson Pine | Slash Pine | Masson Pine | Slash Pine | |||||
| F | P | R2 | ||||||
| ECM fungal community | Host | 3.89 | 0.002 | 0.057 | 0.42 (0.05) c | 0.57 (0.03) b | 0.51 (0.03) ab | 0.64 (0.01) a |
| Site | 10.26 | 0.001 | 0.151 | |||||
| Interaction | 2.94 | 0.004 | 0.043 | |||||
| Soil nutrient status | F | P | R2 | |||||
| Host | 9.66 | 0.007 | 0.012 | 0.04 (0.01) | 0.05 (0.01) | 0.08 (0.02) | 0.04 (0.01) | |
| Site | 828.02 | 0.001 | 0.929 | |||||
| Interaction | 2.5 | 0.101 | 0.003 | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ning, C.; Mueller, G.M.; Egerton-Warburton, L.M.; Xiang, W.; Yan, W. Host Phylogenetic Relatedness and Soil Nutrients Shape Ectomycorrhizal Community Composition in Native and Exotic Pine Plantations. Forests 2019, 10, 263. https://doi.org/10.3390/f10030263
Ning C, Mueller GM, Egerton-Warburton LM, Xiang W, Yan W. Host Phylogenetic Relatedness and Soil Nutrients Shape Ectomycorrhizal Community Composition in Native and Exotic Pine Plantations. Forests. 2019; 10(3):263. https://doi.org/10.3390/f10030263
Chicago/Turabian StyleNing, Chen, Gregory M. Mueller, Louise M. Egerton-Warburton, Wenhua Xiang, and Wende Yan. 2019. "Host Phylogenetic Relatedness and Soil Nutrients Shape Ectomycorrhizal Community Composition in Native and Exotic Pine Plantations" Forests 10, no. 3: 263. https://doi.org/10.3390/f10030263
APA StyleNing, C., Mueller, G. M., Egerton-Warburton, L. M., Xiang, W., & Yan, W. (2019). Host Phylogenetic Relatedness and Soil Nutrients Shape Ectomycorrhizal Community Composition in Native and Exotic Pine Plantations. Forests, 10(3), 263. https://doi.org/10.3390/f10030263