The Colour of Tropical Woods Influenced by Brown Rot



Abstract
:1. Introduction
2. Materials and Methods
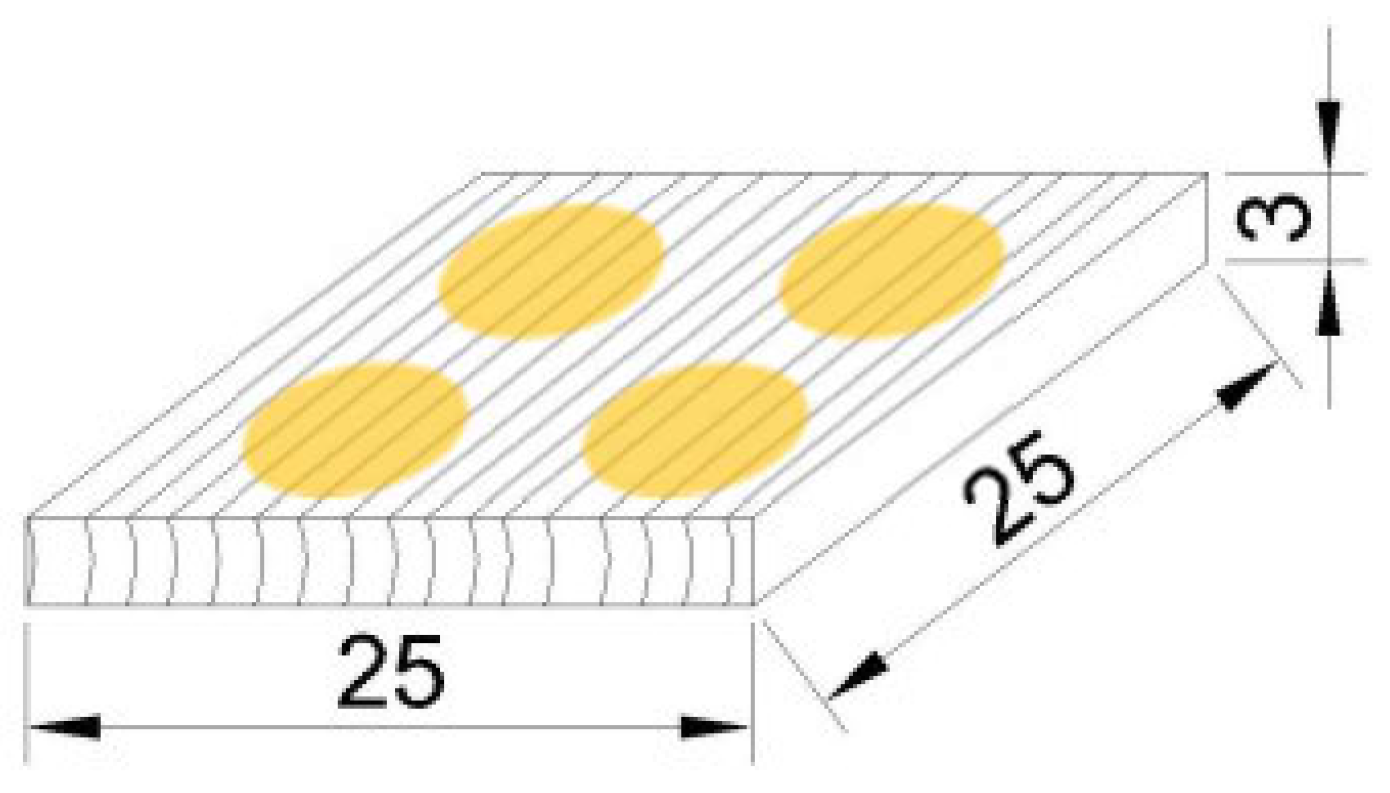
2.1. Woods and Specimens
2.2. Fungal Attack of Woods
2.3. Colour Analyses of Woods
3. Results and Discussion
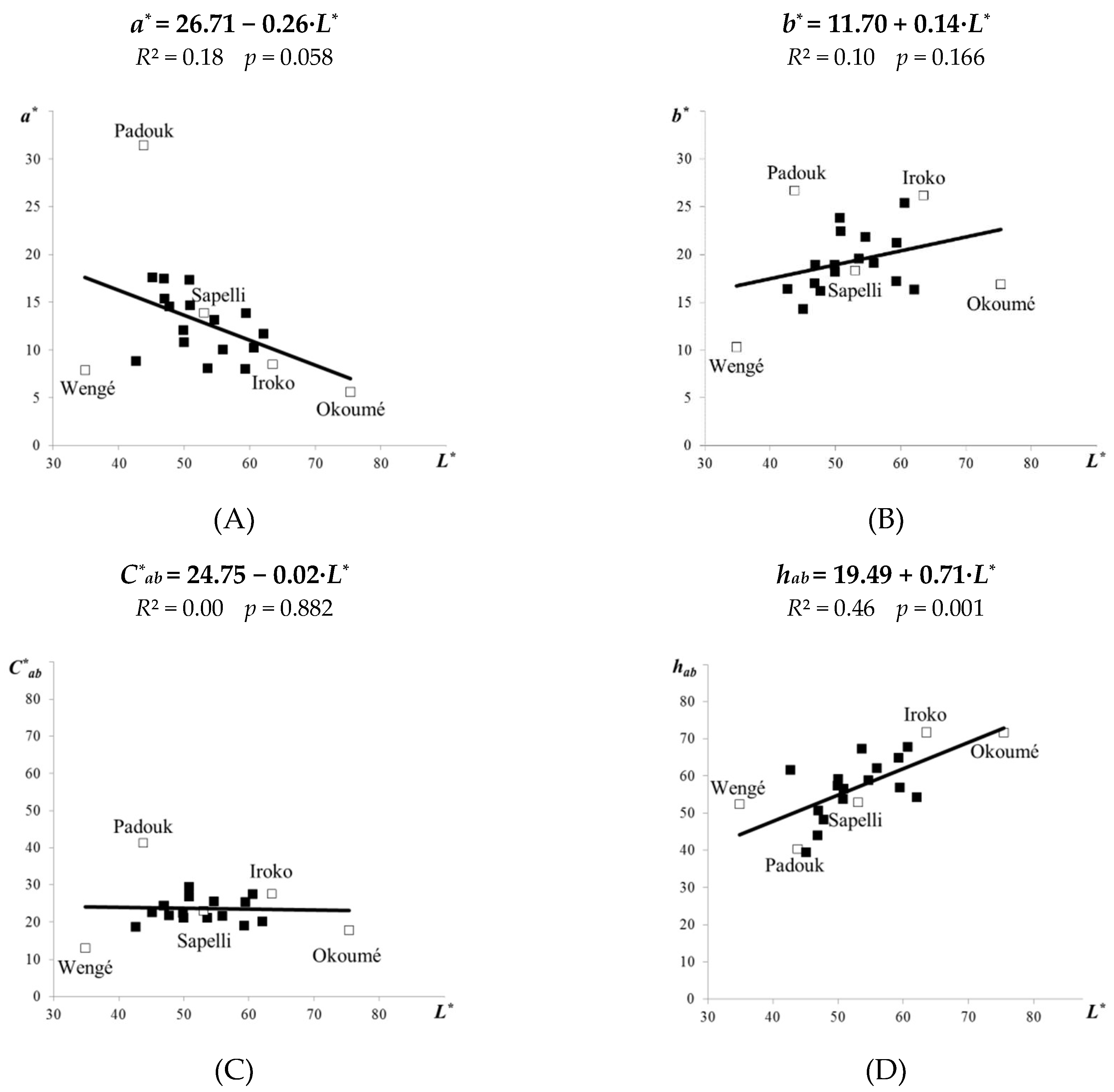
3.1. Colour of Original Tropical Woods
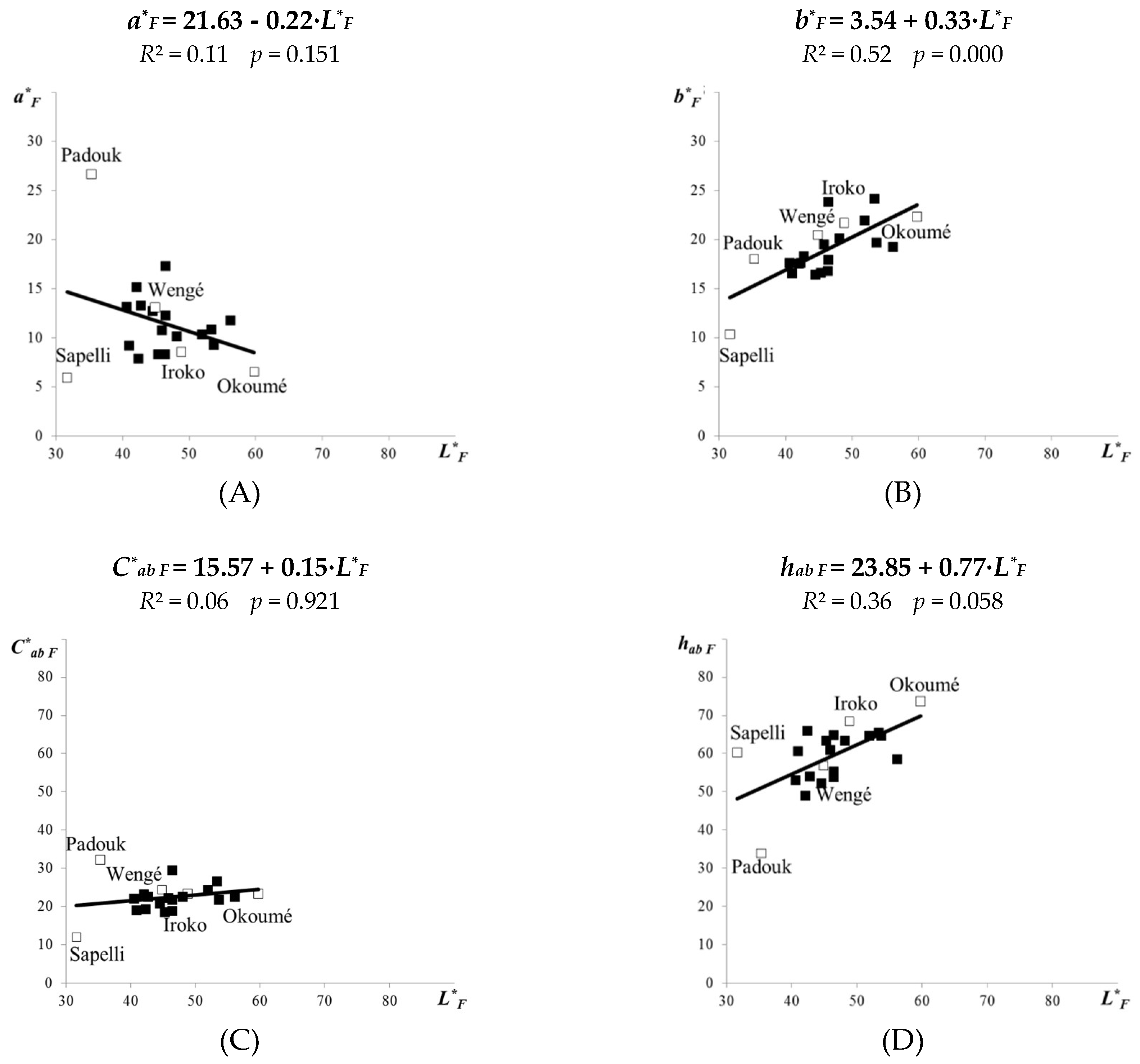
3.2. Colour of Tropical Woods Exposed to the Fungus Coniophora puteana
4. Conclusions
- For the original tropical woods, the a* coordinate (redding) declined with increase of the lightness L*, while the b* (yellowing) and h*ab (hue ton angle) coordinates grew with the lightness. No significance was found between C*ab (colour saturation) and L*.
- For the fungal-attacked tropical woods, the linear correlations between the colour coordinates a*, C*ab or hab and the lightness L* remained almost the same as for the original tropical woods, and only the b* coordinate grew more evidently in relation to L*.
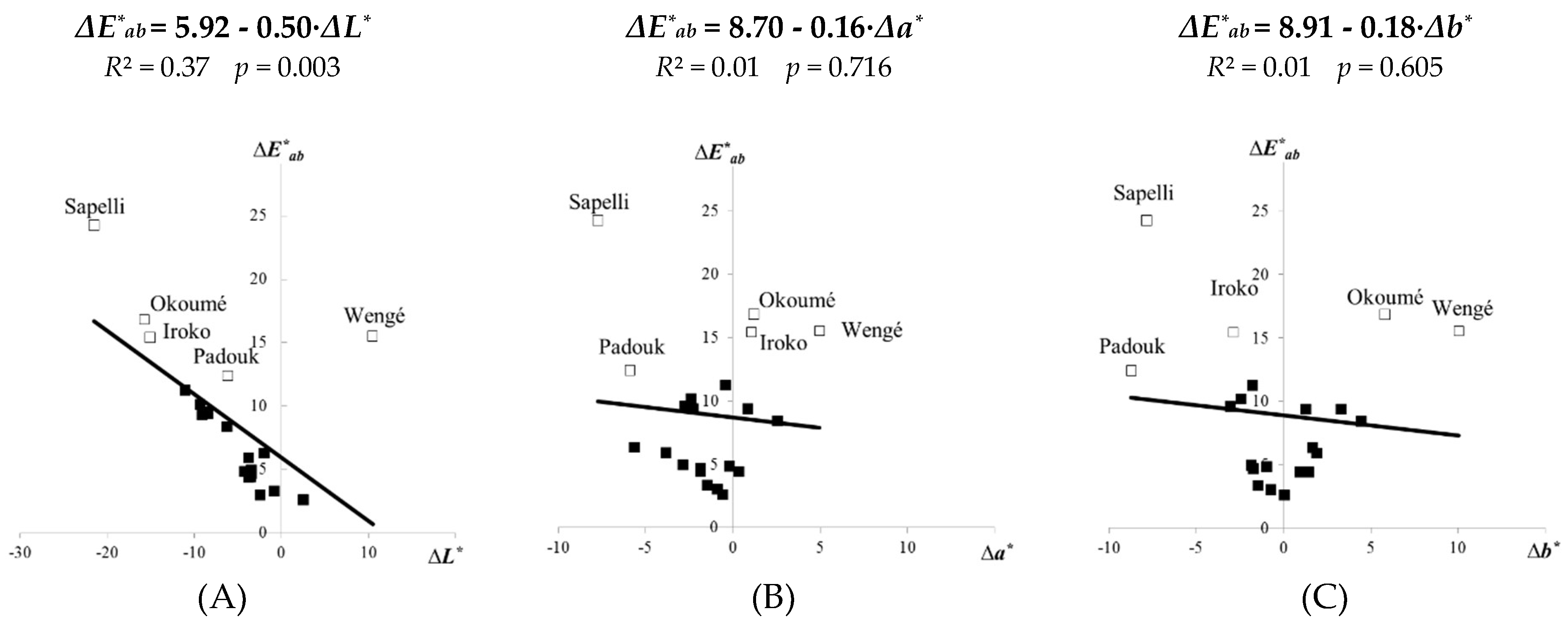
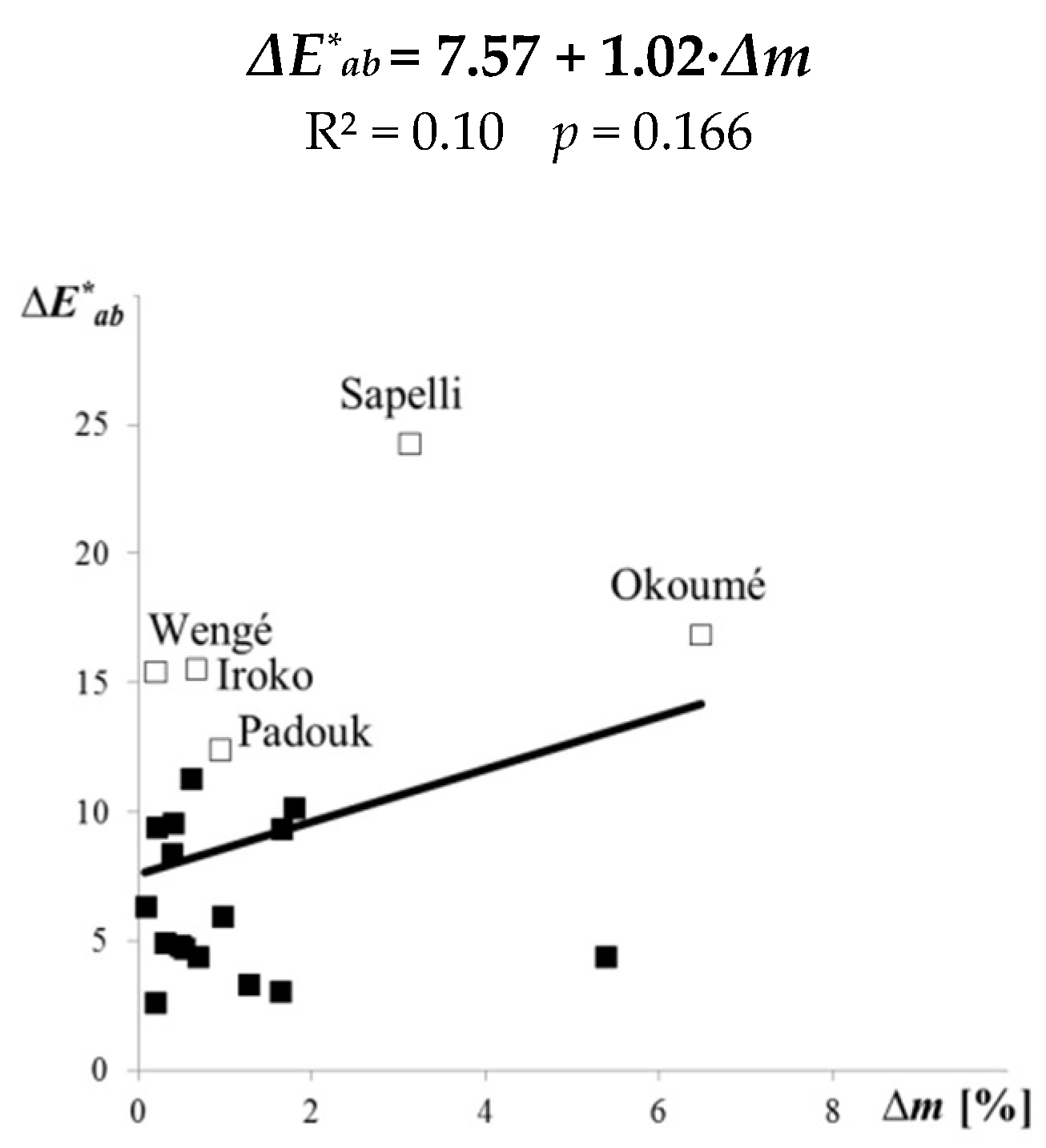
- The total discoloration ΔE∗ab values were highest for the top surfaces of the lighter species (sapelli, okoumé, iroko) and the darkest species (wengé), when the ΔE∗ab were justified by the marked change of the lightness ΔL*.
- Significant changes in the lightness and colouration of the fungal-attacked tropical woods indicated these colour changes could be caused not only by the biodegradation of polysaccharides, but also by biodegradation or leaching of some extractives during laboratory mycological tests.
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Janin, G.; Gonçalez, J.C.; Ananías, R.A.; Charrier, B.; Silva, G.F.D.; Dilem, A. Aesthetics appreciation of wood colour and patterns by colorimetry. Part 1. Colorimetry theory for the CIE Lab system. Maderas Cienc. Tecnol. 2001, 3, 14. [Google Scholar]
- Tolvaj, L.; Persze, L.; Lang, E. Correlation between hue angle and lightness of wood species grown in Hungary. Wood Res. 2013, 58, 141–145. [Google Scholar]
- Slabejová, G.; Šmidriaková, M.; Fekiač, J. Gloss of transparent coating on beech wood surface. Acta Fac. Xylologiae Zvolen 2016, 58, 37–44. [Google Scholar]
- Hon, D.N.-S.; Minemura, N. Color and discoloration. In Wood and Cellulosic Chemistry, 2nd ed.; Hon, D.N.-S., Shiraishi, N., Eds.; CRC Press: New York, USA, 2000; pp. 385–442. [Google Scholar]
- Babiak, M.; Kubovský, I.; Mamoňová, M. Color space of the selected domestic species. In Interaction of Wood with Various Forms of Energy, 1st ed.; Kurjatko, S., Kúdela, J., Eds.; Technical University in Zvolen: Zvolen, Slovakia, 2004; pp. 113–117. [Google Scholar]
- Katuščák, S.; Kucera, J. CIE orthogonal and cylindrical color parameters and the color sequences of the temperate wood species. Wood Res. 2000, 45, 9–21. [Google Scholar]
- da Silva, R.A.F.; Setter, C.; Mazette, S.S.; de Melo, R.R.; Stangerlin, D.M. Colorimetry of wood from thirty tropical species. Ciênc. Madeira 2017, 8, 36–41. [Google Scholar]
- Meints, T.; Teischinger, A.; Stingl, R.; Hansmann, C. Wood colour of central European wood species: CIE Lab characterisation and colour intensification. Eur. J. Wood Wood Prod. 2017, 75, 499–509. [Google Scholar] [CrossRef]
- Klement, I.; Vilkovská, T. Color characteristics of red false heartwood and mature wood of beech (Fagus sylvatica L.) determining by different colour saturation city coordinates. Sustainability 2019, 11, 690. [Google Scholar] [CrossRef]
- Mosedale, J.R.; Charrier, B.; Janin, G. Genetic control of wood colour, density and heartwood ellagitannin content of European oak (Quercus petraea and Quercus robur). Forestry 1996, 69, 111–124. [Google Scholar] [CrossRef]
- Phelps, J.E.; McGinnes, E.A.; Garret, H.E.; Cox, G.S. Growth quality evaluation of black walnut wood. II. Color analyses of veneer produced on different sites. Wood Fiber Sci. 1982, 15, 177–185. [Google Scholar]
- Derkyi, N.S.A.; Bailleres, H.; Chaix, G.; Thevenon, M.F. Colour variation in teak (Tectona grandis) wood from plantations across the ecological zones of Ghana. Ghana J. For. 2009, 25, 40–48. [Google Scholar] [CrossRef]
- Kržišnik, D.; Lesar, B.; Thaler, N.; Humar, M. Influence of natural and artificial weathering on the colour change of different wood and wood-based materials. Forests 2018, 9, 488. [Google Scholar] [CrossRef]
- Reinprecht, L.; Mamoňová, M.; Pánek, M.; Kačík, F. The impact of natural and artificial weathering on the visual, colour and structural changes of seven tropical woods. Eur. J. Wood Wood Prod. 2018, 76, 175–190. [Google Scholar] [CrossRef]
- Reinprecht, L.; Hulla, M. Colour changes in beech wood modified with essential oils due to fungal and ageing-fungal attacks with Coniophora puteana. Drewno 2015, 58, 37–48. [Google Scholar]
- Vidholdová, Z.; Slabejová, G.; Polomský, J. Colour changes of Scots pine wood due to action of the white-rot fungus Trametes versicolor. In Protecting Trees and Wood, 1st ed.; Hlaváč, P., Vidholdová, Z., Eds.; Technical University in Zvolen: Zvolen, Slovakia, 2016; pp. 61–66. [Google Scholar]
- Van Court, R.C.; Robinson, S.C. Stimulating Production of Pigment-Type Secondary Metabolites from Soft Rotting Wood Decay Fungi (“Spalting” Fungi). In Advances in Biochemical Engineering/Biotechnology, 1st ed.; Springer Nature: Basel, Switzerland, 2019; p. 16. [Google Scholar]
- Rayner, A.D.; Boddy, L. Fungal Decomposition of Wood. Its Biology and Ecology; Antony Rowe Ltd.: Chippenham, UK, 1997; p. 587. [Google Scholar]
- Pandey, K.K.; Pitman, A.J. FTIR studies of the changes in wood chemistry following decay by brown-rot and white-rot fungi. Int. Biodeter. Biodegr. 2003, 52, 151–160. [Google Scholar] [CrossRef]
- Zabel, R.A.; Morrell, J.J. Wood Microbiology: Decay and Its Prevention; Academic Press: London, UK, 2012; p. 476. [Google Scholar]
- Nascimento, M.S.; Santana, A.L.B.D.; Maranhão, C.A.; Oliveira, L.S.; Bieber, L. Phenolic extractives and natural resistance of wood. In Biodegradation-Life of Science; Chamy, R., Rosenkranz, F., Eds.; InTech: Rijeka, Croatia, 2013; pp. 349–370. [Google Scholar]
- Sablík, P.; Giagli, K.; Pařil, P.; Baar, J.; Rademacher, P. Impact of extractive chemical compounds from durable wood species on fungal decay after impregnation of nondurable wood species. Eur. J. Wood Wood Prod. 2016, 74, 231–236. [Google Scholar] [CrossRef]
- Schmidt, O. Indoor wood-decay basidiomycetes: Damage, causal fungi, physiology, identification and characterization, prevention and control. Mycol. Prog. 2007, 6, 261–279. [Google Scholar] [CrossRef]
- Frankl, J. Wood-damaging fungi in truss structures of baroque churches. J. Perform. Constr. Facil. 2015, 29, 04014138. [Google Scholar] [CrossRef]
- Hyde, K.D.; Al-Hatmi, A.M.S.; Andersen, B.; Boekhout, T.; Buzina, W.; Dawson, T.L.; Eastwood, D.C.; Jones, E.B.G.; Hoog, S.; Kang, Y.; et al. The world’s ten most feared fungi. Fungal Divers. 2018, 93, 161–194. [Google Scholar] [CrossRef]
- Gabriel, J.; Švec, K. Occurrence of indoor wood decay basidiomycetes in Europe. Fungal Biol. Rev. 2017, 31, 212–217. [Google Scholar] [CrossRef]
- Irbe, I.; Andersone, I.; Andersons, B.; Noldt, G.; Dizhbite, T.; Kurnosova, N.; Nuopponen, M.; Stewart, D. Characterisation of the initial degradation stage of Scots pine (Pinus sylvestris L.) sapwood after attack by brow-rot fungus Coniophora puteana. Biodegradation 2011, 22, 719–728. [Google Scholar] [CrossRef]
- Robinson, S.C.; Laks, P.E. Wood species and culture age affect zone line production of Xylaria polymorpha. Open Mycol. J. 2010, 4, 18–21. [Google Scholar] [CrossRef]
- Robinson, S.C.; Tudor, D.; Cooper, P.A. Wood preference of spalting fungi in urban hardwood species. Int. Biodeter. Biodegr. 2011, 65, 1145–1149. [Google Scholar] [CrossRef]
- Robinson, S.C.; Tudor, D.; Cooper, P.A. Feasibility of using red pigment producing fungi to stain wood for decorative applications. Can. J. For. Res. 2011, 41, 1722–1728. [Google Scholar] [CrossRef]
- Robinson, S.C. Developing fungal pigments for “painting” vascular plants. Appl. Microbiol. Biotechnol. 2012, 93, 1389–1394. [Google Scholar] [CrossRef]
- Robinson, S.C.; Tudor, D.; Cooper, P.A. Utilizing pigment-producing fungi to add commercial value to American beech (Fagus grandifolia). Appl. Microbiol. Biotechnol. 2012, 93, 1041–1048. [Google Scholar] [CrossRef]
- Beck, H.G.; Freitas, S.; Weber, G.; Robinson, S.C.; Morrell, J.J. Resistance of fungal derived pigments to ultraviolet light exposure. In International Research Group in Wood Protection; IRG/WP: St. George, UT, USA, 2014. [Google Scholar]
- Vega Gutierrez, S.; Robinson, S.C. Microscopic analysis of pigments extracted from spalting fungi. J. Fungi 2017, 3, 15. [Google Scholar] [CrossRef]
- Blanchette, R.A. Screening wood decayed by white rot fungi for preferential lignin degradation. Appl. Environ. Microbiol. 1984, 48, 647–653. [Google Scholar]
- Stalpers, J.A.; Vlug, I. Confistulina, the anamorphs of Fistulina hepatica. Can. J. Bot. 1983, 61, 1660–1666. [Google Scholar] [CrossRef]
- Hillis, W.E. Heartwood and Tree Exudates; Springer: Berlin/Heidelberg, Germany, 1987; p. 267. [Google Scholar]
- Coulson, J. Wood in Construction—How to Avoid Costly Mistakes; John Wiley & Sons Ltd.: Chichester, UK, 2012; p. 208. [Google Scholar]
- EN 350. Durability of Wood and Wood-Based Products. Testing and Classification of the Durability to Biological Agents of Wood and Wood-Based Materials; European Committee for Standardization: Brussels, Belgium, 2016. [Google Scholar]
- Wagenführ, R. Holzatlas; Fachbuchverlag Leipzig, Carl Hanser Verlag: Munchen, Germany, 2007; p. 816. [Google Scholar]
- URL 1. The IUCN Red List of Threatened Species. Version 2017-2. Available online: www.iucnredlist.org (accessed on 16 November 2017).
- CIE. Colorimetry—Part 4: CIE 1976 L*a*b Colour Space; CIE DS 014-4.3/E:2007; CIE Central Bureau: Vienna, Austria, 2007. [Google Scholar]
- Nishino, Y.; Janin, G.; Chanson, B.; Détienne, P.; Gril, J.; Thibaut, B. Colorimetry of wood specimens from French Guiana. J. Wood Sci. 1998, 44, 3–8. [Google Scholar] [CrossRef]
- Németh, K. The colour of wood in CIE Lab system. Az Erdészeti és Faipari Egyetem Tudományos Közleményei 1982, 2, 125–135. [Google Scholar]
- Eriksson, K.-E.L.; Blanchett, R.A.; Ander, P. Microbial and Enzymatic Degradation of Wood and Wood Components; Springer Series in Wood Science: Berlin/Heidelberg, Germany; New York, NY, USA, 1990; p. 407. [Google Scholar]
- Gierlinger, N.; Jacques, D.; Grabner, M.; Wimmer, R.; Schwanninger, M.; Rozenberg, P.; Pâques, L.E. Colour of larch heartwood and relationships to extractives and brown-rot decay resistance. Trees 2004, 18, 102–108. [Google Scholar] [CrossRef]
- Kokutse, A.D.; Stokes, A.; Baillères, H.; Kokou, K.; Baudasse, C. Decay resistance of Togolese teak (Tectona grandis Lf) heartwood and relationship with colour. Trees 2006, 20, 219–223. [Google Scholar] [CrossRef]
- Costa, M.D.A.; Costa, A.F.D.; Pastore, T.C.M.; Braga, J.W.B.; Gonçalez, J.C. Characterization of wood decay by rot fungi using colorimetry and infrared spectroscopy. Ciência Florestal 2011, 21, 567–577. [Google Scholar]
- Amusant, N.; Fournier, M.; Beauchene, J. Colour and decay resistance and its relationships in Eperua grandiflora. J. Ann. For. Sci. 2008, 65, 1–6. [Google Scholar] [CrossRef]
- Stangerlin, D.M.; Costa, A.F.D.; Gonçalez, J.C.; Pastore, T.C.M.; Garlet, A. Monitoring of biodeterioration of three Amazonian wood species by the colorimetry technique. Acta Amazon. 2013, 43, 429–438. [Google Scholar] [CrossRef]
- Blanchette, R.A.; Wilmering, A.M.; Baumeister, M. The use of green-stained wood caused by the fungus Chlorociboria in intarsia masterpieces from the 15th century. Holzforschung 1992, 46, 225–232. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Species | Density at MC 12% (kg·m−3) | |
|---|---|---|---|---|
| Common Name 1) | Scientific Name | “Literature” [40] | “Experiment” | |
| Bignoniaceae | Ipé | Handroanthus serratifolius (Vahl) S.O.Grose 3) 4) | 960–1100 | 968 (26) |
| Burseraceae | Okoumé | Aucoumea klaineana Pierre 2) | 370–560 | 566 (27) |
| Cunoniaceae | Tineo | Weinmannia trichosperma Cav. | 570–650 | 646 (31) |
| Dipterocarpaceae | Dark red meranti | Shorea curtisii Dyer ex King 2) | 590–890 | 592 (10) |
| Yellow balau 5) | Shorea laevis Ridl. 2) | 900–1100 | 925 (55) | |
| Ebenaceae | Macassar ebony | Diospyros celebica Bakh. 2) | 1100–1200 | 1 013 (82) |
| Fabaceae | Doussié | Afzelia bipindensis Harms 2) | 750–950 | 889 (18) |
| Cerejeira | Amburana cearensis A. C. Sm. 2) | 550–650 | 651 (10) | |
| Bubinga | Guibourtia demeusei J. Léonard | 830–950 | 830 (13) | |
| Ovengol | Guibourtia ehie J. Léonard 2) | 700–910 | 755 (27) | |
| Merbau | Intsia bijuga O. Ktze. 2) | 830–900 | 837 (50) | |
| Santos rosewood | Machaerium scleroxylon Tul. 2) | 900–1000 | 904 (6) | |
| Zebrano | Microberlinia brazzavillensis Chev. 2) | 700–850 | 718 (23) | |
| Wengé | Millettia laurentii De Wild. 2) | 810–950 | 823 (37) | |
| Padouk | Pterocarpus soyauxii Taub. | 650–850 | 647 (37) | |
| Meliaceae | Sapelli | Entandrophragma cylindricum Sprague 2) | 510–750 | 631 (38) |
| Moraceae | Iroko | Milicia excelsa C. C. Berg 2) | 550–850 | 551 (16) |
| Myrtaceae | Karri | Eucalyptus diversicolor F. Muell. | 800–870 | 804 (30) |
| Blue gum | Eucalyptus globulus Labill. | 720–770 | 760 (59) | |
| Sapotaceae | Maçaranduba 6) | Manilkara bidenta A. Chev. | 900–1000 | 916 (19) |
| Makoré | Tieghemella heckelii Pierre 2) | 530–720 | 570 (25) | |
| Wood Species | Visual | Lightness L* | Redness +a* | Yellowness +b* | Colour Saturation C*ab | Hue Tone Angle hab |
|---|---|---|---|---|---|---|
| Ipé |  | 42.59 (0.79) | 8.87 (0.55) | 16.46 (1.01) | 18.70 (2.59) | 61.66 (1.22) |
| Okoumé |  | 75.36 (1.13) | 5.61 (0.61) | 16.89 (0.57) | 17.81 (0.65) | 71.66 (1.69) |
| Tineo |  | 47.69 (4.98) | 14.56 (1.81) | 16.23 (0.99) | 21.83 (1.77) | 48.26 (2.83) |
| Darkred meranti |  | 62.03 (1.52) | 11.76 (0.81) | 16.35 (0.64) | 20.15 (0.87) | 54.30 (1.56) |
| Yellow balau |  | 54.59 (0.92) | 13.22 (0.83) | 21.92 (0.88) | 25.60 (1.12) | 58.93 (1.02) |
| Macassar ebony |  | 59.40 (0.80) | 13.86 (0.68) | 21.29 (0.81) | 25.41 (0.91) | 56.93 (1.23) |
| Doussié |  | 60.57 (6.61) | 10.28 (1.89) | 25.42 (2.09) | 27.50 (1.81) | 67.93 (4.80) |
| Cerejeira |  | 49.93 (3.34) | 10.85 (1.52) | 18.19 (1.34) | 21.22 (1.59) | 59.24 (3.46) |
| Bubinga |  | 46.81 (1.23) | 17.50 (0.89) | 16.98 (1.17) | 24.40 (1.20) | 44.11 (2.00) |
| Ovengol |  | 53.56 (4.65) | 8.08 (1.07) | 19.63 (4.38) | 21.27 (4.33) | 67.36 (2.69) |
| Merbau |  | 46.92 (1.57) | 15.43 (0.53) | 18.97 (1.12) | 24.46 (1.09) | 50.83 (1.37) |
| Santos rosewood |  | 50.79 (3.39) | 14.68 (1.31) | 22.49 (3.13) | 26.88 (3.23) | 56.63 (2.44) |
| Zebrano |  | 55.85 (4.91) | 10.09 (1.06) | 19.18 (1.49) | 21.68 (1.63) | 62.24 (2.18) |
| Wengé |  | 34.88 (1.68) | 7.89 (0.64) | 10.33 (1.30) | 13.02 (1.24) | 52.42 (3.38) |
| Padouk |  | 43.78 (2.05) | 31.42 (1.61) | 26.72 (2.02) | 41.26 (2.32) | 40.34 (1.57) |
| Sapelli |  | 53.04 (1.69) | 13.88 (0.88) | 18.34 (0.72) | 23.01 (0.94) | 52.90 (1.62) |
| Iroko |  | 63.52 (6.78) | 8.49 (0.88) | 26.20 (5.69) | 27.57 (5.59) | 71.70 (2.54) |
| Karri |  | 50.72 (2.07) | 17.36 (1.13) | 23.84 (1.59) | 29.50 (1.83) | 53.92 (1.32) |
| Blue gum |  | 59.25 (3.37) | 8.05 (0.74) | 17.24 (1.23) | 19.03 (1.27) | 64.95 (1.98) |
| Maçaranduba |  | 45.06 (2.16) | 17.61 (2.39) | 14.34 (0.99) | 22.75 (2.23) | 39.45 (3.79) |
| Makoré |  | 49.85 (2.24) | 12.08 (0.88) | 18.95 (1.10) | 22.49 (1.15) | 57.47 (2.10) |
| Wood species | Δm | ΔL* | Δa* | Δb* | ΔC*ab | Δhab | ΔE*ab | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ipé | 0.20 (0.03) | 2.47 (1.32) | a | −0.59 (0.50) | c | 0.01 (0.31) | d | −0.26 (0.29) | d | 1.68 (1.56) | b | 2.63 (1.26) |
| Okoumé | 6.48 (2.98) | −15.74 (1.55) | a | 1.20 (0.23) | b | 5.80 (0.50) | a | 5.93 (0.49) | a | 1.70 (0.51) | d | 16.84 (1.34) |
| Tineo | 5.38 (3.71) | −3.59 (1.63) | d | −1.88 (1.12) | b | 1.41 (0.43) | c | −0.11 (0.92) | d | 6.03 (2.10) | a | 4.44 (1.68) |
| Dark red meranti | 0.22 (0.09) | −8.44 (0.96) | a | −2.29 (0.27) | a | 3.28 (1.04) | a | 1.71 (0.94) | a | 9.90 (1.70) | a | 9.43 (0.70) |
| Yellow balau | 0.40 (0.08) | −8.61 (1.06) | a | −2.77 (1.00) | a | −3.08 (0.95) | a | −4.04 (1.17) | a | 2.04 (1.77) | b | 9.59 (1.47) |
| Macassar ebony | 0.52 (0.30) | −3.48 (1.87) | a | −1.88 (0.31) | a | −1.76 (0.84) | a | −2.49 (0.86) | a | 1.57 (0.49) | a | 4.73 (0.81) |
| Doussié | 1.63 (1.51) | −2.51 (1.68) | d | −0.91 (0.76) | d | −0.76 (0.66) | d | −1.23 (0.84) | d | 1.36 (1.07) | d | 3.05 (1.60) |
| Cerejeira | 0.30 (0.15) | −3.51 (2.26) | b | −2.89 (1.57) | b | −1.86 (1.28) | b | −2.89 (1.67) | b | 5.73 (2.12) | c | 4.98 (2.93) |
| Bubinga | 0.96 (0.13) | −3.81 (1.24) | a | −3.87 (0.85) | a | 1.89 (0.47) | a | −1.11 (0.90) | c | 10.29 (1.10) | a | 5.96 (0.15) |
| Ovengol | 0.61 (1.02) | −11.08 (1.28) | a | −0.46 (0.48) | d | −1.81 (0.84) | a | −1.83 (0.87) | b | −0.84 (1.10) | d | 11.28 (1.24) |
| Merbau | 0.48 (0.32) | −4.29 (1.71) | a | −0.22 (1.71) | d | −1.01 (1.25) | d | −0.88 (1.95) | d | −1.16 (1.99) | d | 4.87 (1.78) |
| Santos rosewood | 1.28 (0.31) | −0.83 (0.66) | d | −1.49 (1.67) | c | −1.48 (2.55) | d | −2.49 (0.86) | d | 1.57 (0.49) | d | 3.36 (1.89) |
| Zebrano | 1.65 (0.62) | −9.14 (1.01) | a | 0.82 (1.04) | d | 1.26 (1.18) | d | 1.52 (1.46) | d | −0.09 (1.69) | d | 9.39 (1.08) |
| Wengé | 0.67 (0.35) | 10.46 (2.17) | a | 4.98 (0.72) | a | 10.05 (1.87) | a | 11.16 (1.50) | a | 4.69 (2.60) | b | 15.54 (1.55) |
| Padouk | 0.95 (1.69) | −6.18 (1.32) | a | −5.91 (0.96) | a | −8.74 (1.93) | a | −9.93 (1.46) | a | −5.49 (2.70) | a | 12.41 (1.41) |
| Sapelli | 3.12 (1.15) | −21.54 (0.92) | a | −7.76 (1.26) | a | −7.87 (1.52) | a | −10.84 (1.99) | a | 7.21 (1.11) | a | 24.26 (1.52) |
| Iroko | 0.20 (0.97) | −15.08 (3.76) | a | 1.07 (0.57) | c | −2.89 (1.41) | c | −2.37 (1.49) | d | −4.30 (0.71) | a | 15.44 (3.89) |
| Karri | 0.69 (0.47) | −3.83 (1.37) | a | 0.31 (1.23) | d | 0.89 (1.41) | d | 0.91 (1.81) | d | 0.54 (0.89) | d | 4.43 (1.17) |
| Blue gum | 0.38 (0.21) | −6.27 (2.54) | a | 2.53 (1.46) | a | 4.42 (0.88) | a | 5.15 (1.52) | a | −0.99 (1.35) | d | 8.42 (1.91) |
| Maçaranduba | 0.08 (0.05) | −1.99 (1.21) | b | −5.68 (0.40) | a | 1.66 (0.40) | b | −2.81 (0.38) | a | 13.46 (1.08) | a | 6.35 (0.64) |
| Makoré | 1.78 (1.84) | −9.43 (1.69) | a | −2.44 (1.37) | a | −2.45 (1.65) | b | −3.33 (2.09) | b | 2.35 (1.28) | c | 10.19 (2.10) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vidholdová, Z.; Reinprecht, L. The Colour of Tropical Woods Influenced by Brown Rot. Forests 2019, 10, 322. https://doi.org/10.3390/f10040322
Vidholdová Z, Reinprecht L. The Colour of Tropical Woods Influenced by Brown Rot. Forests. 2019; 10(4):322. https://doi.org/10.3390/f10040322
Chicago/Turabian StyleVidholdová, Zuzana, and Ladislav Reinprecht. 2019. "The Colour of Tropical Woods Influenced by Brown Rot" Forests 10, no. 4: 322. https://doi.org/10.3390/f10040322
APA StyleVidholdová, Z., & Reinprecht, L. (2019). The Colour of Tropical Woods Influenced by Brown Rot. Forests, 10(4), 322. https://doi.org/10.3390/f10040322