Site Type Effect on Litter Decomposition Rates: A Three-Year Comparison of Decomposition Process between Spoil Heap and Forest Sites



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Details
2.3. Statistical Analyses
3. Results
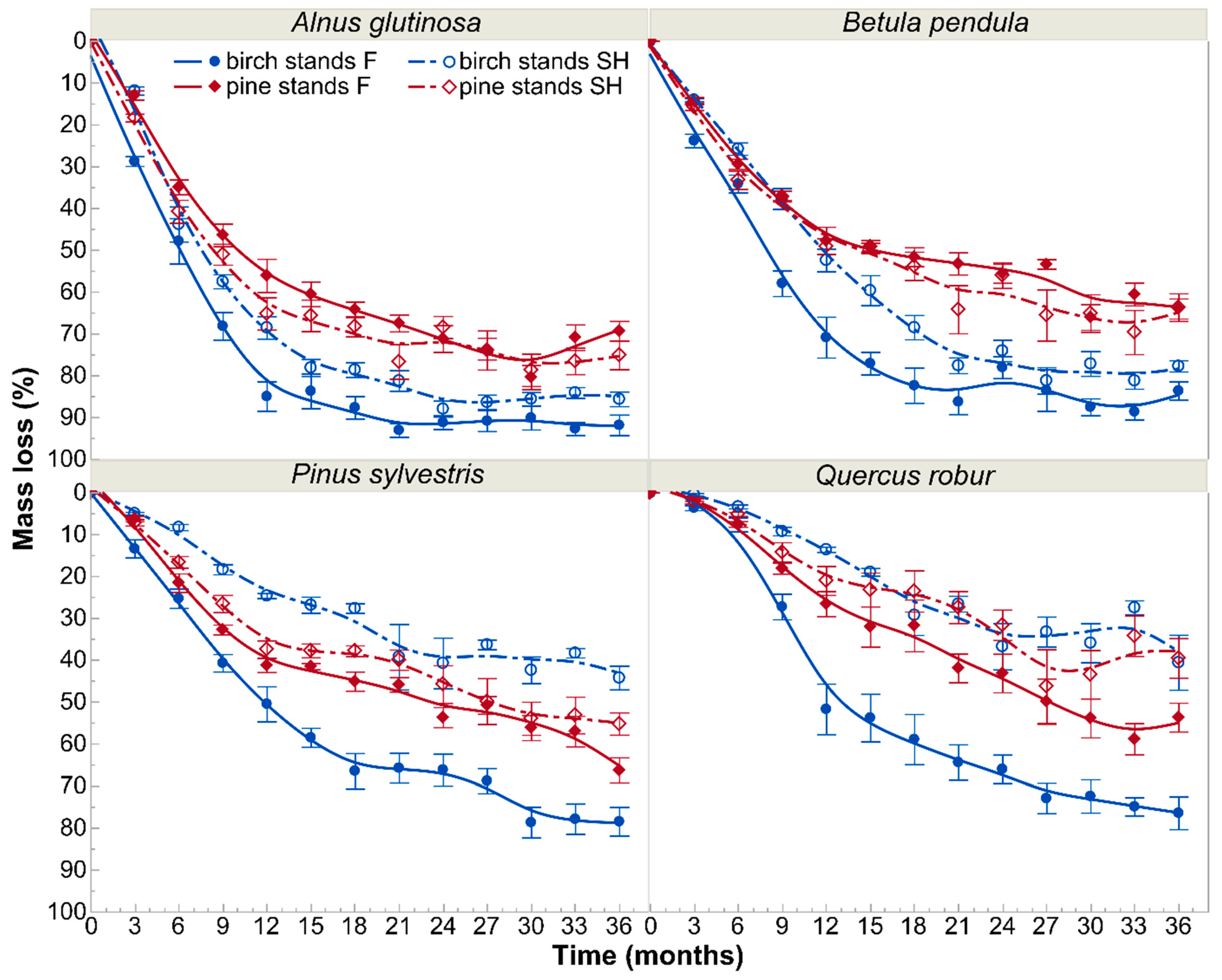
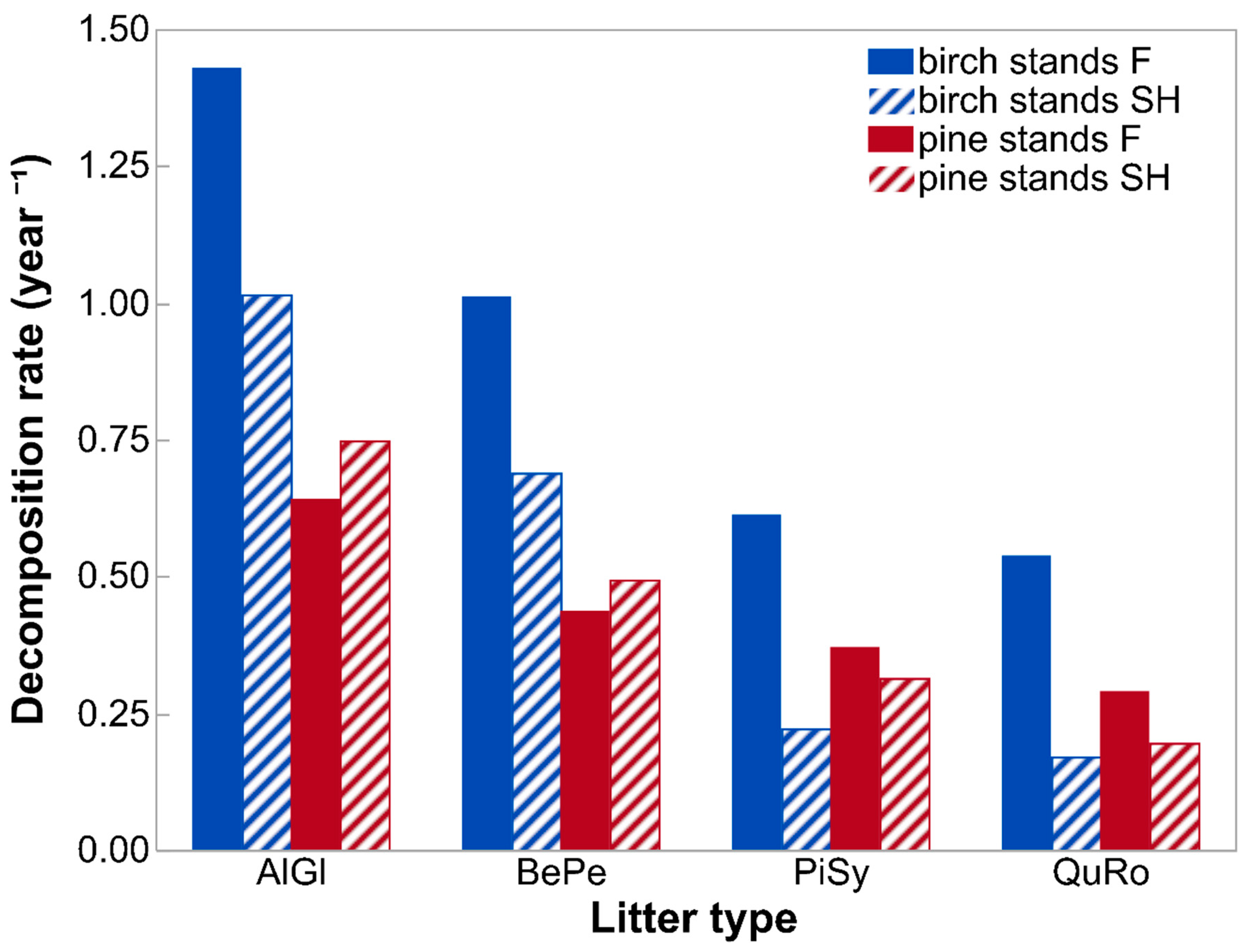
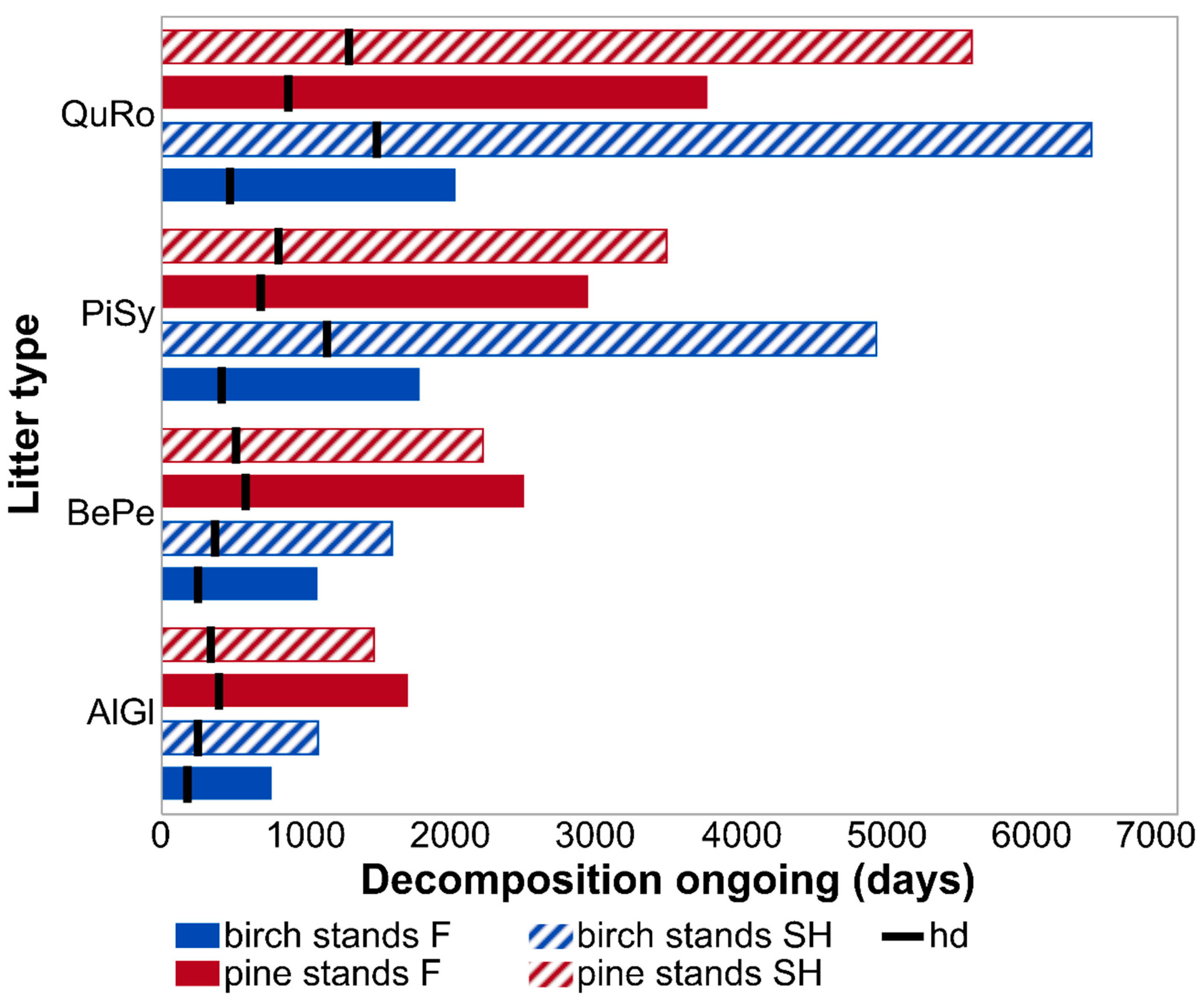
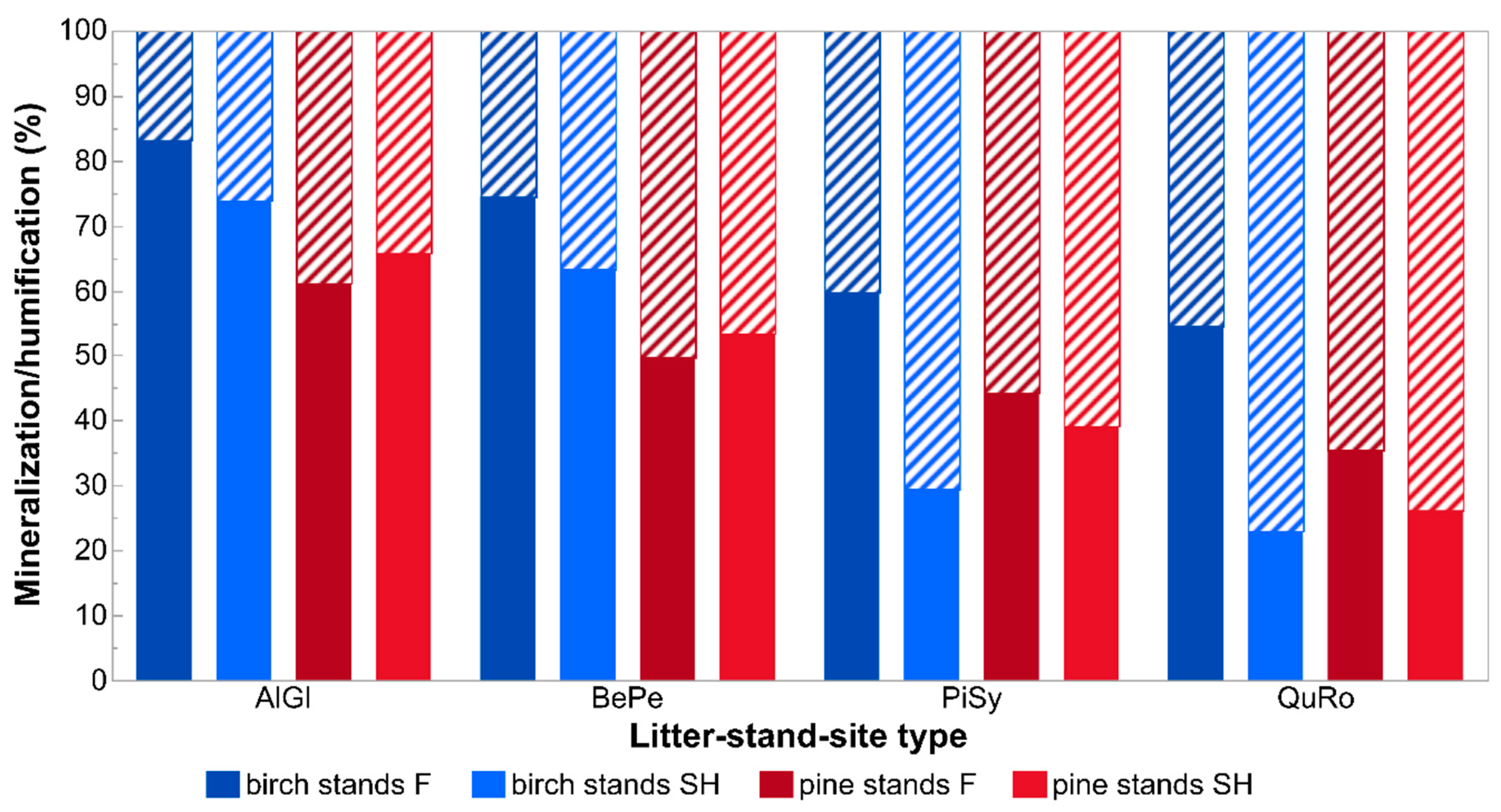
3.1. Decomposition
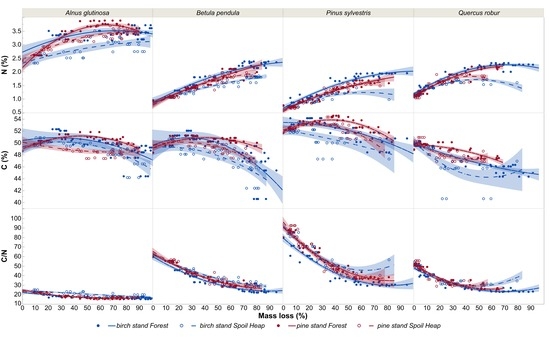
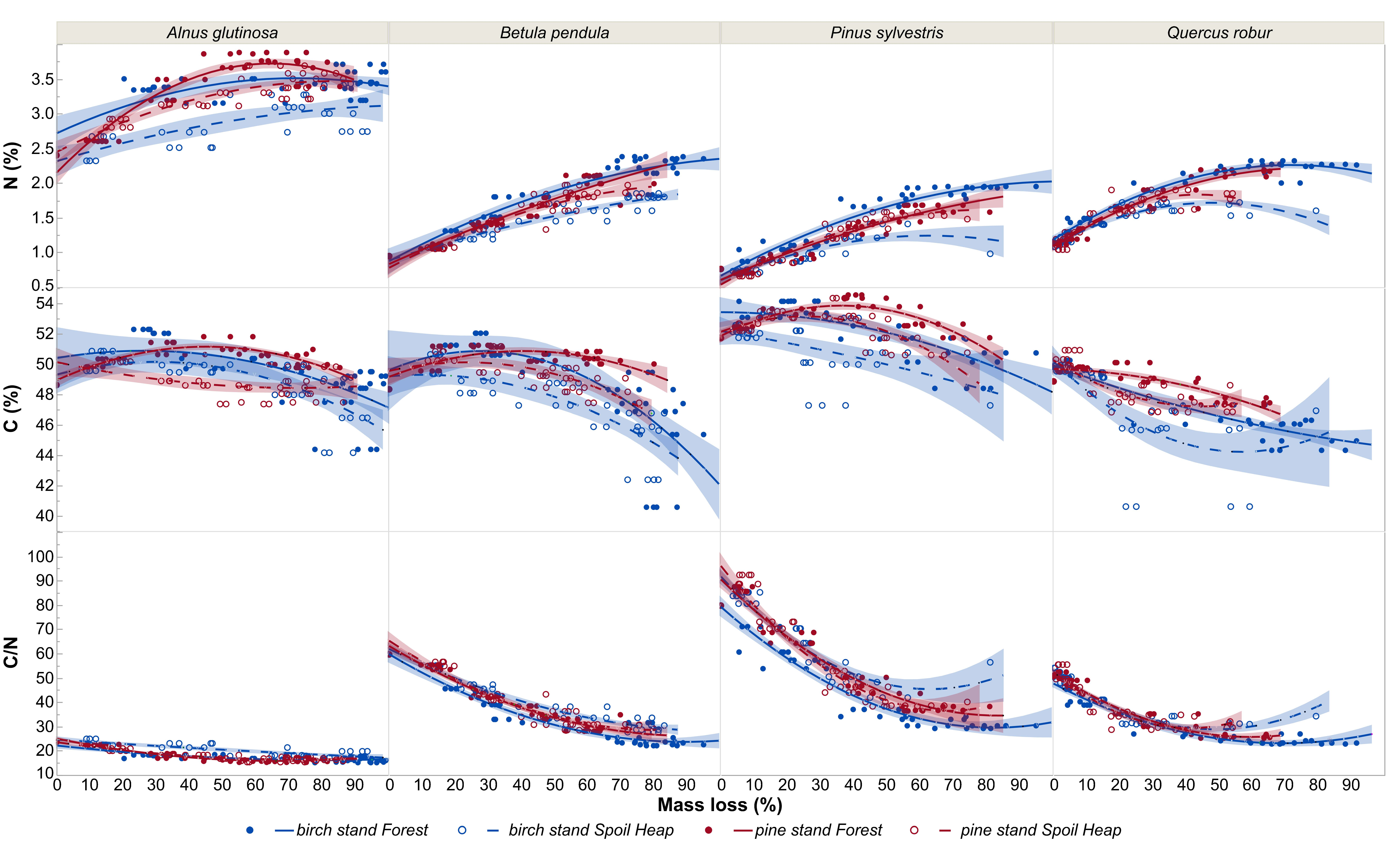
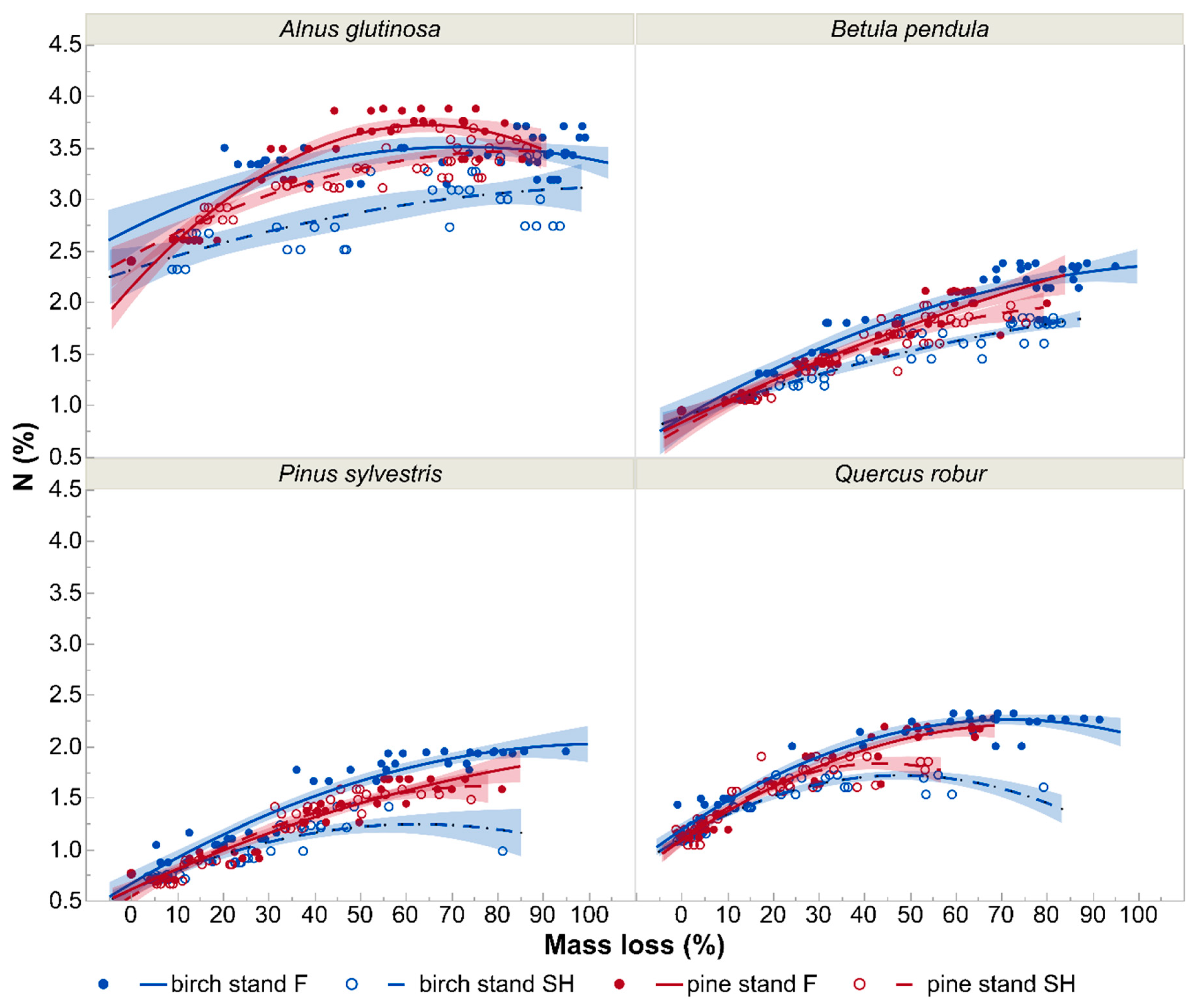
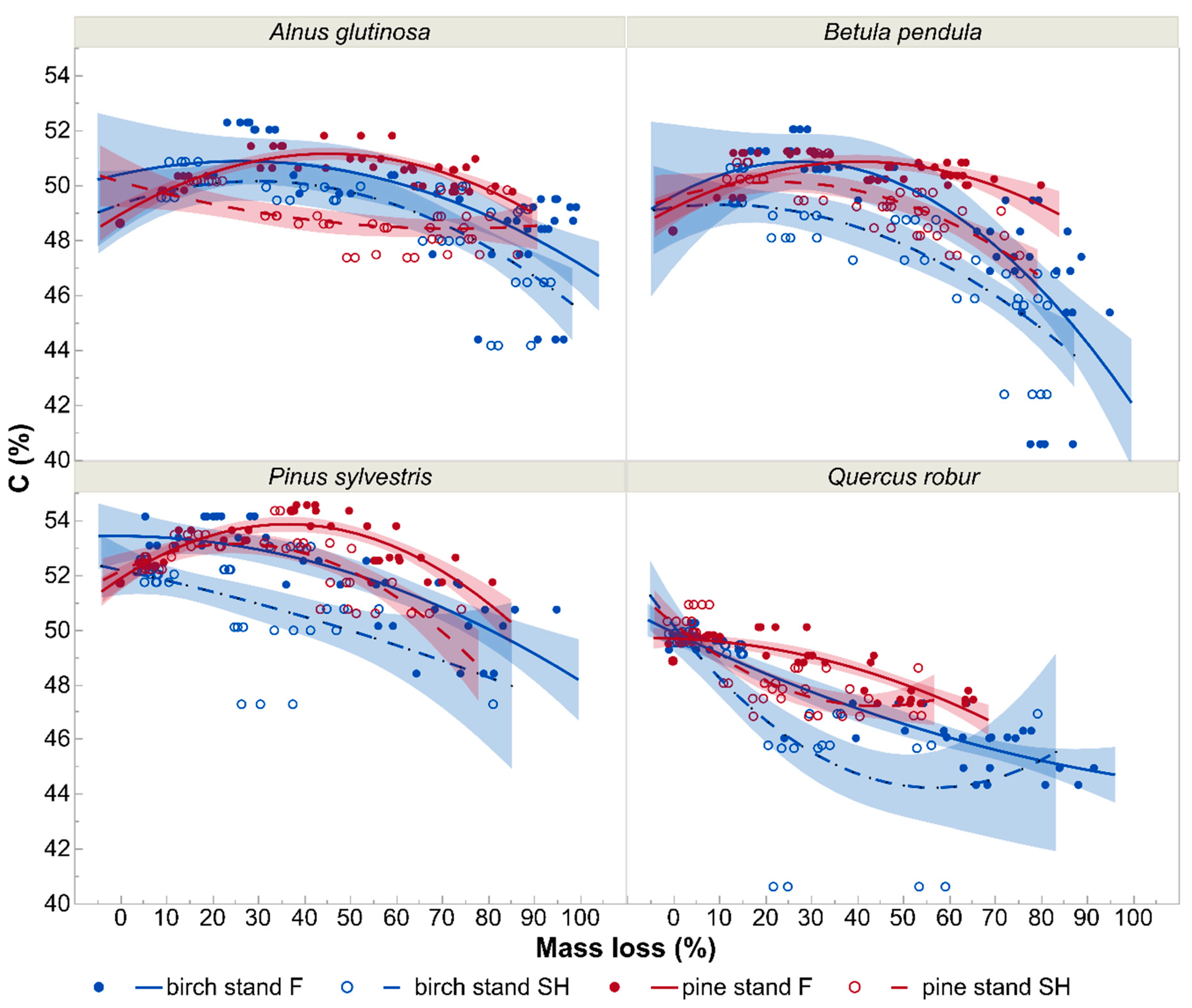
3.2. N and C Concentrations
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
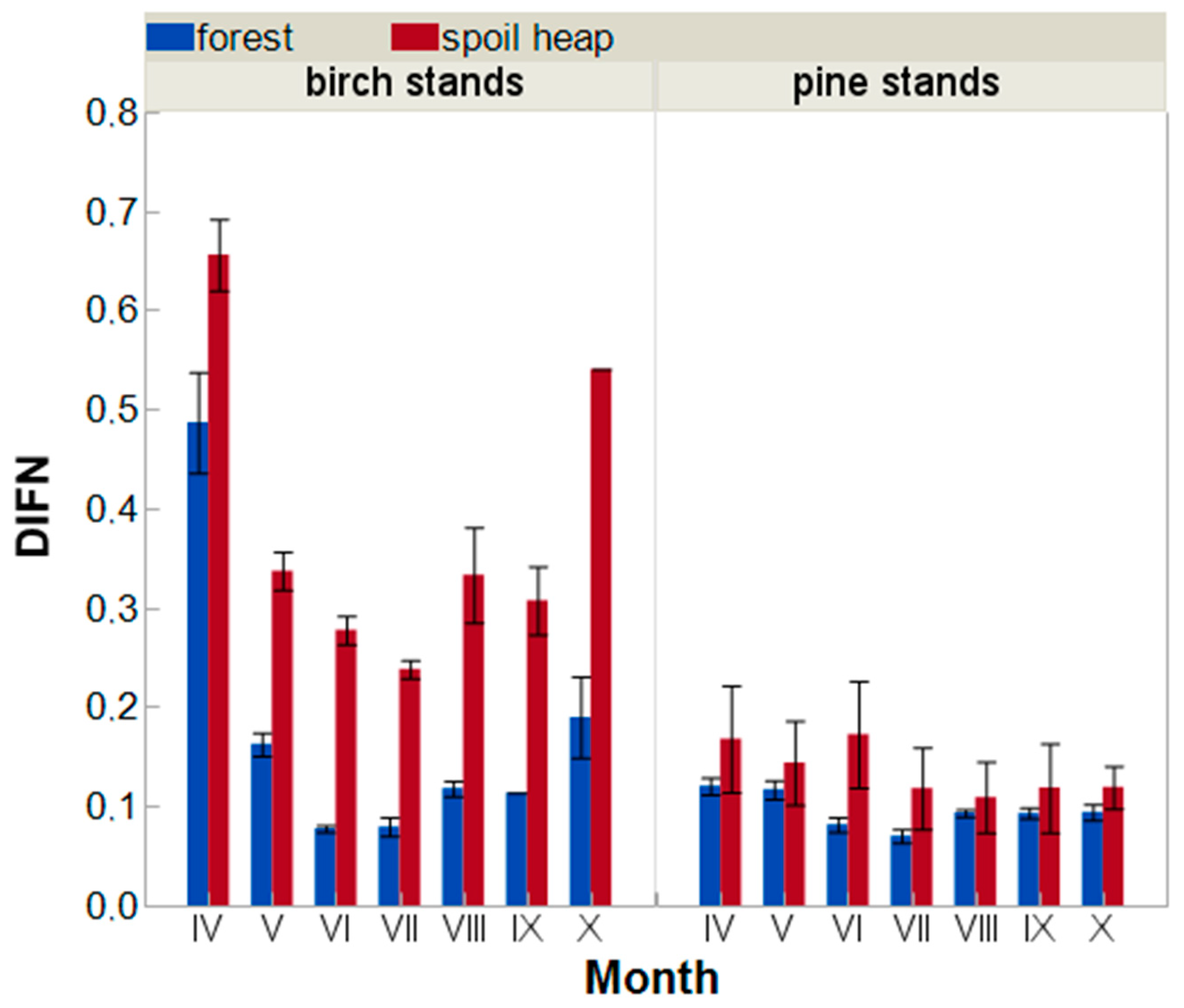
Microhabitat Conditions



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Number | Coordinates | Plot Area (m2) | Stand Type | Site Type | Year of Measurements | Soil Texture | Stand Age | Average DBH (cm) * ±SE | Average H (m) * ±SE | BA (m2 ha−1) *, ** | Stocking Density (individuals ha−1) *, ** |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 6 | 51.2207, 19.4339 | 604.5 | BePe | SH | 2011 | clay loam | 22 | 8.87 (0.30) | 9.21 (0.21) | 8.54 (98.31) | 1274 (74.76) |
| 2015 | 26 | 10.14 (0.36) | 10.61 (0.26) | 11.45 (98.22) | 1290 (62.40) | ||||||
| 7 | 51.2208, 19.4339 | 604.5 | 2011 | clay loam | 22 | 8.96 (0.34) | 9.09 (0.22) | 7.4 (95.55) | 1075 (69.15) | ||
| 2015 | 26 | 10.73 (0.39) | 11.26 (0.25) | 10.35 (95.24) | 1059 (57.66) | ||||||
| 15 | 51.2105, 19.4384 | 900 | PiSy | 2011 | sand | 18 | 5.49 (0.07) | 4.86 (0.04) | 12.76 (99.83) | 5022 (99.12) | |
| 2015 | 22 | 6.52 (0.09) | 6.58 (0.05) | 18.18 (99.80) | 5022 (98.69) | ||||||
| 16 | 51.2117, 19.4265 | 810 | 2011 | sand | 17 | 6.81 (0.11) | 6.05 (0.05) | 20.61 (99.96) | 5123 (99.28) | ||
| 2015 | 21 | 7.92 (0.13) | 8.05 (0.06) | 27.50 (99.94) | 5049 (99.27) | ||||||
| 105 | 51.2744, 19.4324 | 646.52 | BePe | F | 2012 | sandy loam | 20 | 10.40 (0.21) | 13.66 (0.11) | 15.79 (99.95) | 1779 (92.00) |
| 2015 | 23 | 10.76 (0.28) | 14.83 (0.23) | 17.57 (98.95) | 1794 (73.42) | ||||||
| 106 | 51.2747, 19.4326 | 450 | 2012 | clay loam | 20 | 10.10 (0.20) | 13.44 (0.12) | 18.32 (99.99) | 2200 (95.19) | ||
| 2015 | 23 | 10.45 (0.29) | 14.93 (0.25) | 20.05 (99.46) | 2178 (72.06) | ||||||
| 107 | 51.1738, 19.4323 | 750 | PiSy | 2012 | sand | 21 | 8.02 (0.15) | 8.90 (0.08) | 23.41 (100.00) | 4200 (99.68) | |
| 2015 | 24 | 8.99 (0.17) | 10.21 (0.08) | 28.76 (99.64) | 4093 (81.65) | ||||||
| 108 | 51.1746, 19.4321 | 750 | 2012 | sand | 21 | 8.46 (0.16) | 9.16 (0.08) | 21.22 (99.81) | 3467 (99.24) | ||
| 2015 | 24 | 9.30 (0.19) | 10.09 (0.11) | 26.12 (99.22) | 3467 (54.74) |
| Plot Number | Stand Type | Site Type | Humus Type | V (cm) | Corg. (%) | Ntot (%) | C/N | Hum (%) | pH H2O | Ca | Mg | K | Na |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 6 | BePe | SH | FPMD | 1 | 28.20 | 1.07 | 30.65 | 48.62 | 6.10 | 14.7300 | 1.5600 | 0.6600 | 0.0300 |
| 7 | FPMD | 1 | 32.83 | 1.36 | 28.09 | 56.59 | 6.12 | 14.0100 | 2.5100 | 0.9800 | 0.0200 | ||
| 15 | PiSy | FPM | 1 | 38.43 | 0.88 | 52.05 | 66.26 | 5.01 | 11.8420 | 1.0250 | 0.4390 | 0.0200 | |
| 16 | FPM | 2 | 46.20 | 0.96 | 56.21 | 79.65 | 4.93 | 9.2460 | 1.4910 | 1.1225 | 0.0300 | ||
| 105 | BePe | F | FMD | 3 | 34.17 | 1.33 | 36.20 | 58.91 | 5.55 | 13.6667 | 3.4367 | 2.5633 | 0.0300 |
| 106 | FMD | 2 | 37.23 | 1.29 | 37.98 | 64.19 | 5.57 | 13.8100 | 3.3850 | 2.2650 | 0.0300 | ||
| 107 | PiSy | FMM | 4 | 45.09 | 1.30 | 41.94 | 77.74 | 4.68 | 10.1025 | 1.2125 | 1.7075 | 0.0375 | |
| 108 | FM | 6 | 32.64 | 1.07 | 37.03 | 56.27 | 4.37 | 6.7410 | 0.8805 | 0.7256 | 0.0420 |
| Alnus glutinosa | Betula pendula | Pinus sylvestris | Quercus robur | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| birch stands | pine stands | birch stands | pine stands | birch stands | pine stands | birch stands | pine stands | |||
| Collection time (months) | 3 | F | 28.77 (1.18) | 12.97 (1.12) | 23.84 (1.61) | 14.33 (0.90) | 13.4 (2.17) | 6.23 (0.68) | 3.66 (0.59) | 1.55 (0.46) |
| SH | 11.89 (0.99) | 18.38 (0.93) | 14.07 (0.38) | 15.71 (0.82) | 4.98 (0.30) | 7.17 (0.77) | 0.44 (0.30) | 2.03 (0.63) | ||
| ANOVA | <0.0001 | 0.0023 | <0.0001 | 0.2790 | 0.0010 | 0.3591 | 0.0071 | 0.4176 | ||
| 6 | F | 47.84 (5.43) | 34.93 (1.79) | 34.16 (2.13) | 29.42 (1.19) | 25.33 (2.3) | 21.58 (2.21) | 7.63 (1.66) | 7.55 (0.67) | |
| SH | 43.82 (4.19) | 40.78 (2.73) | 25.77 (1.47) | 33.23 (2.19) | 8.29 (0.73) | 16.61 (1.40) | 3.36 (0.46) | 5.15 (0.49) | ||
| ANOVA | 0.5959 | 0.0943 | 0.0051 | 0.1441 | <0.0001 | 0.0874 | 0.0007 | 0.0118 | ||
| 9 | F | 68.18 (3.28) | 46.40 (2.70) | 57.96 (3.06) | 36.97 (1.12) | 40.73 (2.08) | 32.77 (1.15) | 27.3 (3.04) | 18.15 (1.27) | |
| SH | 57.51 (1.67) | 51.06 (2.49) | 37.72 (2.48) | 37.19 (1.26) | 18.38 (1.20) | 26.58 (2.07) | 9.24 (1.01) | 14.25 (2.34) | ||
| ANOVA | 0.0089 | 0.2246 | 0.0002 | 0.9004 | <0.0001 | 0.0206 | <0.0001 | 0.1238 | ||
| 12 | F | 84.98 (3.50) | 56.11 (3.96) | 70.88 (4.89) | 47.76 (3.29) | 50.50 (4.17) | 41.33 (1.63) | 51.74 (6.02) | 26.59 (2.98) | |
| SH | 68.54 (2.69) | 65.22 (3.85) | 52.40 (2.70) | 49.11 (1.90) | 24.67 (0.59) | 37.44 (2.05) | 13.61 (0.63) | 21.06 (3.45) | ||
| ANOVA | 0.0021 | 0.1293 | 0.0049 | 0.7447 | <0.0001 | 0.1627 | <0.0001 | 0.2096 | ||
| 15 | F | 83.71 (4.16) | 60.53 (2.95) | 77.11 (2.71) | 49.35 (1.05) | 58.53 (2.27) | 41.56 (0.77) | 53.81 (5.67) | 32.09 (4.82) | |
| SH | 78.14 (2.02) | 65.66 (3.77) | 59.65 (3.58) | 49.19 (1.57) | 26.88 (1.93) | 37.79 (1.65) | 19.01 (0.92) | 23.22 (4.05) | ||
| ANOVA | 0.1881 | 0.2991 | 0.0015 | 0.9320 | <0.0001 | 0.0558 | <0.0001 | 0.1631 | ||
| 18 | F | 87.66 (2.68) | 64.23 (1.84) | 82.38 (4.2) | 51.64 (2.25) | 66.46 (4.21) | 45.11 (2.29) | 58.95 (5.98) | 31.80 (6.19) | |
| SH | 78.63 (1.76) | 68.26 (2.43) | 68.49 (2.91) | 53.84 (3.38) | 27.69 (1.15) | 37.85 (1.27) | 29.28 (4.82) | 23.57 (4.94) | ||
| ANOVA | 0.0108 | 0.2014 | 0.0167 | 0.6046 | <0.0001 | 0.0145 | 0.0024 | 0.2807 | ||
| 21 | F | 93.08 (1.63) | 67.48 (2.02) | 86.25 (3.03) | 53.25 (2.65) | 65.76 (3.54) | 45.90 (1.71) | 64.40 (4.22) | 41.96 (3.42) | |
| SH | 81.24 (2.46) | 76.70 (4.14) | 77.60 (1.88) | 64.19 (5.76) | 39.29 (7.82) | 39.99 (2.41) | 26.60 (1.88) | 27.42 (3.82) | ||
| ANOVA | 0.001 | 0.0584 | 0.0278 | 0.1020 | 0.0179 | 0.0651 | <0.0001 | 0.0117 | ||
| 24 | F | 91.21 (1.63) | 71.22 (3.19) | 78.04 (2.63) | 55.60 (2.27) | 66.21 (3.78) | 53.71 (2.38) | 66.03 (3.39) | 43.18 (4.59) | |
| SH | 87.97 (1.91) | 68.46 (2.64) | 74.06 (2.56) | 56.06 (3.07) | 40.76 (6.03) | 45.82 (4.49) | 36.79 (5.55) | 31.66 (3.63) | ||
| ANOVA | 0.2167 | 0.4931 | 0.2826 | 0.8945 | 0.0040 | 0.1549 | 0.0005 | 0.0666 | ||
| 27 | F | 90.86 (2.49) | 73.61 (2.60) | 83.54 (4.99) | 53.38 (1.14) | 68.81 (2.98) | 50.79 (2.12) | 72.95 (3.60) | 49.89 (5.39) | |
| SH | 86.37 (1.74) | 73.96 (4.69) | 81.21 (3.18) | 65.56 (6.12) | 36.35 (1.15) | 49.88 (5.46) | 33.29 (3.58) | 46.27 (8.75) | ||
| ANOVA | 0.1158 | 0.7914 | 0.4891 | 0.0828 | <0.0001 | 0.9572 | <0.0001 | 0.7405 | ||
| 30 | F | 90.10 (2.85) | 80.43 (2.88) | 87.54 (2.02) | 66.05 (3.08) | 78.67 (3.65) | 56.19 (2.98) | 72.44 (3.95) | 53.88 (4.65) | |
| SH | 85.59 (1.58) | 78.70 (3.88) | 77.18 (2.95) | 65.11 (4.48) | 42.42 (3.19) | 53.98 (3.94) | 35.94 (4.68) | 43.49 (5.77) | ||
| ANOVA | 0.1296 | 0.7918 | 0.0120 | 0.9147 | <0.0001 | 0.6730 | <0.0001 | 0.1784 | ||
| 33 | F | 92.71 (1.54) | 70.89 (3.05) | 88.66 (1.95) | 60.57 (2.73) | 77.87 (3.61) | 57.03 (3.65) | 74.92 (2.16) | 58.84 (3.71) | |
| SH | 84.09 (1.28) | 76.57 (3.16) | 81.22 (2.00) | 69.66 (5.27) | 38.36 (1.13) | 53.19 (4.36) | 27.48 (1.67) | 34.25 (4.85) | ||
| ANOVA | 0.0013 | 0.2216 | 0.0180 | 0.1363 | <0.0001 | 0.5305 | <0.0001 | 0.0013 | ||
| 36 | F | 91.86 (2.46) | 69.37 (2.36) | 83.64 (2.20) | 64.03 (2.39) | 78.47 (3.42) | 66.29 (3.00) | 76.43 (3.90) | 53.70 (3.44) | |
| SH | 85.64 (1.75) | 75.09 (3.47) | 77.74 (1.34) | 63.70 (3.31) | 44.26 (2.84) | 55.23 (2.66) | 40.61 (6.56) | 39.58 (4.72) | ||
| ANOVA | 0.4901 | 0.1763 | 0.7832 | 0.9527 | <0.0001 | 0.0162 | 0.0004 | 0.0300 | ||
References
- Jagodziński, A.M.; Kałucka, I.; Horodecki, P.; Oleksyn, J. Aboveground biomass allocation and accumulation in a chronosequence of young Pinus sylvestris stands growing on a lignite mine spoil heap. Dendrobiology 2014, 72, 139–150. [Google Scholar] [CrossRef]
- Jimenez, M.D.; Ruiz-Capillas, P.; Mola, I.; Pérez-Corona, E.; Casado, M.A.; Balaguer, L. Soil Development at the Roadside: A Case Study of a Novel Ecosystem. Land Degrad. Dev. 2013, 24, 564–574. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Higgs, E.; Harris, J.A. Novel ecosystems: Implications for conservation and restoration. Trends Ecol. Evol. 2009, 24, 599–605. [Google Scholar] [CrossRef]
- Maiti, S.K. Bioreclamation of coalmine overburden dumps—With special empasis on micronutrients and heavy metals accumulation in tree species. Environ. Monit. Assess. 2007, 125, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Jagodziński, A.M.; Wierzcholska, S.; Dyderski, M.K.; Horodecki, P.; Rusińska, A.; Gdula, A.K.; Kasprowicz, M. Tree species effects on bryophyte guilds on a reclaimed post-mining site. Ecol. Eng. 2018, 110, 117–127. [Google Scholar] [CrossRef]
- Prach, K.; Hobbs, R.J. Spontaneous Succession versus Technical Reclamation in the Restoration of Disturbed Sites. Restor. Ecol. 2008, 16, 363–366. [Google Scholar] [CrossRef]
- Dutta, R.K.; Agrawal, M. Litterfall, litter decomposition and nutrient release in five exotic plant species planted on coal mine spoils. Pedobiologia 2001, 45, 298–312. [Google Scholar] [CrossRef]
- Esperschütz, J.; Zimmermann, C.; Dümig, A.; Welzl, G.; Buegger, F.; Elmer, M.; Munch, J.C.; Schloter, M. Dynamics of microbial communities during decomposition of litter from pioneering plants in initial soil ecosystems. Biogeosciences 2013, 10, 5115–5124. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Kałucka, I. Fine roots biomass and morphology in a chronosequence of young Pinus sylvestris stands growing on a reclaimed lignite mine spoil heap. Dendrobiology 2010, 64, 19–30. [Google Scholar]
- Kałucka, I.L.; Jagodziński, A.M. Successional traits of ectomycorrhizal fungi in forest reclamation after surface mining and agricultural disturbances: A review. Dendrobiology 2016, 76, 91–104. [Google Scholar] [CrossRef]
- Pietrzykowski, M. Reclamation and Reconstruction of Terrestrial Ecosystems on Mine Sites-Ecological Effectiveness Assessment. In Energy Science and Technology; Sivakumar, S., Sharma, U.C., Prasad, R., Eds.; Coal Energy; Studium Press LLC: New Delhi, India, 2015; pp. 121–151. ISBN 1-62699-063-8. [Google Scholar]
- Frouz, J.; Livečková, M.; Albrechtová, J.; Chroňáková, A.; Cajthaml, T.; Pižl, V.; Háněl, L.; Starý, J.; Baldrian, P.; Lhotáková, Z.; et al. Is the effect of trees on soil properties mediated by soil fauna? A case study from post-mining sites. For. Ecol. Manag. 2013, 309, 87–95. [Google Scholar] [CrossRef]
- Helingerová, M.; Frouz, J.; Šantrůčková, H. Microbial activity in reclaimed and unreclaimed post-mining sites near Sokolov (Czech Republic). Ecol. Eng. 2010, 36, 768–776. [Google Scholar] [CrossRef]
- Laganière, J.; Paré, D.; Bradley, R.L. How does a tree species influence litter decomposition? Separating the relative contribution of litter quality, litter mixing, and forest floor conditions. Can. J. For. Res. 2010, 40, 465–475. [Google Scholar] [CrossRef]
- Pietrzykowski, M. Soil quality index as a tool for Scots pine (Pinus sylvestris) monoculture conversion planning on afforested, reclaimed mine land. J. For. Res. 2014, 25, 63–74. [Google Scholar] [CrossRef]
- Józefowska, A.; Pietrzykowski, M.; Woś, B.; Cajthaml, T.; Frouz, J. The effects of tree species and substrate on carbon sequestration and chemical and biological properties in reforested post-mining soils. Geoderma 2017, 292, 9–16. [Google Scholar] [CrossRef]
- Horodecki, P.; Jagodziński, A.M. Tree species effects on litter decomposition in pure stands on afforested post-mining sites. For. Ecol. Manag. 2017, 406, 1–11. [Google Scholar] [CrossRef]
- Sroka, K.; Chodak, M.; Klimek, B.; Pietrzykowski, M. Effect of black alder (Alnus glutinosa) admixture to Scots pine (Pinus sylvestris) plantations on chemical and microbial properties of sandy mine soils. Appl. Soil Ecol. 2018, 124, 62–68. [Google Scholar] [CrossRef]
- Doley, D.; Audet, P. Adopting novel ecosystems as suitable rehabilitation alternatives for former mine sites. Ecol. Process. 2013, 2, 1–11. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Maiti, S.K.; Masto, R.E. Development of mine soil quality index (MSQI) for evaluation of reclamation success: A chronosequence study. Ecol. Eng. 2014, 71, 10–20. [Google Scholar] [CrossRef]
- Macdonald, S.E.; Landhäusser, S.M.; Skousen, J.; Franklin, J.; Frouz, J.; Hall, S.; Jacobs, D.F.; Quideau, S. Forest restoration following surface mining disturbance: Challenges and solutions. New For. 2015, 46, 703–732. [Google Scholar] [CrossRef]
- Pietrzykowski, M.; Woś, B.; Haus, N. Scots pine needles macronutrient (N, P, K, Ca, Mg, and S) supply at different reclaimed mine soil substrates—As an indicator of the stability of developed forest ecosystems. Environ. Monit. Assess. 2013, 185, 7445–7457. [Google Scholar] [CrossRef]
- Ludwig, J.A.; Hindley, N.; Barnett, G. Indicators for monitoring minesite rehabilitation: Trends on waste-rock dumps, northern Australia. Ecol. Indic. 2003, 3, 143–153. [Google Scholar] [CrossRef]
- Pietrzykowski, M.; Pająk, M.; Krzaklewski, W. Assessment of soil-site conditions on the spoil heaps of the lignite mining plant (KWB) “Bełchatów” reclaimed to forest with the use of the site soil index (SIG). Pol. J. Soil Sci. 2011, 44, 81–88. [Google Scholar]
- Pietrzykowski, M.; Pająk, M.; Krzaklewski, W. Próba zastosowania metod liczbowej wyceny gleb na podstawie Indeksu Trofizmu Gleb Leśnych (ITGL) oraz Siedliskowego Indeksu Glebowego (SIG) do opisu zmienności warunków siedliskowych na zrekultywowanych dla leśnictwa zwałowiskach KWB “Bełchatów”. Gospod. Surowcami Miner. 2010, 26, 155–165. [Google Scholar]
- Asensio, V.; Guala, S.D.; Vega, F.A.; Covelo, E.F. A soil quality index for reclaimed mine soils. Environ. Toxicol. Chem. 2013, 32, 2240–2248. [Google Scholar] [CrossRef]
- Monokrousos, N.; Boutsis, G.; Diamantopoulos, J.D. Development of soil chemical and biological properties in the initial stages of post-mining deposition sites. Environ. Monit. Assess. 2014, 186, 9065–9074. [Google Scholar] [CrossRef]
- Berg, B. Decomposition patterns for foliar litter—A theory for influencing factors. Soil Biol. Biochem. 2014, 78, 222–232. [Google Scholar] [CrossRef]
- Frouz, J.; Roubíčková, A.; Heděnec, P.; Tajovský, K. Do soil fauna really hasten litter decomposition? A meta-analysis of enclosure studies. Eur. J. Soil Biol. 2015, 68, 18–24. [Google Scholar] [CrossRef]
- Lawrey, J.D. The relative decomposition potential of habitats variously affected by surface coal mining. Can. J. Bot. 1977, 55, 1544–1552. [Google Scholar] [CrossRef]
- Horodecki, P.; Nowiński, M.; Jagodziński, A.M. Advantages of mixed tree stands in restoration of upper soil layers on postmining sites: A five-year leaf litter decomposition experiment. Land Degrad. Dev. 2019, 30, 3–13. [Google Scholar] [CrossRef]
- Frouz, J. The effect of litter type and macrofauna community on litter decomposition and organic matter accumulation in post-mining sites. Biologia 2008, 63, 249–253. [Google Scholar] [CrossRef]
- Urbanová, M.; Šnajdr, J.; Brabcová, V.; Merhautová, V.; Dobiášová, P.; Cajthaml, T.; Vaněk, D.; Frouz, J.; Šantrůčková, H.; Baldrian, P. Litter decomposition along a primary post-mining chronosequence. Biol. Fertil. Soils 2014, 50, 827–837. [Google Scholar] [CrossRef]
- Zielony, R.; Kliczkowska, A. Regionalizacja Przyrodniczo-Leśna Polski 2010; Centrum Informacyjne Lasów Państwowych: Na zlec; Dyrekcji Generalnej Lasów Państwowych: Warszawa, Poland, 2012; ISBN 978-83-61633-62-4.
- Goździk, J.; Jończyk, W.; Niżnik, A.M. Kopalnia Węgla Brunatnego Bełchatów i Elektrownia Bełchatów—Zmiany środowiska geograficznego spowodowane ich działalnością. In Proceedings of the Obszary Metropolitalne We Współczesnym Środowisku Geograficznym, Łódź, Poland, 8–12 September 2010; Barwiński, M., Ed.; Wydawnictwo Triada: Łódź, Poland, 2010; Volume 2, pp. 397–422. [Google Scholar]
- Pietrzykowski, M. Scots pine (Pinus sylvestris L.) ecosystem macronutrients budget on reclaimed mine sites—Stand trees supply and stability. Nat. Sci. 2010, 2, 590–599. [Google Scholar] [CrossRef]
- Rawlik, M.; Kasprowicz, M.; Jagodziński, A.M. Differentiation of herb layer vascular flora in reclaimed areas depends on the species composition of forest stands. For. Ecol. Manag. 2018, 409, 541–551. [Google Scholar] [CrossRef]
- Wójcik, J.; Krzaklewski, W. Metody rekultywacji leśnej terenów bezglebowych w górnictwie odkrywkowym. Prz. Górniczy 2010, 10, 115–119. [Google Scholar]
- Olson, J.S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Berg, B.; Ekbohm, G. Litter mass-loss rates and decomposition patterns in some needle and leaf litter types. Long-term decomposition in a Scots pine forest. VII. Can. J. Bot. 1991, 69, 1449–1456. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C.; Santo, A.V.D.; Johnson, D. Humus buildup in boreal forests: Effects of litter fall and its N concentration. Can. J. For. Res. 2001, 31, 988–998. [Google Scholar] [CrossRef]
- Benayas, J.M.R.; Newton, A.C.; Diaz, A.; Bullock, J.M. Enhancement of Biodiversity and Ecosystem Services by Ecological Restoration: A Meta-Analysis. Science 2009, 325, 1121–1124. [Google Scholar] [CrossRef]
- Audet, P.; Pinno, B.D.; Thiffault, E. Reclamation of boreal forest after oil sands mining: Anticipating novel challenges in novel environments. Can. J. For. Res. 2015, 45, 364–371. [Google Scholar] [CrossRef]
- Frouz, J. Effects of Soil Development Time and Litter Quality on Soil Carbon Sequestration: Assessing Soil Carbon Saturation with a Field Transplant Experiment along a Post-mining Chronosequence: Carbon Sequestration and Saturation in Post-mining Sites. Land Degrad. Dev. 2017, 28, 664–672. [Google Scholar] [CrossRef]
- Cornelissen, J.H.; Pérez-Harguindeguy, N.; Díaz, S.; Grime, J.P.; Marzano, B.; Cabido, M.; Vendramini, F.; Cerabolini, B. Leaf structure and defence control litter decomposition rate across species and life forms in regional floras on two continents. New Phytol. 1999, 143, 191–200. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Thompson, K. Functional leaf attributes predict litter decomposition rate in herbaceous plants. New Phytol. 1997, 135, 109–114. [Google Scholar] [CrossRef]
- Austin, A.T.; Vivanco, L.; González-Arzac, A.; Pérez, L.I. There’s no place like home? An exploration of the mechanisms behind plant litter-decomposer affinity in terrestrial ecosystems. New Phytol. 2014, 204, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Ayres, E.; Steltzer, H.; Simmons, B.L.; Simpson, R.T.; Steinweg, J.M.; Wallenstein, M.D.; Mellor, N.; Parton, W.J.; Moore, J.C.; Wall, D.H. Home-field advantage accelerates leaf litter decomposition in forests. Soil Biol. Biochem. 2009, 41, 606–610. [Google Scholar] [CrossRef]
- Aerts, R. Climate, Leaf Litter Chemistry and Leaf Litter Decomposition in Terrestrial Ecosystems: A Triangular Relationship. Oikos 1997, 79, 439–449. [Google Scholar] [CrossRef]
- Berg, B.; Staaf, H. Decomposition rate and chemical changes of Scots pine needle litter. II. Influence of chemical composition. Ecol. Bull. 1980, 32, 373–390. [Google Scholar]
- Frouz, J.; Thébault, E.; Pižl, V.; Adl, S.; Cajthaml, T.; Baldrián, P.; Háněl, L.; Starý, J.; Tajovský, K.; Materna, J.; et al. Soil Food Web Changes during Spontaneous Succession at Post Mining Sites: A Possible Ecosystem Engineering Effect on Food Web Organization? PLoS ONE 2013, 8, e79694. [Google Scholar] [CrossRef]
- Frouz, J.; Prach, K.; Pižl, V.; Háněl, L.; Starý, J.; Tajovský, K.; Materna, J.; Balík, V.; Kalčík, J.; Řehounková, K. Interactions between soil development, vegetation and soil fauna during spontaneous succession in post mining sites. Eur. J. Soil Biol. 2008, 44, 109–121. [Google Scholar] [CrossRef]
- Hopkins, D.W.; Badalucco, L.; English, L.C.; Meli, S.M.; Chudek, J.A.; Ioppolo, A. Plant litter decomposition and microbial characteristics in volcanic soils (Mt Etna, Sicily) at different stages of development. Biol. Fertil. Soils 2007, 43, 461–469. [Google Scholar] [CrossRef]
- Jangid, K.; Williams, M.A.; Franzluebbers, A.J.; Schmidt, T.M.; Coleman, D.C.; Whitman, W.B. Land-use history has a stronger impact on soil microbial community composition than aboveground vegetation and soil properties. Soil Biol. Biochem. 2011, 43, 2184–2193. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Dyderski, M.K.; Horodecki, P.; Rawlik, K. Limited dispersal prevents Quercus rubra invasion in a 14-species common garden experiment. Divers. Distrib. 2018, 24, 403–414. [Google Scholar] [CrossRef]
- Mueller, K.E.; Eisenhauer, N.; Reich, P.B.; Hobbie, S.E.; Chadwick, O.A.; Chorover, J.; Dobies, T.; Hale, C.M.; Jagodziński, A.M.; Kałucka, I.; et al. Light, earthworms, and soil resources as predictors of diversity of 10 soil invertebrate groups across monocultures of 14 tree species. Soil Biol. Biochem. 2016, 92, 184–198. [Google Scholar] [CrossRef]
- Frouz, J.; Keplin, B.; Pižl, V.; Tajovský, K.; Starý, J.; Lukešová, A.; Nováková, A.; Balík, V.; Háněl, L.; Materna, J.; et al. Soil biota and upper soil layer development in two contrasting post-mining chronosequences. Ecol. Eng. 2001, 17, 275–284. [Google Scholar] [CrossRef]
- Kraus, T.E.C.; Dahlgren, R.A.; Zasoski, R.J. Tannins in nutrient dynamics of forest ecosystems—A review. Plant Soil 2003, 256, 41–66. [Google Scholar] [CrossRef]
- Gao, J.; Kang, F.; Han, H. Effect of Litter Quality on Leaf-Litter Decomposition in the Context of Home-Field Advantage and Non-Additive Effects in Temperate Forests in China. Pol. J. Environ. Stud. 2016, 25, 1911–1920. [Google Scholar] [CrossRef]
- Berg, B. Decomposing litter; limit values; humus accumulation, locally and regionally. Appl. Soil Ecol. 2018, 123, 494–508. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Plant Litter. Decomposition, Humus Formation, Carbon Sequestration, 2nd ed.; Springer: Berlin, Germany, 2008; ISBN 978-3-540-74923-3. [Google Scholar]
- Bocock, K.L.; Gilbert, O.; Capstick, C.K.; Twinn, D.C.; Waid, J.S.; Woodman, M.J. Changes in Leaf Litter When Placed on the Surface of Soils with Contrasting Humus Types. J. Soil Sci. 1960, 11, 1–9. [Google Scholar] [CrossRef]
- Howard, D.M.; Howard, P.J.A. Effect of Species, Source of Litter, Type of Soil, and Climate on Litter Decomposition: Microbial Decomposition of Tree and Shrub Leaf Litter 3. Oikos 1980, 34, 115–124. [Google Scholar] [CrossRef]
- Smolander, A.; Kitunen, V. Soil microbial activities and characteristics of dissolved organic C and N in relation to tree species. Soil Biol. Biochem. 2002, 34, 651–660. [Google Scholar] [CrossRef]
- Meentemeyer, V.; Berg, B. Regional variation in rate of mass loss of Pinus sylvestris needle litter in Swedish pine forests as influenced by climate and litter quality. Scand. J. For. Res. 1986, 1, 167–180. [Google Scholar] [CrossRef]
- Berg, B.; Matzner, E. Effect of N deposition on decomposition of plant litter and soil organic matter in forest systems. Environ. Rev. 1997, 5, 1–25. [Google Scholar] [CrossRef]
- Zhang, W.; Gao, D.; Chen, Z.; Li, H.; Deng, J.; Qiao, W.; Han, X.; Yang, G.; Feng, Y.; Huang, J. Substrate quality and soil environmental conditions predict litter decomposition and drive soil nutrient dynamics following afforestation on the Loess Plateau of China. Geoderma 2018, 325, 152–161. [Google Scholar] [CrossRef]
- Mo, J.; Brown, S.; Xue, J.; Fang, Y.; Li, Z. Response of litter decomposition to simulated N deposition in disturbed, rehabilitated and mature forests in subtropical China. Plant Soil 2006, 282, 135–151. [Google Scholar] [CrossRef]






| Source | DF | Sum of Squares | F Ratio | Prob > F |
|---|---|---|---|---|
| TIME | 11 | 197,834.63 | 501.8998 | <0.0001 |
| LITTER | 3 | 115,072.35 | 1070.4271 | <0.0001 |
| TIME × LITTER | 33 | 3685.83 | 3.1169 | <0.0001 |
| SITE | 1 | 17,644.88 | 492.4091 | <0.0001 |
| TIME × SITE | 11 | 1276.65 | 3.2388 | 0.0002 |
| LITTER × SITE | 3 | 7632.87 | 71.0026 | <0.0001 |
| TIME × LITTER × SITE | 33 | 2378.31 | 2.0112 | 0.0006 |
| STAND | 1 | 16,147.70 | 450.6278 | <0.0001 |
| TIME × STAND | 11 | 3450.82 | 8.7546 | <0.0001 |
| LITTER × STAND | 3 | 5066.24 | 47.1272 | <0.0001 |
| TIME × LITTER × STAND | 33 | 1446.44 | 1.2232 | 0.1808 |
| SITE × STAND | 1 | 13,541.15 | 377.8877 | <0.0001 |
| TIME × SITE × STAND | 11 | 833.87 | 2.1155 | 0.0168 |
| LITTER × SITE × STAND | 3 | 454.71 | 4.2298 | 0.0055 |
| TIME × LITTER × SITE × STAND | 33 | 1082.21 | 0.9152 | 0.6068 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horodecki, P.; Jagodziński, A.M. Site Type Effect on Litter Decomposition Rates: A Three-Year Comparison of Decomposition Process between Spoil Heap and Forest Sites. Forests 2019, 10, 353. https://doi.org/10.3390/f10040353
Horodecki P, Jagodziński AM. Site Type Effect on Litter Decomposition Rates: A Three-Year Comparison of Decomposition Process between Spoil Heap and Forest Sites. Forests. 2019; 10(4):353. https://doi.org/10.3390/f10040353
Chicago/Turabian StyleHorodecki, Paweł, and Andrzej M. Jagodziński. 2019. "Site Type Effect on Litter Decomposition Rates: A Three-Year Comparison of Decomposition Process between Spoil Heap and Forest Sites" Forests 10, no. 4: 353. https://doi.org/10.3390/f10040353
APA StyleHorodecki, P., & Jagodziński, A. M. (2019). Site Type Effect on Litter Decomposition Rates: A Three-Year Comparison of Decomposition Process between Spoil Heap and Forest Sites. Forests, 10(4), 353. https://doi.org/10.3390/f10040353