Three Main Genes in the MAPK Cascade Involved in the Chinese Jujube-Phytoplasma Interaction

Abstract
:1. Introduction
2. Results
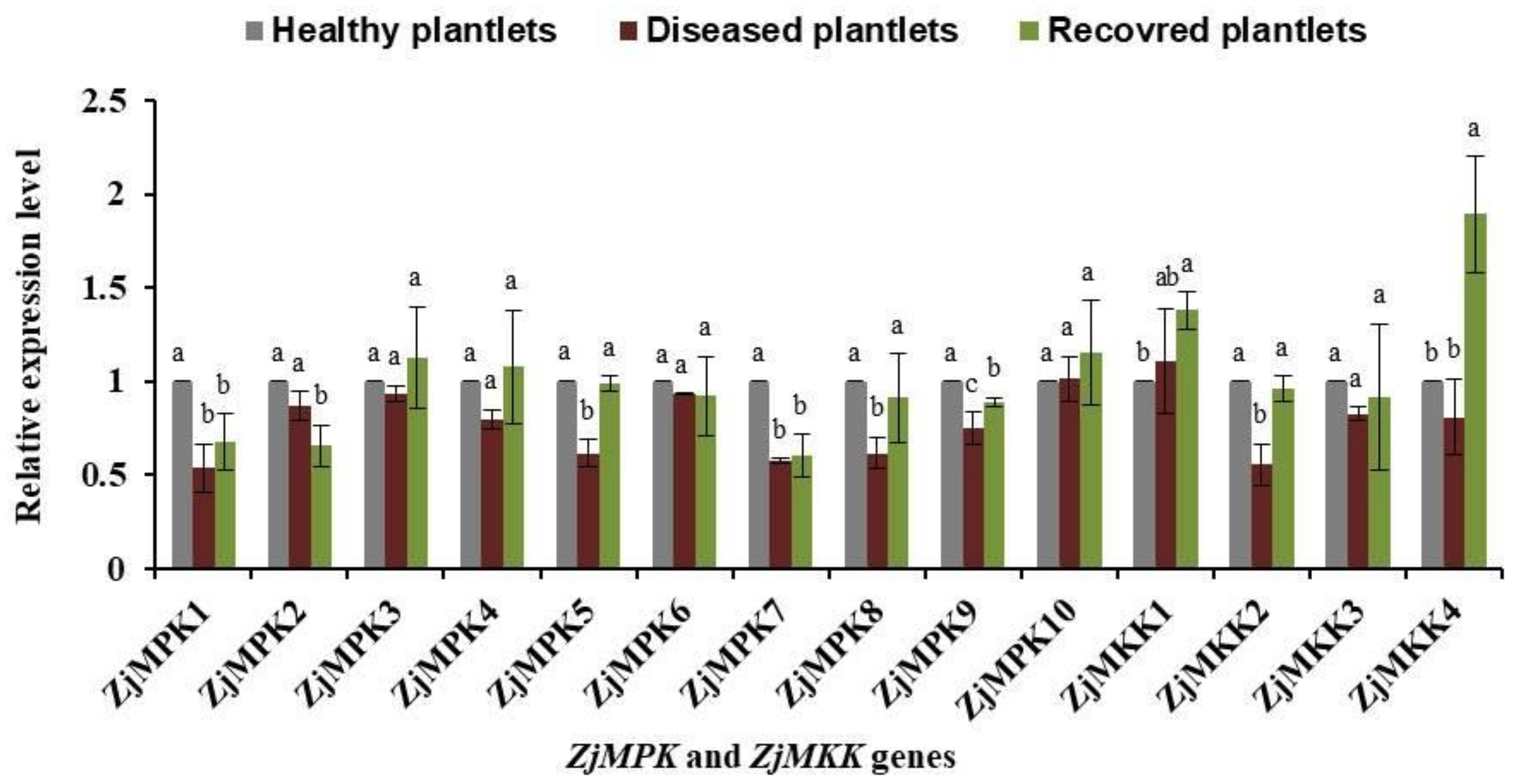
2.1. Expression Patterns of ZjMPK and ZjMKK Genes in Healthy, JWB-diseased and Recovered Plantlets
2.2. Expression Modules of ZjMPK and ZjMKK Genes in Tissues Showing Different JWB Disease Symptoms
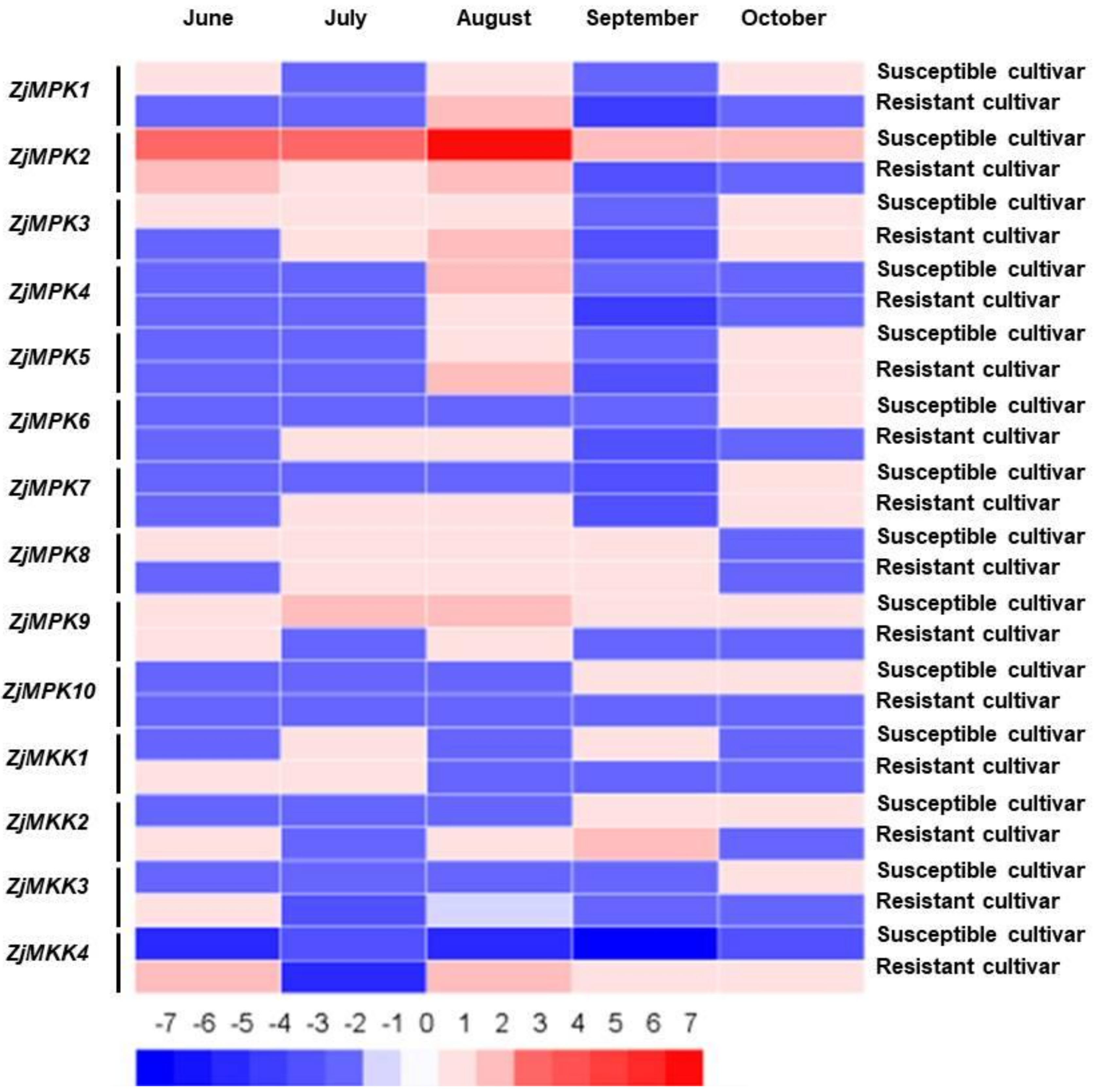
2.3. Expression Profiles of ZjMPK and ZjMKK Genes in Resistant and Susceptible Cultivars
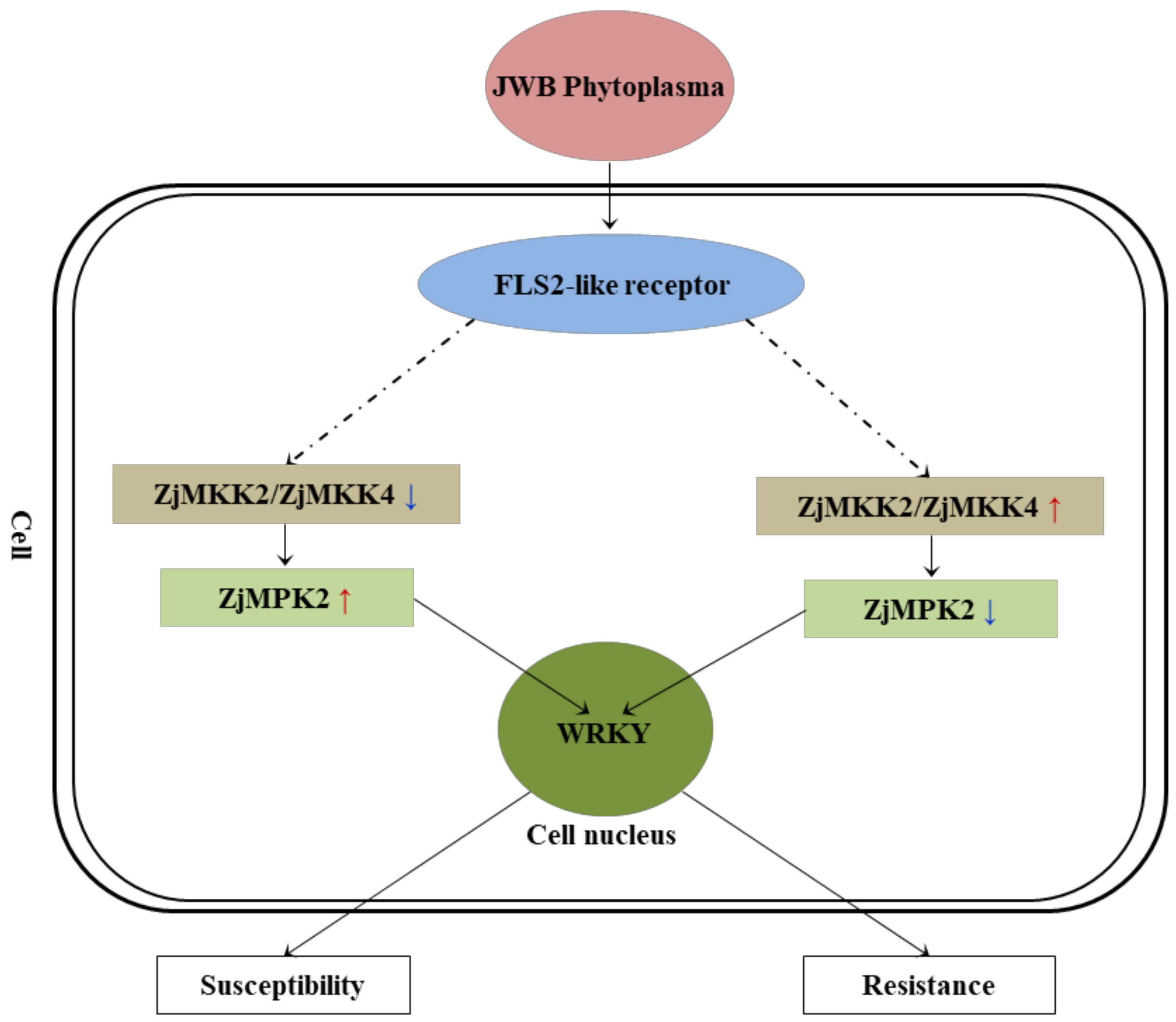
2.4. Candidate Genes Identification and Protein-protein Interaction Analysis
2.5. The Transcriptional Difference of Candidate Genes between Susceptible and Resistant Cultivars
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Diseased, Recovered and Healthy Plantlets
5.2. The Tissues Showing Different JWB Disease Symptoms
5.3. Resistant and Susceptible Cultivars
5.4. Total RNA Extraction
5.5. Quantitative Real-Time PCR System
5.6. Expression Analysis
5.7. Heatmap Construction
5.8. Protein-Protein Interaction Analysis
5.9. Yeast Two-Hybrid Screening (Y2H)
5.10. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- DoI, Y.; Teranaka, M.; Yora, K.; Yora, K.; Asuyama, H. Mycoplasma- or PLT group-like microorganisms found in the phloem elements of plants infected with Mulberry dwarf, Potato witches’ broom, Aster yellows, or Paulownia witches’ broom. Jpn. J. Phytopathol. 1967, 33, 259–266. [Google Scholar] [CrossRef]
- Strauss, E. Microbiology. Phytoplasma research begins to bloom. Science 2009, 325, 388–390. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.M.; Davis, R.E.; Gundersen-Rindal, D.E. Phytoplasma: Phytopathogenic mollicutes. Annu. Rev. Microbiol. 2000, 54, 221–255. [Google Scholar] [CrossRef] [PubMed]
- Seemüller, E.; Marcone, C.; Lauer, U.; Ragozzino, A.; Göschl, M. Current status of molecular classification of the phytoplasmas. J. Plant Pathol. 1998, 80, 3–26. [Google Scholar]
- Maclean, A.M.; Sugio, A.; Makarova, O.V.; Findlay, K.C.; Grieve, V.M.; Tóth, R.; Nicolaisen, M.; Hogenhout, S.A. Phytoplasma effector sap54 induces indeterminate leaf-like flower development in arabidopsis plants. Plant Physiol. 2011, 157, 831–841. [Google Scholar] [CrossRef]
- Sugio, A.; Kingdom, H.N.; Maclean, A.M.; Grieve, V.M.; Hogenhout, S.A. Phytoplasma protein effector sap11 enhances insect vector reproduction by manipulating plant development and defense hormone biosynthesis. Proc. Natl. Acad. Sci. USA 2011, 108, E1254–E1263. [Google Scholar] [CrossRef]
- Minato, N.; Himeno, M.; Hoshi, A.; Maejima, K.; Komatsu, K.; Takebayashi, Y.; Kasahara, H.; Yusa, A.; Yamaji, Y.; Oshima, K.; et al. The phytoplasmal virulence factor tengu causes plant sterility by downregulating of the jasmonic acid and auxin pathways. Sci. Rep. 2014, 4, 7399. [Google Scholar] [CrossRef]
- Shi, Q.H.; Liu, P.; Liu, M.J.; Wang, J.R.; Xu, J. A novel method for rapid in vivo induction of homogeneous polyploids via calluses in a woody fruit tree (Ziziphus jujuba Mill.). Plant Cell Tiss. Organ Cult. 2015, 121, 423–433. [Google Scholar] [CrossRef]
- Jung, H.Y.; Sawayanagi, T.; Kakizawa, S.; Nishigawa, H.; Wei, W.; Oshima, K.; Miyata, S.; Ugaki, M.; Hibi, T.; Namba, S. ’Candidatus Phytoplasma ziziphi’, a novel phytoplasma taxon associated with jujube witches’-broom disease. Int. J. Syst. Evol. Microbiol. 2003, 53 (Pt 4), 1037–1041. [Google Scholar] [CrossRef]
- Liu, M.J.; Zhao, J.; Zhou, J.Y. Jujube Witches’ Broom Disease; China Agriculture Press: Beijing, China, 2010; p. 2. [Google Scholar]
- Seemüller, E.; Garnier, M.; Schneider, B. Mycoplasmas of Plants and Insects. In Molecular Biology and Pathogenicity of Mycoplasmas; Razin, S., Herrmann, R., Eds.; Springer: Boston, MA, USA, 2002; pp. 91–115. [Google Scholar]
- Osler, R.; Borselli, S.; Ermacora, P.; Loschi, A.; Martini, M.; Musetti, R.; Loi, N. Acquired tolerance in Apricot plants that stably recovered from European stone fruit yellows. Plant Dis. 2014, 98, 492–496. [Google Scholar] [CrossRef]
- Contaldo, N.; Satta, E.; Zambon, Y.; Paltrinieri, S.; Bertaccini, A. Development and evaluation of different complex media for phytoplasma isolation and growth. J. Microbiol. Methods 2016, 127, 105–110. [Google Scholar] [CrossRef]
- Wang, J.; Song, L.; Jiao, Q.; Yang, S.; Gao, R.; Lu, X.; Zhou, G. Comparative genome analysis of jujube witches’-broom phytoplasma, an obligate pathogen that causes jujube witches’-broom disease. BMC Genomics 2018, 19, 689. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, B.C. Mycoplasma-Like Organisms-Plant and Invertebrate Pathogens; Springer: New York, NY, USA, 1992; pp. 4050–4067. [Google Scholar]
- Mccoy, R.E.; Caudwell, A.; Chang, C.J.; Chen, T.A.; Chiykowski, L.N.; Cousin, M.T.; Dale, J.L.; de Leeuw, G.T.N.; Golino, D.A.; Hackett, K.J.; et al. Plant diseases associated with mycoplasma-like organisms. Mycoplasmas 1989, 545–640. [Google Scholar] [CrossRef]
- Tena, G.; Asai, T.; Chiu, W.L.; Sheen, J. Plant mitogen-activated protein kinase signaling cascades. Curr. Opin. Plant Biol. 2001, 4, 392–400. [Google Scholar] [CrossRef]
- Chen, Z.; Gibson, T.B.; Robinson, F.; Silvestro, L.; Pearson, G.; Xu, B.; Wright, A.; Vanderbilt, C.; Cobb, M.H. MAP kinases. Chem. Rev. 2001, 101, 2449–2476. [Google Scholar] [CrossRef]
- MAPK Group. Mitogen-activated protein kinase cascades in plants: A new nomenclature. Trends Plant Sci. 2002, 7, 301–308. [Google Scholar] [CrossRef]
- Rodriguez, M.C.; Petersen, M.; Mundy, J. Mitogen-activated protein kinase signaling in plants. Annu. Rev. Plant Biol. 2010, 61, 621–649. [Google Scholar] [CrossRef] [PubMed]
- Jonak, C.; Okrész, L.; Bögre, L.; Hirt, H. Complexity, cross talk and integration of plant MAP kinase signaling. Curr. Opin. Plant Biol. 2002, 5, 415–424. [Google Scholar] [CrossRef]
- Asai, T.; Tena, G.; Plotnikova, J.; Willmann, M.R.; Chiu, W.L.; Gomez-Gomez, L.; Boller, T.; Ausubel, F.M.; Sheen, J. MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 2002, 415, 977–983. [Google Scholar] [CrossRef]
- Berriri, S.; Garcia, A.V.; dit Frey, N.F.; Rozhon, W.; Pateyron, S.; Leonhardt, N.; Montillet, J.L.; Leung, J.; Hirt, H.; Colcombet, J. Constitutively active mitogen-activated protein kinase versions reveal functions of Arabidopsis MPK4 in pathogen defense signaling. The Plant Cell. 2012, 24, 4281–4293. [Google Scholar] [CrossRef]
- Gao, M.; Liu, J.; Bi, D.; Zhang, Z.; Cheng, F.; Chen, S.; Zhang, Y. MEKK1, MKK1/MKK2 and MPK4 function together in a mitogen-activated protein kinase cascade to regulate innate immunity in plants. Cell Res. 2008, 18, 1190–1198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, Y.; Klessig, D.F. Multiple levels of tobacco WIPK activation during the induction of cell death by fungal elicitins. Plant J. 2000, 23, 339–347. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Du, H.; Klessig, D.F. Activation of the tobacco SIP kinase by both a cell wall-derived carbohydrate elicitor and purified proteinaceous elicitins from Phytophthora spp. Plant Cell 1998, 10, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Kishi-Kaboshi, M.; Okada, K.; Kurimoto, L.; Murakami, S.; Umezawa, T.; Shibuya, N.; Yamane, H.; Miyao, A.; Takatsuji, H.; Takahashi, A.; et al. A rice fungal MAMP-responsive MAPK cascade regulates metabolic flow to antimicrobial metabolite synthesis. Plant J. 2010, 63, 599–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Zhang, L.; An, H.; Wu, C.; Guo, X. GhMPK16, a novel stress-responsive group D MAPK gene from cotton, is involved in disease resistance and drought sensitivity. BMC Mol. Biol. 2011, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.G.; Zhang, L.M.; Xue, C.L.; Fang, H.; Zhao, J.; Liu, M. Genome-wide identification and analysis of MAPK and MAPKK gene family in Chinese jujube (Ziziphus jujuba Mill.). BMC Genomics 2017, 18, 855. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Wang, H.; Chen, P.; Fu, B.; Zhang, M.; Li, J.; Zheng, X.; Tan, B.; Feng, J. Combination of iTRAQ proteomics and RNA-seq transcriptomics reveals multiple levels of regulation in phytoplasma-infected Ziziphus jujuba Mill. Horticult. Res. 2017, 4, 17080. [Google Scholar] [CrossRef]
- Liu, M.J.; Zhou, J.Y.; Zhao, J.; Wang, J.R.; Liu, P.; Dai, L.; Wang, Y.X. An excellent new cultivar of Chinese jujube with high resistance to Jujube Witches Broom disease ‘Xingguang’. Acta Horticulturae Sinica. 2006, 33, 687. [Google Scholar]
- Takahashi, Y.; Soyano, T.; Kosetsu, K.; Sasabe, M.; Machida, Y. HINKEL kinesin, ANP MAPKKKs and MKK6/ANQ MAPKK, which phosphorylates and activates MPK4 MAPK, constitute a pathway that is required for cytokinesis in Arabidopsis thaliana. Plant Cell Physiol. 2010, 51, 1766–1776. [Google Scholar] [CrossRef]
- Soyano, T.; Nishihama, R.; Morikiyo, K.; Ishikawa, M.; Machida, Y. NQK1/NtMEK1 is a MAPKK that acts in the NPK1 MAPKKK-mediated MAPK cascade and is required for plant cytokinesis. Genes Dev. 2003, 17, 1055–1067. [Google Scholar] [CrossRef] [Green Version]
- Beck, M.; Komis, G.; Müller, J.; Menzel, D.; Samaj, J. Arabidopsis homologs of nucleus-and phragmoplast-localized kinase 2 and 3 and mitogen-activated protein kinase 4 are essential for microtubule organization. Plant Cell. 2010, 22, 755–771. [Google Scholar] [CrossRef] [PubMed]
- Beck, M.; Komis, G.; Ziemann, A.; Menzel, D.; Samaj, J. Mitogen-activated protein kinase 4 is involved in the regulation of mitotic and cytokinetic microtubule transitions in Arabidopsis thaliana. New Phytol. 2011, 189, 1069–1083. [Google Scholar] [CrossRef]
- Beckers, G.J.; Jaskiewicz, M.; Liu, Y.; Underwood, W.R.; He, S.Y.; Zhang, S.; Conrath, U. Mitogen-activated protein kinases 3 and 6 are required for full priming of stress responses in Arabidopsis thaliana. Plant Cell. 2009, 21, 944–953. [Google Scholar] [CrossRef]
- Romeis, T.; Ludwig, A.A.; Martin, R.D.G.; Jones, J. Calcium-dependent protein kinases play an essential role in a plant defence response. EMBO J. 2001, 20, 5556–5567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagami, H.; Pitzschke, A.; Hirt, H. Emerging MAP kinase pathways in plant stress signaling. Trends Plant Sci. 2005, 10, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Dai, Y.; Xiong, Y.; DeFraia, C.; Li, J.; Dong, X.; Mou, Z. Overexpression of Arabidopsis MAP kinase kinase 7 leads to activation of plant basal and systemic acquired resistance. Plant J. 2007, 52, 1066–1079. [Google Scholar] [CrossRef]
- Nicole, M.C.; Hamel, L.P.; Morency, M.J.; Beaudoin, N.; Ellis, B.E.; Séguin, A. MAP-ping genomic organization and organ-specific expression profiles of poplar MAP kinases and MAP kinase kinases. BMC Genom. 2006, 7, 223. [Google Scholar] [CrossRef]
- Pitzschke, A.; Schikora, A.; Hirt, H. MAPK cascade signalling networks in plant defence. Curr. Opin. Plant Biol. 2009, 12, 421–426. [Google Scholar] [CrossRef]
- Zhang, S.; Klessig, D.F. MAPK cascades in plant defense signaling. Trends Plant Sci. 2001, 6, 520–527. [Google Scholar] [CrossRef]
- Liu, X.; Liu, M.; Ning, Q.; Liu, G. Reverse-cleft in vitro micrografting of Ziziphus jujuba Mill. infected with jujube witches’ broom (JWB). Plant Cell Tiss Organ Cult. 2012, 108, 339–344. [Google Scholar] [CrossRef]
- Xue, C.; Liu, Z.; Dai, L.; Bu, J.; Liu, M.; Zhao, Z.; Jiang, Z.; Gao, W.; Zhao, J. Changing host photosynthetic, carbohydrate and energy metabolisms play important roles in phytoplasma infection. Phytopathology 2018, 108, 1067–1077. [Google Scholar] [CrossRef] [PubMed]
- Bu, J.; Zhao, J.; Liu, M. Expression stabilities of candidate reference genes for RT-qPCR in Chinese jujube (Ziziphus jujuba Mill.) under a variety of conditions. PLoS ONE 2016, 11, e0154212. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein–protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Up-Regulated Expression Times | Down-Regulated Expression Times | Total Times |
|---|---|---|---|
| ZjMPK1 | 0 | 1 | 1 |
| ZjMPK2 | 9 | 1 | 10 |
| ZjMPK3 | 0 | 4 | 4 |
| ZjMPK4 | 0 | 2 | 2 |
| ZjMPK5 | 2 | 2 | 4 |
| ZjMPK6 | 0 | 2 | 2 |
| ZjMPK7 | 0 | 4 | 4 |
| ZjMPK8 | 0 | 0 | 0 |
| ZjMPK9 | 7 | 0 | 7 |
| ZjMPK10 | 0 | 1 | 1 |
| ZjMKK1 | 0 | 0 | 0 |
| ZjMKK2 | 3 | 6 | 9 |
| ZjMKK3 | 0 | 0 | 0 |
| ZjMKK4 | 0 | 1 | 1 |
| In all | 21 | 24 | 45 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Zhao, Z.; Xue, C.; Wang, L.; Wang, L.; Feng, C.; Zhang, L.; Yu, Z.; Zhao, J.; Liu, M. Three Main Genes in the MAPK Cascade Involved in the Chinese Jujube-Phytoplasma Interaction. Forests 2019, 10, 392. https://doi.org/10.3390/f10050392
Liu Z, Zhao Z, Xue C, Wang L, Wang L, Feng C, Zhang L, Yu Z, Zhao J, Liu M. Three Main Genes in the MAPK Cascade Involved in the Chinese Jujube-Phytoplasma Interaction. Forests. 2019; 10(5):392. https://doi.org/10.3390/f10050392
Chicago/Turabian StyleLiu, Zhiguo, Zhihui Zhao, Chaoling Xue, Lixin Wang, Lili Wang, Chunfang Feng, Liman Zhang, Zhe Yu, Jin Zhao, and Mengjun Liu. 2019. "Three Main Genes in the MAPK Cascade Involved in the Chinese Jujube-Phytoplasma Interaction" Forests 10, no. 5: 392. https://doi.org/10.3390/f10050392
APA StyleLiu, Z., Zhao, Z., Xue, C., Wang, L., Wang, L., Feng, C., Zhang, L., Yu, Z., Zhao, J., & Liu, M. (2019). Three Main Genes in the MAPK Cascade Involved in the Chinese Jujube-Phytoplasma Interaction. Forests, 10(5), 392. https://doi.org/10.3390/f10050392