Soil Silicon Amendment Increases Phyllostachys praecox Cold Tolerance in a Pot Experiment

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bamboo and Soil
2.2. Bamboo Pot Experiment
Low Temperature Incubation
2.3. Si in Plant Measurement
2.4. Photosynthesis Parameter Measurement and Physiological Indicator Analysis
2.5. Observation of Bamboo Leaf Chloroplast Ultrastructure
2.6. Data Analysis
3. Results
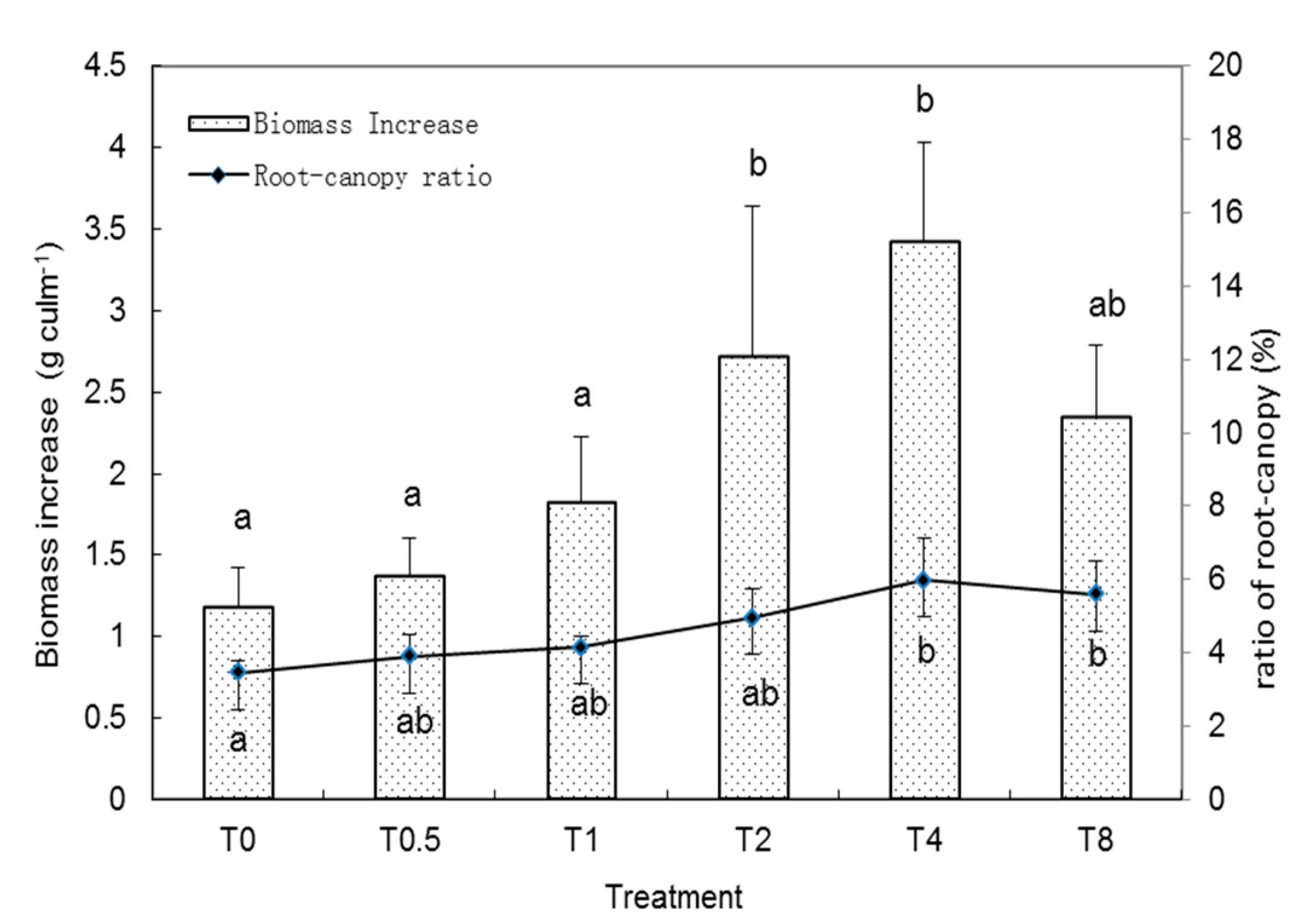
3.1. Bamboo Biomass Change with Si Amendment
3.2. Si Content in Bamboo Plants
3.3. Photosynthesis Parameters
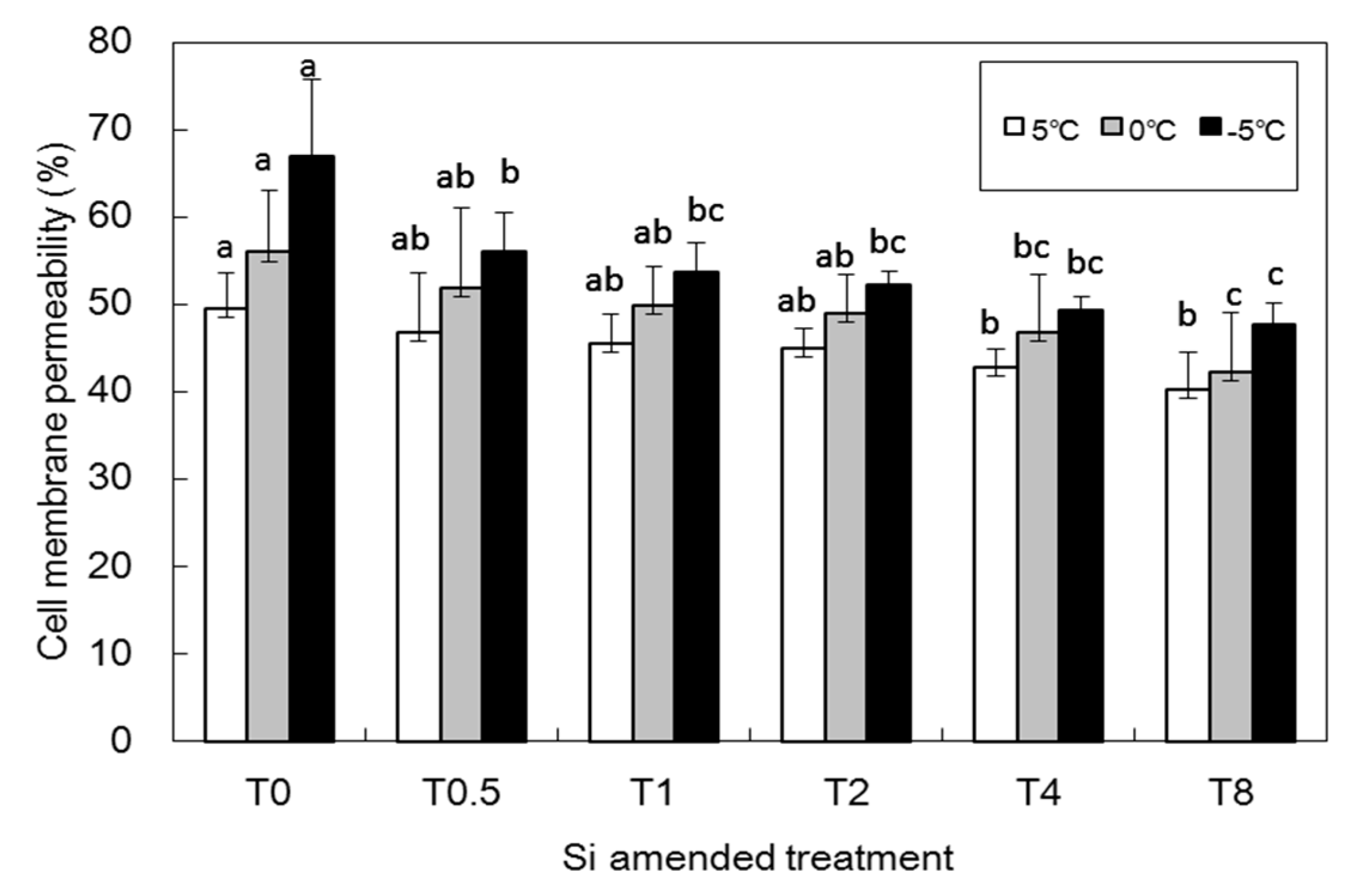
3.4. Physiological Indicators Treated with Low Temperature
3.5. Effect of Si Amendment on Bamboo Leaf Chloroplast Ultrastructure
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Epstein, E. Silicon. Annu. Rev. Plant Phys. 1999, 50, 641–664. [Google Scholar] [CrossRef] [PubMed]
- Goto, M.; Ehara, H.; Karita, S.; Takabe, K.; Ogawa, N.; Yamada, Y.; Ogawa, S.; Yahaya, M.S.; Morita, O. Protective effect of silicon on phenolic biosynthesis and ultraviolet spectral stress in rice crop. Plant Sci. 2003, 164, 349–356. [Google Scholar] [CrossRef]
- Agarie, S.; Hanaoka, N.; Ueno, O.; Miyazaki, A.; Kubota, F.; Agata, W.; Kaufman, P.B. Effects of silicon on tolerance to water deficit and heat stress in rice plants (Oryza sativa L.), monitored by electrolyte leakage. Plant Prod. Sci. 1998, 1, 96–103. [Google Scholar] [CrossRef]
- Liang, Y.C.; Zhang, W.H.; Chen, Q.; Liu, Y.L.; Ding, R.X. Effect of exogenous silicon (Si) on H+- ATPase activity, phospholipids and fluidity of plasma membrane in leaves of salt-stressed barley (Hordeum vulgare L.). Environ. Exp. Bot. 2006, 57, 212–219. [Google Scholar] [CrossRef]
- Liang, Y.C.; Shen, Q.R.; Shen, Z.G.; Ma, T. Effects of silicon on salinity tolerance of two barley cultivars. J. Plant Nutr. 1996, 19, 173–183. [Google Scholar] [CrossRef]
- Liang, Y.C. Effects of silicon on enzyme activity, and sodium, potassium and calcium concentration in barley under salt stress. Plant Soil. 1999, 209, 217–224. [Google Scholar] [CrossRef]
- Liang, Y.C.; Zhu, J.; Li, Z.J.; Chu, G.X.; Ding, Y.F.; Zhang, J.; Sun, W.C. Role of silicon in enhancing resistance to freezing stress in two contrasting winter wheat cultivars. Environ. Exp. Bot. 2008, 64, 286–294. [Google Scholar] [CrossRef]
- Liang, Y.C.; Yang, C.G.; Shi, H.H. Effects of silicon on growth and mineral composition of barley grown under toxic levels of aluminum. J. Plant Nutr. 2001, 24, 229–243. [Google Scholar] [CrossRef]
- Iwasaki, K.; Matsumura, A. Effect of silicon on alleviation of manganese toxicity in pumpkin (Cucurbita moschata Duch cv. Shintosa). Soil Sci. Plant Nutr. 1999, 45, 909–920. [Google Scholar] [CrossRef]
- Shi, Q.H.; Bao, Z.Y.; Zhu, Z.J.; He, Y.; Qian, Q.Q.; Yu, J.Q. Silicon-mediated alleviation of Mn toxicity in Cucumis sativus in relation to activities of superoxide dismutase and ascorbate peroxidase. Phytochemistry 2005, 66, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.J.; Wei, G.Q.; Li, J.; Qian, Q.Q.; Yu, J.Q. Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.). Plant Sci. 2004, 167, 527–533. [Google Scholar] [CrossRef]
- Liang, Y.C.; Zhang, W.; Chen, Q.; Ding, R. Effect of silicon on H+- ATPase and H+- PPase activity, fatty acid composition and fluidity of tonoplast vesiciles from roots of salt-stressed barley (Hordeum vulagare L.). Environ. Exp. Bot. 2004, 102, 34–39. [Google Scholar]
- Ding, Y.; Liang, Y.C.; Zhu, J.; Li, Z. Effects of silicon on plant growth, photosynthetic parameters and soluble sugar content in leaves of wheat under drought stress. Plant Nutr. Fert. Sci. 2007, 13, 471–478. (In Chinese) [Google Scholar]
- Yeo, A.R.; Flowers, S.A.; Rao, G.; Welfare, K.; Senanayake, N.; Flowers, T.J. Silicon reduces sodium uptake in rice (Oryza sativa L.) in saline conditions and this is accounted for by a reduction in the transpirational bypass flow. Plant Cell Environ. 1999, 22, 559–565. [Google Scholar] [CrossRef]
- Chen, W.; Yao, X.Q.; Cai, K.Z.; Chen, J.N. Silicon alleviates drought stress of rice plants by improving plant water status, photosynthesis and mineral nutrient absorption. Biol. Trace Elem. Res. 2011, 142, 67–76. [Google Scholar] [CrossRef]
- Steponkus, P.L. Role of the plasma membrane in freezing injury and cold acclimation. Annu. Rev. Plant. Physiol. 1984, 35, 543–584. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, J.Z.; Chow, W.S.; Sun, L.L.; Chen, J.W.; Chen, Y.J.; Peng, C.L. The influence of low temperature on photosynthesis and antioxidant enzymes in sensitive banana and tolerant plantain (Musa sp.) cultivars. Photosynthetica 2011, 49, 201–208. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef] [Green Version]
- Kawano, T. Roles of the reactive oxygen species-generating peroxidase reactions in plant defense and growth induction. Plant Cell Rep. 2003, 21, 829–837. [Google Scholar] [PubMed]
- Polidoros, A.N.; Scandalios, J.G. Role of hydrogen peroxide and different classes of antioxidants in the regulation of catalase and glutathione S-transferase gene expression in maize (Zea mays L.). Physiol. Plantarum. 1999, 106, 112–120. [Google Scholar] [CrossRef]
- Liu, J.J.; Lin, S.H.; Xu, P.L.; Wang, X.J.; Ba, J.G. Effects of exogenous silicon on the activities of antioxidant enzymes and lipid peroxidation in chilling-stressed cucumber leaves. Agric. Sci. China 2009, 8, 1075–1086. [Google Scholar] [CrossRef]
- Zhan, H.; Zhang, L.; Deng, L.; Niu, Z.; Li, M.; Wang, C.; Wang, S. Physiological and anatomical response of foliar silicon application to Dendrocalamus brandisii plantlet leaves under chilling. Acta Physiol. Plant. 2018, 40, 208. [Google Scholar] [CrossRef]
- Xu, C.X.; Liu, Y.L. Si absorption, transport and accumulation in plants. Acta Bot. Boreali Occid. Sin. 2006, 26, 1071–1078. (In Chinese) [Google Scholar]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N. Si uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef]
- Ma, J.F. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Zhou, B.; Wang, X.; Cao, Y.; Ge, X.; Gu, L.; Meng, J. Damage assessment to subtropical forests following the 2008 Chinese ice storm. iForest 2017, 10, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Zhuang, S.Y.; He, J.C.; Gui, R.Y. Intensive management measures: Effects on freeze injury of Phyllostachys praecox. Chin. Agric. Sci. Bull. 2018, 34, 95–100. (In Chinese) [Google Scholar]
- He, D.; Song, H.; Qiu, Z.; Shi, G.; Chen, J. Analysis on effect of snow disaster on Phyllostachys praecox cv. Prevernalis stand and countermeasures. J. Zhejiang For. Sci. Tech. 2008, 5, 54–56. (In Chinese) [Google Scholar]
- Liang, Y.C.; Sun, W.; Zhu, Y.G.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 42. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Tamai, K.; Ichii, M.; Wu, G.F. A rice mutant defective in Si uptake. Plant Physiol. 2002, 30, 2111–2117. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; Fu, Z.; Zhang, L.; Zhang, Z.; Zhang, J. The positive effect of different 24-epiBL pretreatments on salinity tolerance in Robinia pseudoacacia L. seedlings. Forests 2019, 10, 4. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Maehly, A. The assay of catalases and peroxidases. Methods Biochem. Anal. 1954, 357–424. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalase and peroxidases. Methods Enzymol. 1955, 1, 764–775. [Google Scholar]
- Xu, P.L.; Guo, Y.K.; Bai, J.G.; Shang, L.; Wang, X.J. Effects of long-term chilling on ultrastructure and antioxidant activity in leaves of two cucumber cultivars under low light. Physiol. Plant. 2008, 132, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Huang, X.; Chen, Y. Experimental Course of Plant Physiology, 2nd ed.; Higher Education Press: Beijing, China, 2009; pp. 129–139. (In Chinese) [Google Scholar]
- Tian, H.Q.; Kuang, A.; Musgrave, M.E.; Russell, S.D. Calcium distribution in fertile and sterile anther of a photoperiod-sensitive genic male-sterile rice. Planta 1998, 204, 183–192. [Google Scholar] [CrossRef]
- Jiang, P.; Ye, Z.; Xu, Q. Effect of mulching on soil chemical properties and enzyme activities in bamboo plantation of phyllostachy praecox. Commun. Soil Sci. Plan. 2002, 33, 11. [Google Scholar] [CrossRef]
- Gu, D.X.; Chen, S.L.; Zheng, W.M.; Mao, X.Q. Review of the ecological adaptability of bamboo. J. Bamboo Res. 2010, 29, 17–23. (In Chinese) [Google Scholar]
- Li, Z.; Lin, P.; He, J.; Yang, Z.; Lin, Y. Silicon’s organic pool and biological cycle in moso bamboo community of Wuyishan Biosphere Reserve. J. Zhejiang Univ.-SC B 2006, 7, 849–857. [Google Scholar] [CrossRef]
- Ding, T.P.; Zhou, J.X.; Wan, D.F.; Chen, Z.Y.; Wang, C.Y.; Zhang, F. Silicon isotope fractionation in bamboo and its significance to the biogeochemical cycle of silicon. Geochim Cosmochim Ac. 2008, 72, 1395. [Google Scholar] [CrossRef]
- Detmann, K.C.; Araújo, W.L.; Martins, S.C.; Sanglard, L.M.; Reis, J.V.; Detmann, E.; Rodrigues, F.A.; Nunes-Nesi, A.; Fernie, A.R.; DaMatta, F.M. Silicon nutrition increases grain yield, which, in turn, exerts a feed-forward stimulation of photosynthetic rates via enhanced mesophyll conductance and alters primary metabolism in rice. New Phytol. 2012, 196, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Tamai, K.; Ma, J.F. Reexamination of silicon effects on rice growth and production under field conditions using a low silicon mutant. Plant Soil. 2008, 307, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Barbour, M.M.; Brendel, O.; Cabrera, H.M.; Carriqui, M.; Diaz-Espejo, A.; Douthe, C.; Dreyer, E.; Ferrio, J.P.; Gago, J.; et al. Mesophyll diffusion conductance to CO2: An unappreciated central player in photosynthesis. Plant Sci. 2012, 193, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Alscher, R.G.; Donahue, J.L.; Cramer, C.L. Reactive oxygen species and antioxidants: Relationship in green cells. Physiol. Plant. 1997, 100, 224–233. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Bioch. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Baek, K.H.; Skinner, D.Z. Production of reactive oxygen species by freezing stress, and the protective roles of antioxidant enzymes in plants. J. Agric. Chem. Environ. 2012, 1, 34–40. [Google Scholar] [CrossRef]
- Saeidnejad, A.H.; Pouramir, F.; Naghizedek, M. Improving chilling tolerance of maize seedlings under cold conditions by spermine application. Not Sci. Biol. 2012, 4, 110–117. [Google Scholar] [CrossRef]
- Moradtalab, N.; Weinmann, M.; Walker, F.; Höglinger, B.; Ludewig, U.; Neumann, G. Silicon improves chilling tolerance during early growth of maize by effects on micronutrient homeostasis and hormonal balances. Front Plant Sci. 2018, 9, 420. [Google Scholar] [CrossRef]
- Neto, A.D.D.; Prisco, J.T.; Eneas, J.; de Abreu, C.E.B.; Gomes, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Foyer, C.H.; Lopez-Delgado, H.; Dat, J.F.; Scott, I.M. Hydrogen peroxide and glutathione-associated mechanisms of acclamatory stress tolerance and signaling. Physiol. Plant. 1997, 100, 241–254. [Google Scholar] [CrossRef]
- Zgallai, H.; Steppe, K.; Lemeur, R. Effects of different levels of water stress on leaf water potential, stomatal resistance, protein and chlorophyll content and certain anti-oxidative enzymes in tomato plants. J. Integr. Plant Biol. 2006, 48, 679–685. [Google Scholar] [CrossRef]
- Sgherri, C.L.M.; Loggini, B.; Puliga, S.; Navari-Izzo, F. Antioxidant system in Sporobolus stapfianus: Changes in response to desiccation and rehydration. Phytochemistry 1994, 35, 561–565. [Google Scholar] [CrossRef]
- Wang, L.J.; Huang, W.D.; Li, J.Y.; Liu, Y.F.; Shi, Y.L. Peroxidation of membrane lipid and Ca2+ homeostasis in grape mesophyll cells during the process of cross-adaptation to temperature stresses. Plant Sci. 2004, 167, 71–77. [Google Scholar] [CrossRef]
- Ji, S.X.; Dai, S.J.; Liu, W. The advances of plants in response and adaption to low temperature stress. Chin. Bull Life Sci. 2010, 22, 1013–1019. (In Chinese) [Google Scholar]
- He, Y.; Xiao, H.; Wang, H.; Chen, Y.; Yu, M. Effect of silicon on chilling-induced changes of solutes, antioxidants, and membrane stability in seashore paspalum turfgrass. Acta Physiol. Plant. 2010, 32, 487–494. [Google Scholar] [CrossRef]
- Chen, B.; Shi, Y.; Chen, Z. Studies on the effect of different P, K, Si fertilizer of corn cold-resistant. Chin. Agric. Sci. Bull. 2011, 27, 85–89. (In Chinese) [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Leaf | Stem | Rhizome | Root |
|---|---|---|---|---|
| T0 | 27.77 ± 4.70 a1 | 11.18 ± 1.53 a | 3.50 ± 0.72 a | 3.07 ± 0.99 a |
| T0.5 | 31.93 ± 3.68 a | 11.35 ± 1.40 a | 3.31 ± 0.81 a | 3.69 ± 1.18 a |
| T1 | 43.19 ± 3.25 b | 16.71 ± 4.37 ab | 4.82 ± 0.95 a | 5.33 ± 0.71 b |
| T2 | 48.88 ± 2.13 b | 18.88 ± 3.69 b | 5.64 ± 1.10 ab | 6.14 ± 0.74 bc |
| T4 | 68.95 ± 10.11 c | 22.60 ± 3.71 c | 7.92 ± 2.51 bc | 7.16 ± 1.11 c |
| T8 | 66.26 ± 9.20 c | 23.50 ± 5.57 c | 9.13 ± 3.25 c | 7.63 ± 1.19 c |
| a | b | c | R2 | P | |
|---|---|---|---|---|---|
| Leaf | −1.290 | 15.31 | 26.61 | 0.9828 | 0.003 |
| Stem | −0.3743 | 4.554 | 10.92 | 0.9566 | 0.009 |
| Rhizome | −0.1003 | 1.557 | 3.122 | 0.9789 | 0.003 |
| Root | −0.1309 | 1.584 | 3.272 | 0.9585 | 0.008 |
| Treatment | Photosynthesis Rate (Pn) (μmol CO2• m−2 s−1) | Transpiration Rate (Tr) (mmol H2O•m−2 s−1) | Water Use Efficiency (WUE) (μmol CO2•mmol−1 H2O) | CO2 of Intercellular Space (Ci) (μL L−1) | Stomatal Conductance (Gs) (mmol•m−2 s−1) |
|---|---|---|---|---|---|
| T0 | 7.03 ± 2.69 a | 3.74 ± 1.26 a | 2.80 ± 1.83 a | 224.33 ± 45.89 a | 109.0 ± 14.91 a |
| T0.5 | 7.01 ± 0.65 a | 2.64 ± 0.89 b | 2.38 ± 0.58 ab | 240.62 ± 40.19 a | 99.4 ± 28.6 a |
| T1 | 8.11 ± 3.97 ab | 2.5 ± 0.32 b | 3.84 ± 1.58 ab | 241.12 ± 45.21 a | 97.63 ± 12.83 a |
| T2 | 9.57 ± 2.6 abc | 2.58 ± 0.89 b | 3.94 ± 1.17 ab | 240.19 ± 47.57 a | 111.07 ± 30.65 a |
| T4 | 10.11 ± 2.55 bc | 2.27 ± 0.6 b | 4.55 ± 1.12 b | 228.47 ± 19.73 a | 103.96 ± 17.01 a |
| T8 | 11.89 ± 1.63 c | 1.96 ± 0.75 b | 7.39 ± 3.79 c | 230.99 ± 37.23 a | 118.7 ± 38.06 a |
| a | b | c | R2 | P | |
|---|---|---|---|---|---|
| SOD | −1.783 | 23.266 | 633.6 | 0.89389 | 0.016 |
| POD | −0.7196 | 9.935 | 112.1 | 0.96943 | 0.002 |
| CAT | −0.3301 | 6.390 | 27.49 | 0.98354 | 9.82 × 10−4 |
| MDA | 0.05001 | −1.202 | 27.95 | 0.97371 | 0.002 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, Z.Z.; Zhuang, S.Y.; Li, Q.; Gui, R.Y. Soil Silicon Amendment Increases Phyllostachys praecox Cold Tolerance in a Pot Experiment. Forests 2019, 10, 405. https://doi.org/10.3390/f10050405
Qian ZZ, Zhuang SY, Li Q, Gui RY. Soil Silicon Amendment Increases Phyllostachys praecox Cold Tolerance in a Pot Experiment. Forests. 2019; 10(5):405. https://doi.org/10.3390/f10050405
Chicago/Turabian StyleQian, Zhuang Zhuang, Shun Yao Zhuang, Qiang Li, and Ren Yi Gui. 2019. "Soil Silicon Amendment Increases Phyllostachys praecox Cold Tolerance in a Pot Experiment" Forests 10, no. 5: 405. https://doi.org/10.3390/f10050405
APA StyleQian, Z. Z., Zhuang, S. Y., Li, Q., & Gui, R. Y. (2019). Soil Silicon Amendment Increases Phyllostachys praecox Cold Tolerance in a Pot Experiment. Forests, 10(5), 405. https://doi.org/10.3390/f10050405