Mortality of Different Populus Genotypes in Recently Established Mixed Short Rotation Coppice with Robinia pseudoacacia L.

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
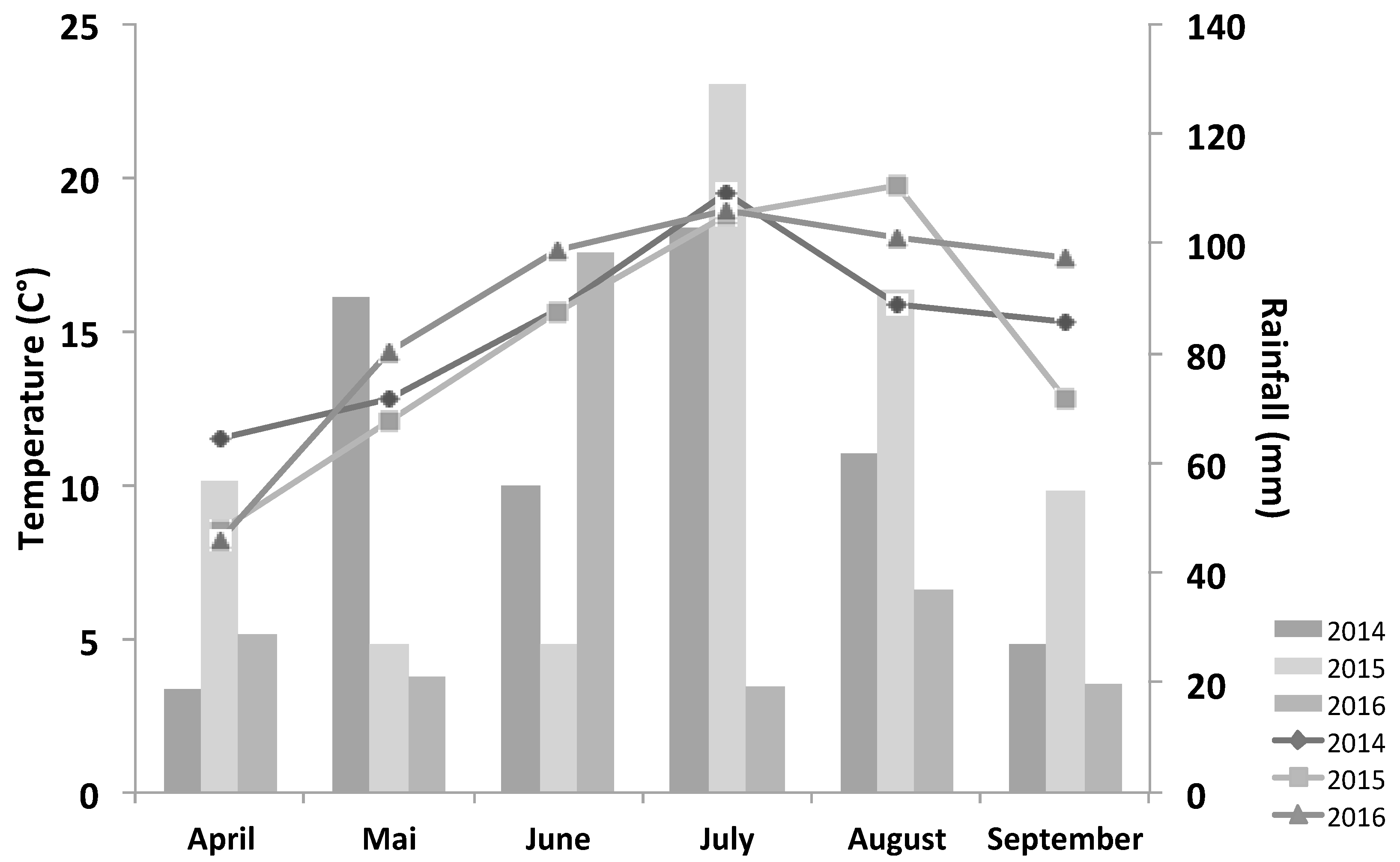
2.1. Study Sites and Species
2.2. Survival and Competition Survey
2.3. Statistical Data Analyses
3. Results
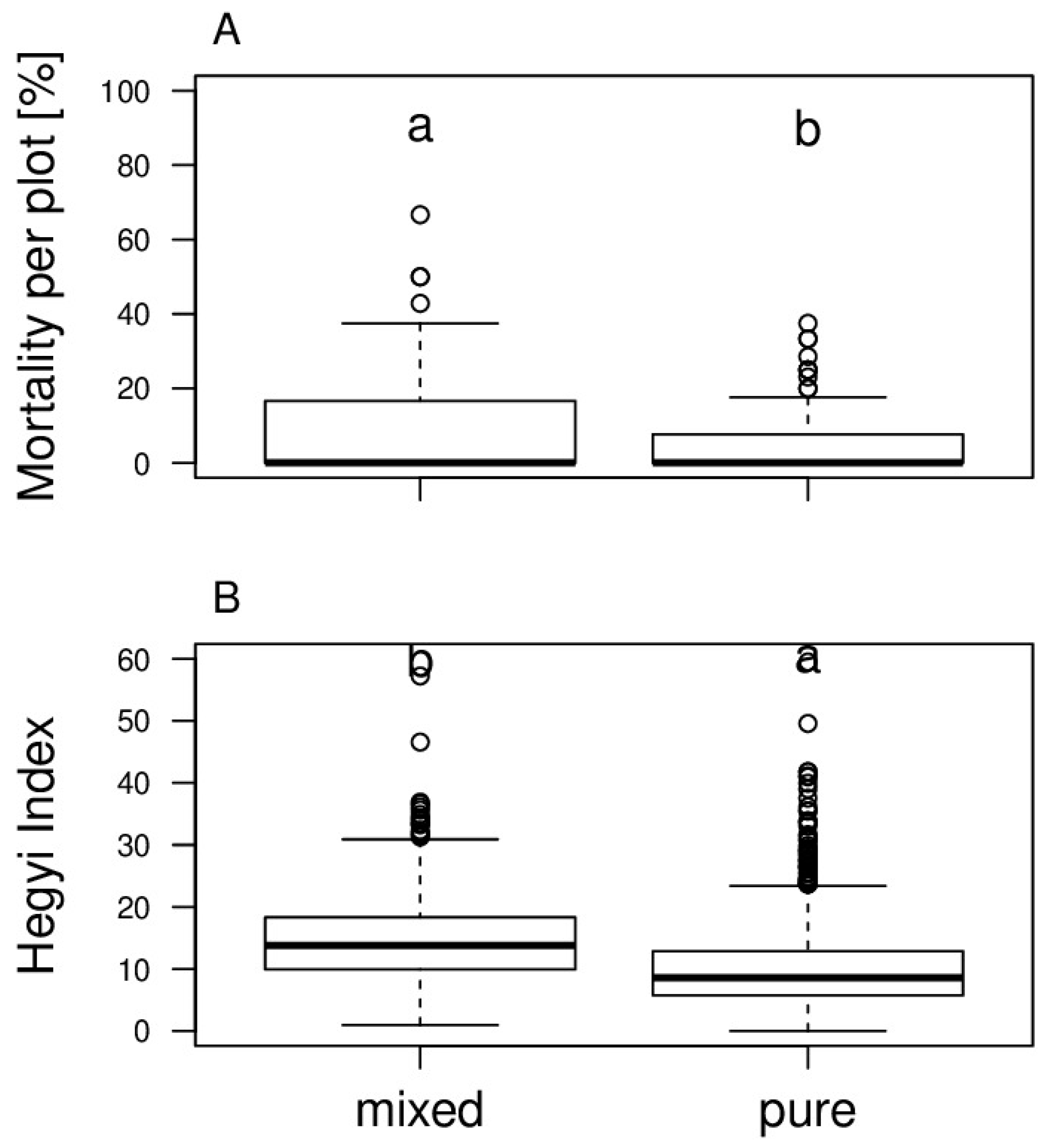
3.1. Mortality of Monocultures and Mixtures across Genotypes
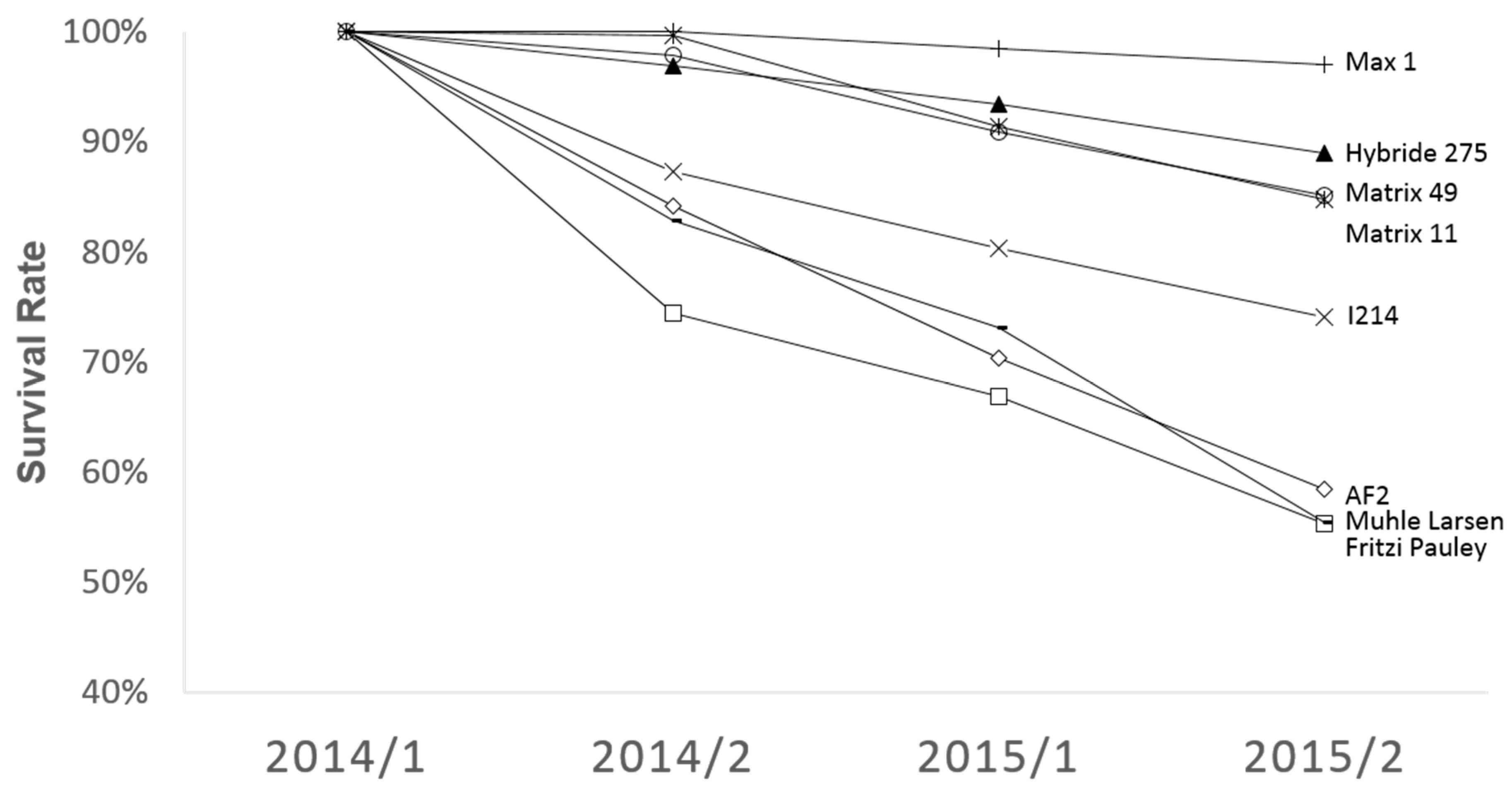
3.2. Mortality of the Different Poplar Genotypes
3.3. Competition Induced Mortality
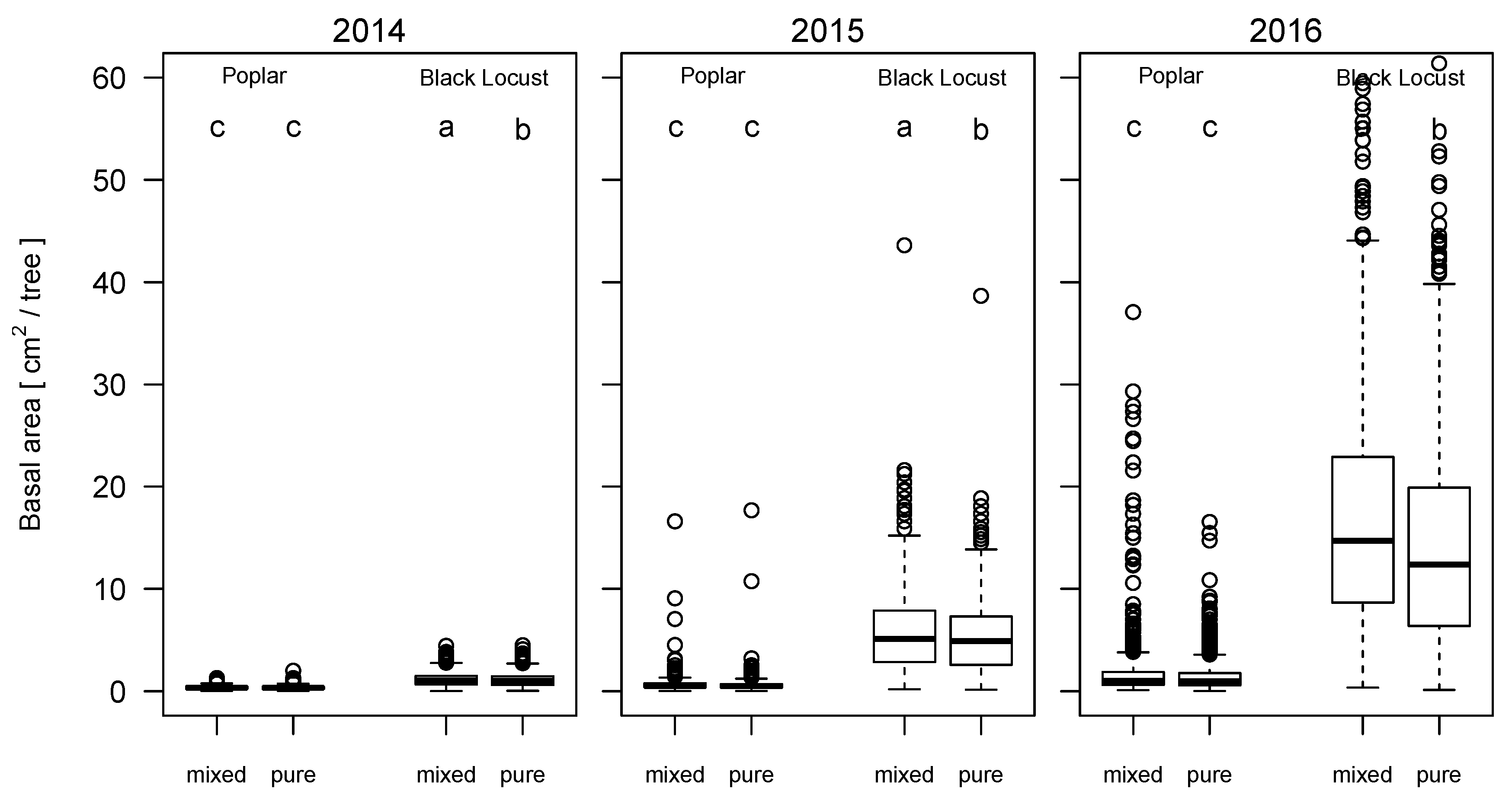
3.4. Growth Performance of Poplar in Pure and Mixed Stands
4. Discussion
4.1. Genotypical Variation
4.2. Mixing Effects on Mortality
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Demirbaş, A. Biomass resource facilities and biomass conversion processing for fuels and chemicals. Energy Convers. Manag. 2001, 42, 1357–1378. [Google Scholar] [CrossRef]
- Butler Manning, D.; Bemmann, A.; Bredemeier, M.; Lamersdorf, N.; Ammer, C. Bioenergy from Dendromass for the Sustainable Development of Rural Areas; Wiley-VCH: Weinheim, Germany, 2015. [Google Scholar]
- Shortall, O.K. Marginal land” for energy crops: Exploring definitions and embedded assumptions. Energy Policy 2013, 62, 19–27. [Google Scholar] [CrossRef]
- Ehlting, B.; Dluzniewska, P.; Dietrich, H.; Selle, A.; Teuber, M.; Hänsch, R.; Nehls, U.; Polle, A.; Schnitzler, J.; Rennenberg, H. Interaction of nitrogen nutrition and salinity in grey poplar (Populus tremula × alba). Plant. Cell Environ. 2007, 30, 796–811. [Google Scholar] [CrossRef]
- Euring, D.; Löfke, C.; Teichmann, T.; Polle, A. Nitrogen fertilization has differential effects on N allocation and lignin in two Populus species with contrasting ecology. Trees 2012, 26, 1933–1942. [Google Scholar] [CrossRef]
- Kumazawa, K. Nitrogen fertilization and nitrate pollution in groundwater in Japan: Present status and measures for sustainable agriculture. Nutr. Cycl. Agroecosyst. 2002, 63, 129–137. [Google Scholar] [CrossRef]
- Sebilo, M.; Mayer, B.; Nicolardot, B.; Pinay, G.; Mariotti, A. Long-term fate of nitrate fertilizer in agricultural soils. Proc. Natl. Acad. Sci. USA. 2013, 110, 18185–18189. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.L.; Tian, Z.X.; Zhang, N.; Li, X.Q. Nitrate pollution of groundwater in northern China. Agric. Ecosyst. Environ. 1996, 59, 223–231. [Google Scholar] [CrossRef]
- Binkley, D. Mixtures of nitrogen2-fixing and non-nitrogen2-fixing tree species. In The Ecology of Mixed-Species Stands of Trees; Cannell, M.G.R., Malcolm, D.C., Robertson, P.A., Eds.; Blackwell Scientific: Boston, MA, USA, 1992; pp. 99–123. [Google Scholar]
- Vandermeer, J.H. The Ecology of Intercropping; Cambridge University Press: Cambridge, NY, USA, 1989. [Google Scholar]
- Veste, M.; Böhm, C.; Quinkenstein, A.; Freese, D. Biologische Stickstoff-Fixierung der Robinie. AFZ/Wald 2013, 2, 40–42. [Google Scholar]
- Ammer, C. Diversity and forest productivity in a changing climate. New Phytol. 2019, 221, 50–66. [Google Scholar]
- Pretzsch, H. Individual Tree Structure and Growth in Mixed Compared with Monospecific Stands. In Mixed-Species Forests; Pretzsch, H., Forrester, D.I., Bauhus, J., Eds.; Springer: Berlin, Germany, 2017; pp. 271–336. [Google Scholar]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L.; Vanclay, J.K. Mixed-Species Plantations of Eucalyptus with Nitrogen-Fixing Trees: A Review. For. Ecol. Manag. 2006, 233, 211–230. [Google Scholar] [CrossRef]
- Hansen, E.A.; Dawson, J.O. Effect of Alnus glutinosa on hybrid Populus height growth in a short-rotation intensively cultured plantation. For. Sci. 1982, 28, 49–59. [Google Scholar]
- Oliveira, N.; del Río, M.; Forrester, D.I.; Rodríguez-Soalleiro, R.; Pérez-Cruzado, C. Mixed short rotation plantations of Populus alba and Robinia pseudoacacia for biomass yield. For. Ecol. Manag. 2018, 410, 48–55. [Google Scholar] [CrossRef]
- Rédei, K.; Veperd, I.; Meilby, H. Stand structure and growth of mixed white poplar (Populus alba L.) and black locust (Robinia pseudoacacia L.) plantations in Hungary. Acta Silv. Lign. Hung. 2006, 2, 23–32. [Google Scholar]
- Rédei, K. Black Locust (Robinia pseudoacacia L.) Growing in Hungary; ERTI Forest Research Institute: Sárvar, Hungary, 2013. [Google Scholar]
- Reeg, T.; Bemmann, A.; Konold, W.; Murach, D.; Spiecker, H. Anbau und Nutzung von Bäumen auf landwirtschaftlichen Flächen; Wiley-VCH: Weinheim, Germany, 2009. [Google Scholar]
- Meyer-Münzer, B.; Grotehusmann, H.; Vor, T. Robinie (Robinia pseudoacacia L.). In Potenziale und Risiken eingeführter Baumarten; Universitätsverlag Göttingen: Göttingen, Germany, 2015; pp. 277–293. [Google Scholar]
- Forrester, D.; Bauhus, J.; Khanna, P.K. Growth dynamics in a mixed-species plantation of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Manag. 2004, 193, 81–95. [Google Scholar] [CrossRef]
- Helbig, C.; Müller, M. Abiotische und biotische Schadfaktoren. In Anbau und Nutzung von Bäumen auf landwirtschaftlichen Flächen Kurzumtriebsplantagen; Reeg, T., Bemmann, A., Konold, W., Murach, D., Spiecker, H., Eds.; Wiley-VCH: Weinheim, Germany, 2009; pp. 83–97. [Google Scholar]
- Schildbach, M.; Landgraf, D.; Böcker, L. Steckhölzer zur Begründung von Kurzumtriebsplantagen. AFZ/Wald 2008, 18, 992–993. [Google Scholar]
- Schildbach, M.; Hofmann, M.; Wolf, H. Anlage und Etablierung von Kurzumtriebsplantagen. In AGROWOOD-Kurzumtriebsplantagen in Deutschland und europäische Perspektiven; Bemmann, A., Knust, C., Eds.; Weißensee Verlag: Dresden, Germany, 2010; pp. 65–73. [Google Scholar]
- Knust, C.; Schua, K.; Wolf, H.; Feger, K.-H. Natürliche Rahmenbedingungen. In Energieholzplantagen in der Landwirtschaft; Bemmann, A., Butler Manning, D., Eds.; Agrimedia Erling Verlag: Clense, Germany, 2013; pp. 19–53. [Google Scholar]
- Schirmer, R.; Haikali, A. Sortenprüfung von Pappelhybriden für Energiewälder. LWF Wissen 2014, 74, 106–118. [Google Scholar]
- Spangenberg, G.; Hein, S. Herbizidfreie Begründung von Kurzumtriebsflächen. AFZ/Wald 2011, 10, 18–20. [Google Scholar]
- Deutscher Wetterdienst-Climate Data Center. Available online: www.DWD.de (accessed on 10 December 2018).
- Hegyi, F. A simulation model for managing Jack-pine stands. In Growth Models for Tree and Stand Simulations; Fries, J., Ed.; Royal College of Forestry: Stockholm, Sweden, 1974; pp. 74–90. [Google Scholar]
- Burkhart, H.E.; Tomé, M. Modeling Forest Trees and Stands; Springer: Heidelberg, Germany, 2012. [Google Scholar]
- Therneau, T. A Package for Survival Analysis in S. Available online: https://CRAN.R-project.org/package=survival (accessed on 8 February 2018).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Crawley, M.J. The R Book; Wiley: West Sussex, UK, 2007. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Stanton, B.J.; Serapiglia, M.J.; Smart, L.B. The domestication and conservation of Populus and Salix genetic resources. In Poplars and Willows: Trees for Society and Environment; Isebrands, J.G., Richardson, J., Eds.; FAO/CABI: Rome, Italy, 2014. [Google Scholar]
- Broeckx, L.S.; Verlinden, M.S.; Ceulemans, R. Establishment and two-year growth of a bio-energy plantation with fast-growing Populus trees in Flanders (Belgium): Effects of genotype and former land use. Biomass Bioenergy 2012, 42, 151–163. [Google Scholar] [CrossRef]
- Ceulemans, R.; Scarascia-Mugnozza, G.; Wiard, B.M.; Braatne, J.H.; Hinckley, T.M.; Stettler, R.F. Production physiology and morphology of Populus species and their hybrids grown under short rotation. I. Clonal comparisons of 4-year growth and phenology. Can. J. For. Res. 1992, 22, 1937–1948. [Google Scholar] [CrossRef]
- Grotehusmann, H.; Janssen, A.; Haikali, A.; Hartmann, K.-U.; Hüller, W.; Karopka, M.; Schildbach, M.; Schirmer, R.; Schuppelius, T.; Töpfner, K. Pappelsortenprüfung im Projekt FastWOOD. Forstarchiv 2015, 86, 67–79. [Google Scholar]
- Verlinden, M.S.; Broeckx, L.S.; van den Bulcke, J.; van Acker, J.; Ceulemans, R. Comparative study of biomass determinants of 12 poplar (Populus) genotypes in a high-density short-rotation culture. For. Ecol. Manag. 2013, 307, 101–111. [Google Scholar] [CrossRef]
- Melnikova, N.V.; Borkhert, E.V.; Snezhkina, A.V.; Kudryavtseva, A.V.; Dmitriev, A.A. Sex-Specific Response to Stress in Populus. Front. Plant Sci. 2017, 8, 1827. [Google Scholar] [CrossRef] [PubMed]
- Garavillon-Tournayre, M.; Gousset-Dupont, A.; Gautier, F.; Benoit, P.; Conchon, P.; Souchal, R.; Lopez, D.; Petel, G.; Venisse, J.S.; Bastien, C.; et al. Integrated drought responses of black poplar: How important is phenotypic plasticity? Physiol. Plant 2018, 163, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Yıldırım, K.; Kaya, Z. Gene regulation network behind drought escape, avoidance and tolerance strategies in black poplar (Populus nigra L.). Plant Physiol. Biochem. 2017, 115, 183–199. [Google Scholar] [CrossRef]
- Löf, M. Establishment and growth in seedlings of Fagus sylvatica and Quercus robur: Influence of interference from herbaceous vegetation. Can. J. For. Res. 2000, 30, 855–864. [Google Scholar] [CrossRef]
- Bäumler, W. Unkrautbekämpfung und Mäuse in Forstkulturen. Anz. Schändl.kd. Pflanzenschutz Umweltschutz 1992, 65, 143–144. [Google Scholar] [CrossRef]
- Radosevich, S.R.; Hibbs, D.E.; Ghersa, C.M. Effects of species mixtures and growth and stand development of Douglas-fir and red alder. Can. J. For. Res. 2006, 36, 768–782. [Google Scholar] [CrossRef]
- Forrester, D.; Bauhus, J.; Cowie, A. On the success and failure of mixed-species tree plantations: Lessons learned from a model system of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Manag. 2005, 209, 147–155. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Genotype | Parentage |
|---|---|
| AF2 | P. deltoides × P. nigra |
| Fritzi Pauley | P. trichocarpa |
| Hybride 275 | P. maximowiczii × P. trichocarpa |
| I214 | P. deltoides × P. nigra |
| Matrix 11 | P. maximowiczii × P. trichocarpa |
| Matrix 49 | P. maximowiczii × P. trichocarpa |
| Max 1 | P. nigra × P. maximowiczii |
| Muhle Larsen | P. trichocarpa |
| Stand Type | AF2 | Fritzi Pauley | Hybride 275 | I214 | Matrix 11 | Matrix 49 | Max 1 | Muhle Larsen | |
|---|---|---|---|---|---|---|---|---|---|
| 2014 | mixed | 0.17 ± 0.18 ab | 0.26 ± 0.27 b | 0.052 ± 0.094 ac | 0.10 ± 0.13 bc | 0.00 ± 0.00 bc | 0.029 ± 0.084 c | 0.00 ± 0.00 bc | 0.15 ± 0.10 bc |
| pure | 0.14 ± 0.15 bc | 0.24 ± 0.24 b | 0.022 ± 0.061 c | 0.14 ± 0.18 bc | 0.073 ± 0.021 ac | 0.017 ± 0.032 c | 0.00 ± 0.00 bc | 0.20 ± 0.21 ab | |
| 2015 | mixed | 0.34 ± 0.24 ab | 0.33 ± 0.32 ab | 0.085 ± 0.17 bc | 0.17 ± 0.19 ac | 0.14 ± 0.17 bc | 0.14 ± 0.13 bc | 0.034 ± 0.056 c | 0.41 ± 0.19 a |
| pure | 0.36 ± 0.28 a | 0.31 ± 0.23 a | 0.091 ± 0.13 b | 0.14 ± 0.15 ab | 0.16 ± 0.16 ab | 0.13 ± 0.16 ab | 0.025 ± 0.035 b | 0.31 ± 0.22 a | |
| 2016 | mixed | 0.10 ± 0.13 ab | 0.11 ± 0.15 ab | 0.043 ± 0.069 ab | 0.13 ± 0.15 ab | 0.066 ± 0.087 ab | 0.063 ± 0.093 ab | 0.032 ± 0.054 b | 0.19 ± 0.17 a |
| pure | 0.056 ± 0.092 ab | 0.025 ± 0.047 ab | 0.014 ± 0.031 ab | 0.071 ± 0.096 ab | 0.069 ± 0.11 ab | 0.042 ± 0.063 ab | 0.011 ± 0.029 b | 0.091 ± 0.11 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebola-Lichtenberg, J.; Schall, P.; Annighöfer, P.; Ammer, C.; Leinemann, L.; Polle, A.; Euring, D. Mortality of Different Populus Genotypes in Recently Established Mixed Short Rotation Coppice with Robinia pseudoacacia L. Forests 2019, 10, 410. https://doi.org/10.3390/f10050410
Rebola-Lichtenberg J, Schall P, Annighöfer P, Ammer C, Leinemann L, Polle A, Euring D. Mortality of Different Populus Genotypes in Recently Established Mixed Short Rotation Coppice with Robinia pseudoacacia L. Forests. 2019; 10(5):410. https://doi.org/10.3390/f10050410
Chicago/Turabian StyleRebola-Lichtenberg, Jessica, Peter Schall, Peter Annighöfer, Christian Ammer, Ludger Leinemann, Andrea Polle, and Dejuan Euring. 2019. "Mortality of Different Populus Genotypes in Recently Established Mixed Short Rotation Coppice with Robinia pseudoacacia L." Forests 10, no. 5: 410. https://doi.org/10.3390/f10050410
APA StyleRebola-Lichtenberg, J., Schall, P., Annighöfer, P., Ammer, C., Leinemann, L., Polle, A., & Euring, D. (2019). Mortality of Different Populus Genotypes in Recently Established Mixed Short Rotation Coppice with Robinia pseudoacacia L. Forests, 10(5), 410. https://doi.org/10.3390/f10050410