1. Introduction
Plant architecture is the result of long-term evolution and natural selection, which involve complex regulatory processes based on genetic and environmental interactions [
1]. According to the different growth angles and branching directions, plant architecture can be roughly classified as standard, weeping, pillar, upright, brachytic, and dwarf [
2]. Plant architecture traits have important application value for the improvement of crop production; for example, the cultivation of dwarf or semi-dwarf horticultural or agronomic crops improves yield and production efficiency because mechanized management is more conducive. The discovery and utilization of dwarf genes in rice, which greatly promoted grain production, led to the first “green revolution” in agriculture, demonstrating the importance of genetic resources [
3,
4]. The genetic regulation of plant architecture traits in woody plants is more complex than in herbaceous plants and plant architecture traits are susceptible to external environmental conditions [
5]. Many studies have been performed on plant architecture for extreme plant types of trees, including pillar and weeping peach [
6], dwarf and compact peach (
Prunus persica L.) [
7], weeping
Prunus mume [
8], weeping
Cercis chinensis [
9], and columnar and weeping apple (
Malus ×
domestica) [
10,
11].
Molecular marker technology can reflect the differences between DNA sequences for different species and is widely used in plant genetic diversity analysis, genetic map construction, map-based cloning, and marker-assisted selective breeding [
12,
13,
14]. Individual hybrids with target traits can be selected at the seedling stage using molecular marker-assisted selection, which reduces resource waste and breeding cost and speeds up the breeding process [
15]. Bulked segregant analysis (BSA) is derived from near-isogenic line analysis, which was reported for the first time in 1991 [
16]. SSR markers are widely used in relationship analysis, genetic diversity analysis, mapping quantitative trait locus (QTL), and so on. Four expressed sequence tag (EST)-SSR marker loci closely linked to the dwarf trait in pear were found by BSA technology. In
Actinidia chinensis Planch, an EST-SSR marker was screened and the genetic distance between the marker and dwarf gene was 8.8 cM [
17]. Three selected EST-SSR markers were used to determine genetic structure in 29 cultivars and were used for fruit color selection in
Prunus salicina breeding [
18]. In addition, DNA markers associated with the dwarf trait from
Brassica napus [
19],
Prunus persica [
20], and
Avena sativa [
21] have been studied.
Lagerstroemia L. (crape myrtle) belongs to the Lythraceae family, which includes at least 50 species of deciduous or evergreen shrubs or trees native to Southeast Asia [
22]. China is located in the center of the worldwide
Lagerstroemia distribution and origin [
23]. Crape myrtle was first cultivated as an ornamental species in China approximately 1800 years ago [
24]. Species in this genus are highly valued in landscaping for their graceful plant architecture, long-lasting flowering period, and colorful flowers during the summer [
25]. When exposed to natural conditions, crape myrtle is a diploid plant (2n = 2x = 48) [
26]. Crape myrtle is susceptible to powdery mildew; therefore, the initial purpose for hybrid breeding by interspecific hybridization between
L. indica and
L. fauriei was for disease resistance and then a series of excellent varieties with various plant architecture types and colorful flowers was selected from the resistant hybrid offspring (‘Pocomoke”) [
27,
28,
29].
Lagerstroemia species are self-compatible and easily produce interspecific hybrids with related species. To further improve the ornamental value of hybrids with a larger flower diameter,
Lagerstroemia speciosa and
L. indica were used for interspecific hybridization and hybrid seedlings with traits of the parents were obtained; however, only plant height and plant width had the characteristics of the hybrids and no variation in flower color or flower size was observed [
30]. Recently, research in America, Japan, and China has focused on breeding new cultivars with unique plant architecture, especially dwarf or potted crape myrtle. A few dwarf crape myrtle cultivars with many branches and large flowers were bred through intraspecific hybridization (
L. indica) with the aim of generating dwarf plant architecture [
31,
32,
33,
34,
35].
The molecular study of
Lagerstroemia species has led to the development of a series of genomic SSR markers [
26,
36,
37,
38,
39,
40]. Two single nucleotide polymorphism (SNP) markers (M16337 and M38412) that are highly correlated with internode length and one SNP marker (M25207) that is highly correlated with primary lateral branch height were validated in the F
1 population of
L. indica [
41]. In addition, an SSR marker linked to the dwarf gene, with a genetic distance of 23.33 cM, was screened from the
L. fauriei ×
L. indica “Pocomoke” F
1 population [
35]. Currently, less is known about the plant architecture of the crape myrtle, especially the creeping trait. To analyze the linkage relation between SSR molecular markers and creeping trait, the segregation of the creeping trait was analyzed by the BSA method using the BC
1 population with 174 hybrids and 322 SSR primers. Our results provide an important technical and theoretical basis for plant architecture molecular marker-assisted selective breeding for the
Lagerstroemia species.
4. Discussion
Crape myrtle is popular and widely used in gardens because of its long flowering time and rich and colorful flowers. It can be planted alone or in clusters, used for garden flower belts, or cut flowers can be displayed in vases after pruning and pinning. The lack of varieties with diverse plant architecture types, flowers with a pleasant fragrance, and early-flowering features limits the further application of crape myrtle. Plant architecture traits (e.g., dwarf, weeping, creeping, columnar, and branching angle) have attracted much attention; these have potentially important application value for the plant architecture improvement of horticultural crops. Dwarf crape myrtles are becoming increasingly popular for use as potted plants and indoor flowers [
47]. However, dwarfing and the arborization of trees are complex quantitative traits, which have greater non-additive effects on genetic performance and heritability. The columnar trait is controlled by a single gene (
Co); however, other modification genes that may also play a role have been confirmed in apple [
11]. Incomplete dominant columnar traits (pillar) also exist in peach, with a small branching angle and vertical growth of branches, but no obvious genetic segregation rule was observed for dwarf and compact traits [
7].
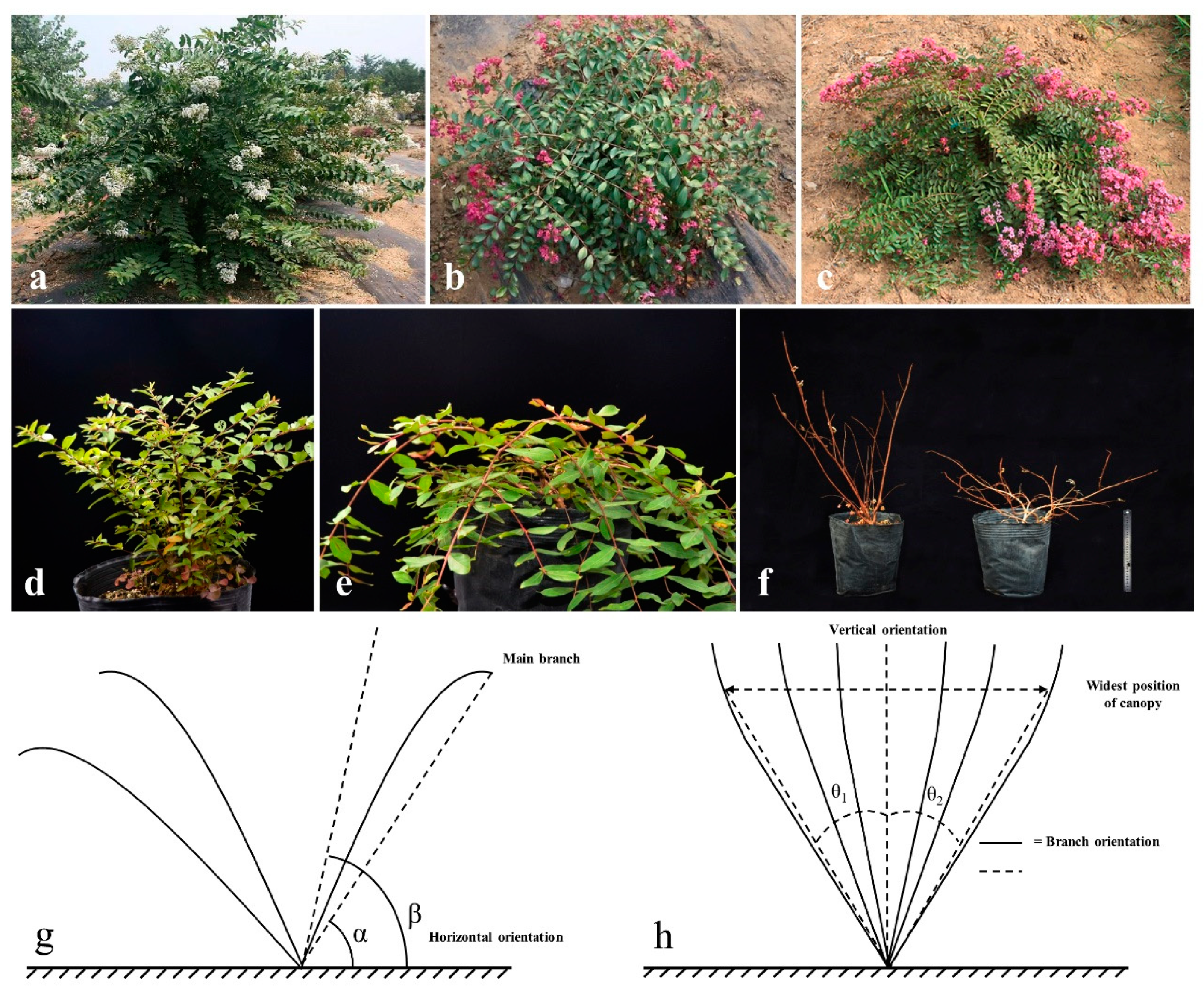
In this study, the number of upright and creeping individuals for the F
1 of
L. fauriei and
L. indica “Creole” was 140 and 52, respectively, and the ratio of the two plant types was 2.69/1. In the BC
1 population of S82 ×
L. fauriei, the ratio of upright (138) to creeping (36) individuals was 3.83/1. However, in the BC
1 population of S82 ×
L. indica “Creole”, the number of upright and creeping individuals was 42 and 150, respectively, with a ratio of 1/3.57 (
Table S4). In the F
1 and S82 ×
L. fauriei populations, the number of upright individuals was much larger than that of creeping individuals because all hybrids were derived from crosses between upright and creeping parents. Although both parents (S82 ×
L. indica “Creole”) were creeping types, upright individuals still appeared in their hybrids (
Table S4). The comprehensive analysis of the segregation of two plant architecture types in three genetic populations cannot determine the number of genes controlling the creeping trait. However, the analysis shows that the upright type has a dominant effect on the creeping type. The upright/creeping plant type of crape myrtle is speculated to be controlled by a major gene and also regulated by minor genes. Our recent research results show that the dwarf trait of crape myrtle is also likely controlled by a major gene and modified by minor genes based on the phenotypic data in the F
1 population of
L. fauriei (♀) ×
L. indica “Pocomoke” (♂) [
35]. Correlation analysis between phenotypic traits is helpful to advance the selection of plant architecture in crape myrtle seedlings, which can shorten the breeding time and enhance breeding efficiency. A significant positive correlation was observed between PH and PW. PCA is verified by individual traits in the BC
1 population, which is similar to the correlation obtained for the F
1 population generated from
L. caudate (♀) and
L. indica “Xiang Xueyun” (♂) [
36]. Eleven quantitative characters for 192 individuals in the F
1 population from a cross between
L. fauriei (♀) and
L. indica “Creole” (♂) were measured and analyzed; the results showed that the diversity index for plant architecture was 1.05 times greater than 1, which means that there is great potential for genetic improvement of the plant architecture [
48]. In addition, most quantitative characters were significantly correlated with plant architecture and branching pattern. The results of heredity for some traits in the F
1 population of
L. speciosa and
L. indica showed that the coefficient of variation in the F
1 generation was 14.58–40.16%, which indicates significant variation [
49].
Traditional breeding methods have the disadvantages of a long cycle, heavy workload, and low effectiveness for improving tree architecture. By using molecular marker-assisted selection, healthy individual plants with target traits can be selected at the seedling stage, which reduces the waste of resources, reduces the cost of breeding, and speeds up the breeding process [
15]. BSA overcomes the constraints of near-isogenic lines and saves time and effort; it is widely used in marker development and gene mapping [
16]. Because BSA does not require a large population, it is very popular in the study of the plant architecture traits of woody plants, such as apple (vertical traits) and peach (columnar and vertical traits) and in the development of genes and markers for peach weeping traits [
2,
10,
50]. In ground-cover chrysanthemum, a random amplified polymorphic DNA (RAPD) marker A-10555 linked to the creeping trait was 7.96 cM from the loci controlling creeping/standard traits [
51]. In crape myrtle, an SSR marker was identified and linked to the dwarf gene with a distance of 23.33 cM between the loci and dwarf gene [
35]. In this study, two SSR markers (S364 and LYS12) closely linked to creeping traits in crape myrtle were obtained according to the principle of BSA. One marker was linked to PCA, with a genetic distance of 23.49 cM, and the other was linked to BA, with a genetic distance of 25.86 cM. The two markers were verified in the BC
1 population, parents, species, and varieties with high accuracy, which suggests that it is feasible to use these two markers to perform molecular marker-assisted breeding of a creeping plant type for crape myrtle. Previous research indicates that the polymorphic loci assayed within 15 cm of the target locus can be identified; loci are detected with decreasing frequency as genetic distance increases [
16]. Therefore, the difference between two near-isogenic pools cannot be determined with the genetic distance of 23.49 cM and 25.86 cM from creeping genes. In addition, BSA has limitations in locating quantitative traits, which can only detect large-effect QTLs and require samples with large phenotypic differences when near-isogenic pools are constructed. In the future, we can use multi-generational hybridization and backcrossing to obtain populations with more significant phenotypic segregation to construct pools and screen markers. Additionally, the number of species-specific molecular markers is a critical factor that influenced the results of this experiment. To achieve an ideal effect in the marker-assisted selection of the creeping trait, we will develop more species-specific molecular markers to enhance the coverage area of the crape myrtle genome.

 ,
, 




{kind=link}
{kind=link}
{kind=link}