Phosphorus Mobilizing Enzymes of Alnus-Associated Ectomycorrhizal Fungi in an Alaskan Boreal Floodplain

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Root Tip Selection and Sampling
2.3. Enzyme Assays
2.4. Molecular Analyses
2.5. Statistical Analyses
3. Results
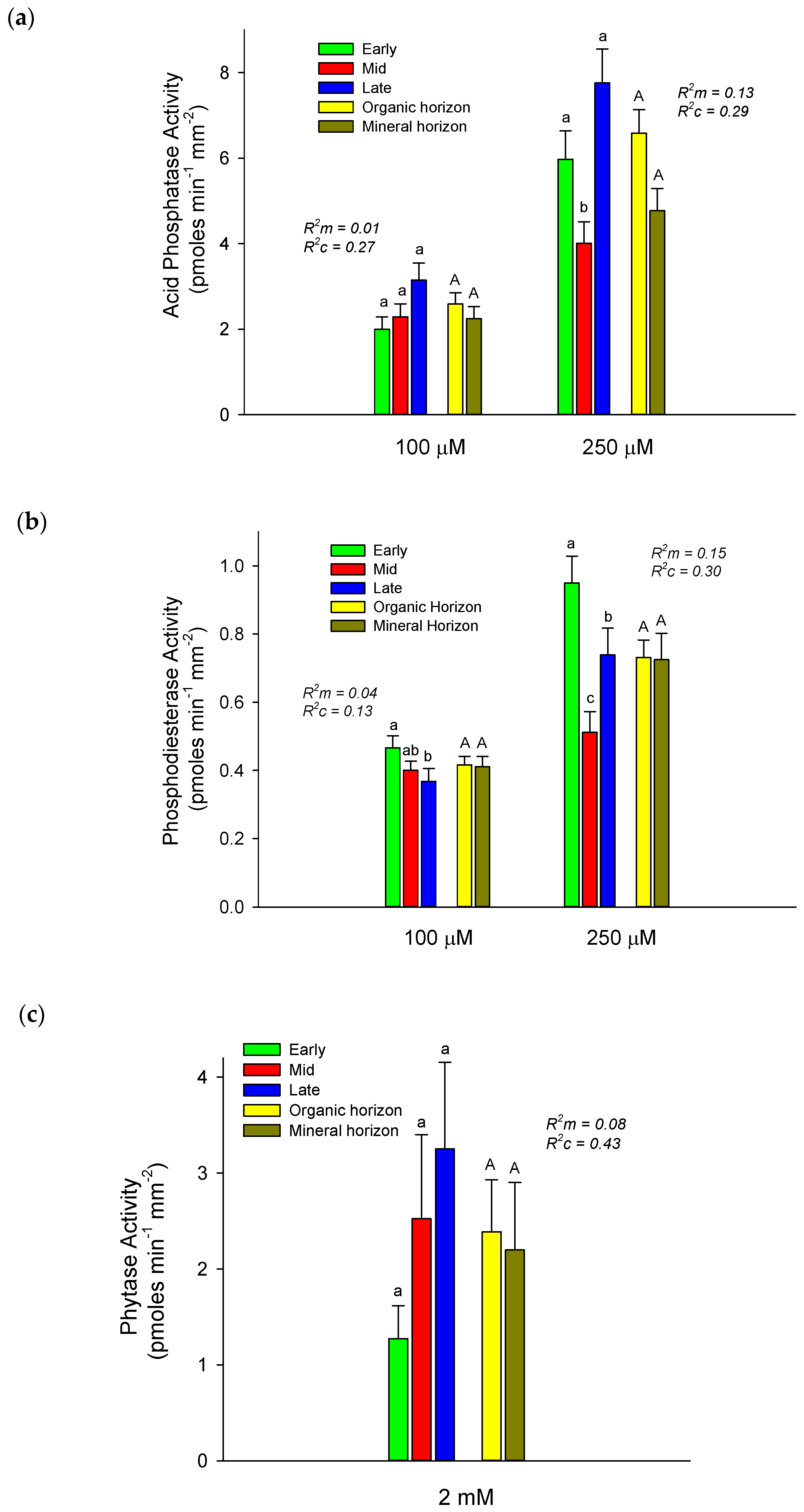
3.1. Alder Root Tip Enzyme Activities
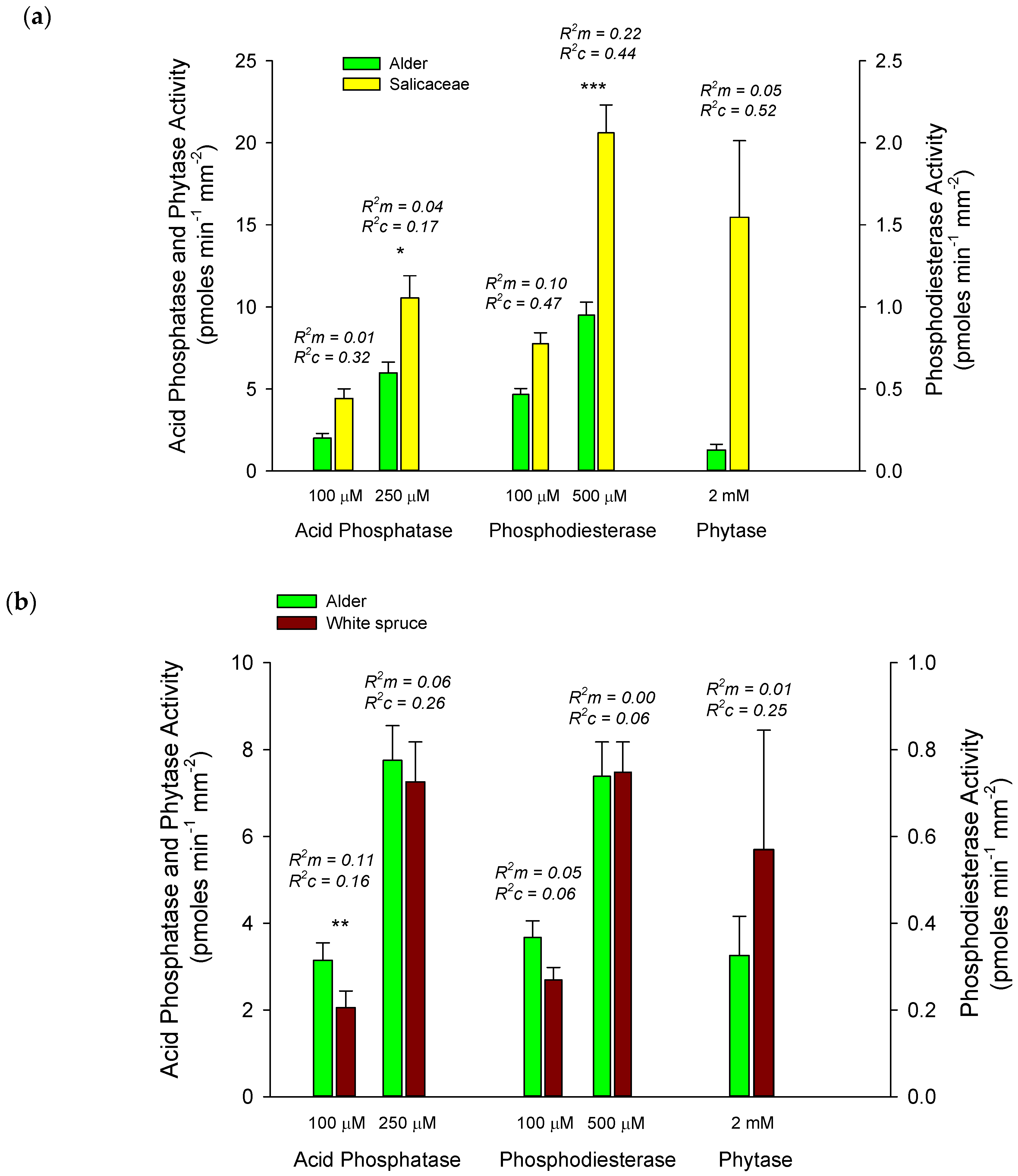
3.2. Comparing Alder Enzyme Activities with Other Host Species
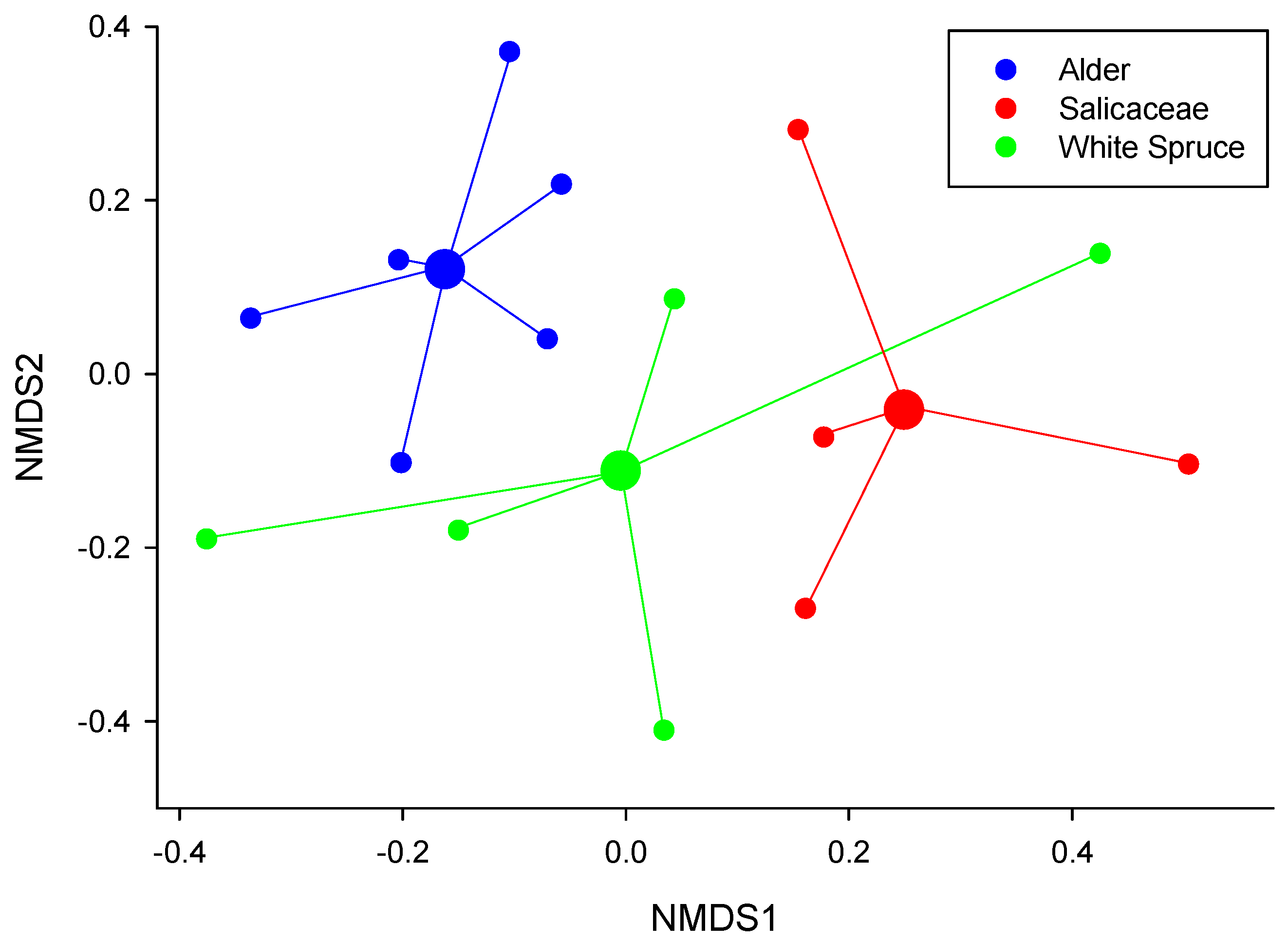
3.3. Fungal Community Identification
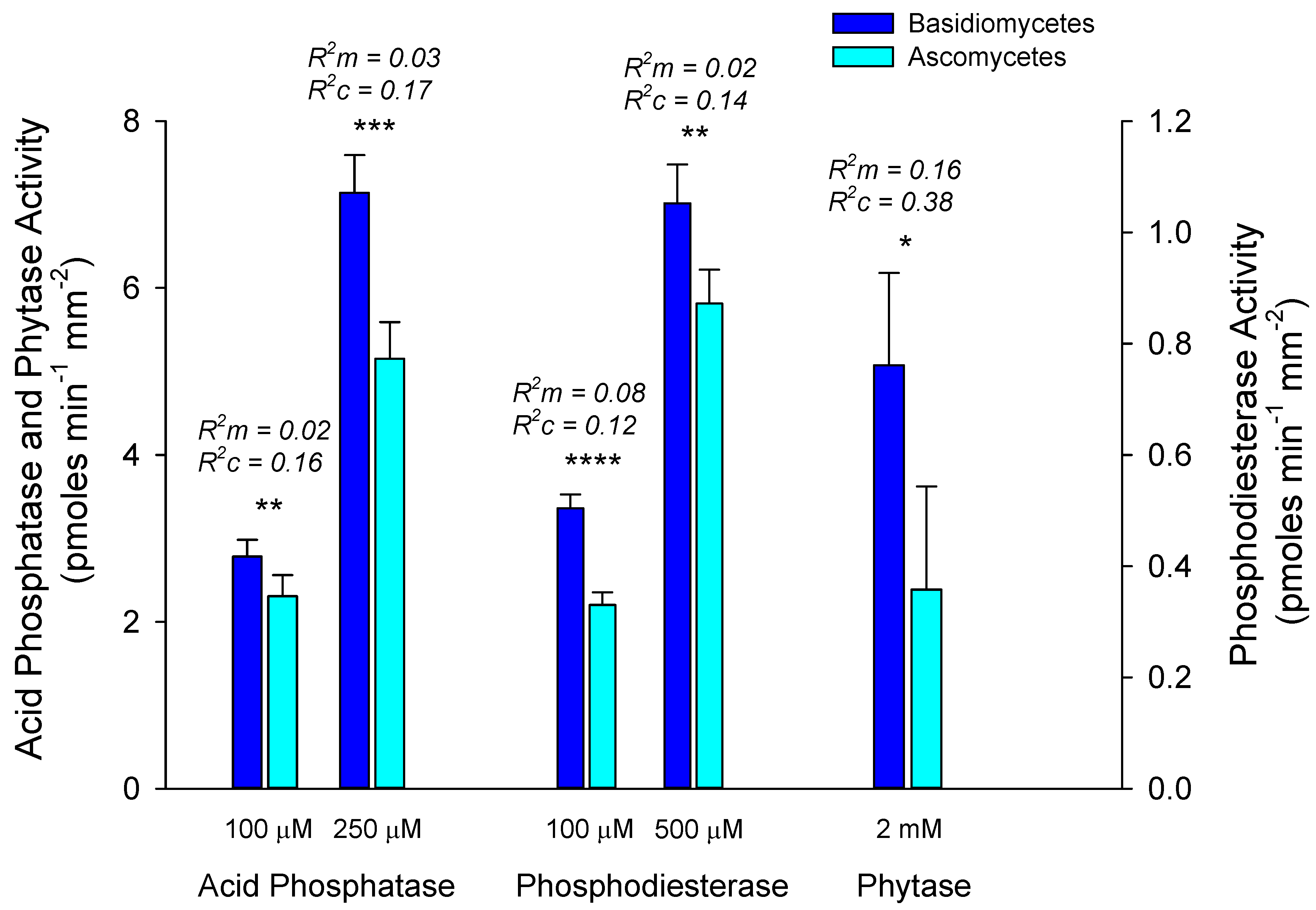
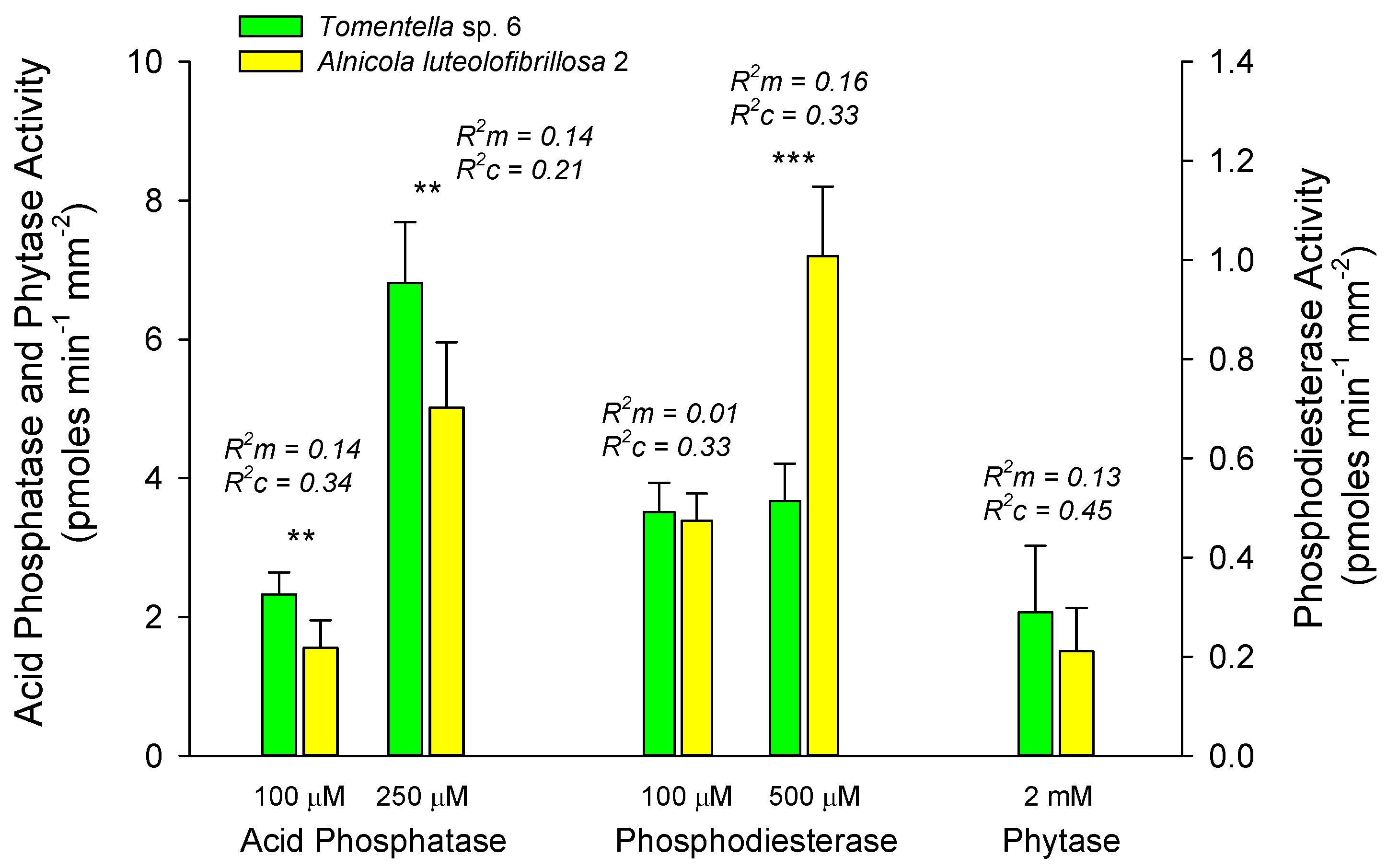
3.4. EMF Specific Activities within and Among Hosts
4. Discussion
4.1. Successional Patterns of Root-Tip P Mobilizing Enzymes
4.2. Differences in Enzyme Activities among Host Species
4.3. Structure of Alnus Ectomycorrhizal Communities
4.4. Patterns of EMF Enzyme Activities
4.5. Phytase Activity
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Enzyme Analyses
Appendix B. Molecular Characterization of Root Tips
Appendix C

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylobin | Assigned Study Name | Best Match | Phylum | GenBank | UNITE ID | Match | Alignment Length (bp) | Score | Alder | Willow | Balsam Poplar | White Spruce | UNKN |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 292 | Alnicola luteolofibrillosa 1 | Alnicola luteolofibrillosa | Basid | JN943976 | 97.9 | 714 | 1234 | 2 | 0 | 0 | 0 | 0 | |
| 297 | Alnicola luteolofibrillosa 2 | Alnicola luteolofibrillosa | Basid | SH190719.06FU | 99 | 599 | 1072 | 26 | 0 | 1 | 1 | 3 | |
| 274 | Alpova alpestris | Alpova alpestris | Basid | SH241746.06FU | 97.24 | 688 | 1162 | 1 | 0 | 0 | 0 | 0 | |
| 167 | Amphinema sp. 1 | Amphinema sp. | Basid | SH229867.06FU | 99.8 | 507 | 931 | 1 | 0 | 0 | 0 | 0 | |
| 68 | Articulospora sp. | uncultured Articulospora | Asco | SH234748.06FU | 96.55 | 464 | 767 | 1 | 0 | 0 | 0 | 0 | |
| 26 | Ascomycete sp. 1 | Ascomycete sp. | Asco | SH037210.06FU | 91.01 | 367 | 488 | 1 | 0 | 0 | 0 | 0 | |
| 27 | Ascomycete sp. 2 | Ascomycete sp. | Asco | SH037210.06FU | 90.44 | 366 | 473 | 0 | 0 | 0 | 1 | 0 | |
| 28 | Ascomycete sp. 3 | Ascomycete sp. | Asco | SH037210.06FU | 93.44 | 673 | 590 | 6 | 0 | 0 | 6 | 4 | |
| 30 | Cadophora finlandica 1 | Cadophora finlandica | Asco | SH207167.06FU | 98.92 | 465 | 830 | 0 | 0 | 0 | 1 | 0 | |
| 31 | Cadophora finlandica 2 | Cadophora finlandica | Asco | SH207167.06FU | 98.07 | 466 | 809 | 0 | 0 | 0 | 1 | 0 | |
| 364 | Cadophora luteo-olivacea | Cadophora luteo-olivacea | Asco | SH204718.06FU | 97.24 | 544 | 918 | 0 | 0 | 0 | 0 | 1 | |
| 376 | Cenococcum geophilum | Cenococcum geophilum | Asco | JN943893 | 97.61 | 964 | 1648 | 0 | 0 | 0 | 0 | 1 | |
| 139 | Chalara holubovae | Chalara holubovae | Asco | SH209190.06FU | 97.78 | 451 | 773 | 0 | 0 | 0 | 0 | 1 | |
| 141 | Chalara microchona | Chalara microchona | Asco | DQ093752 | 98.9 | 451 | 778 | 0 | 0 | 0 | 1 | 0 | |
| 64 | Cladophialophora sp. 1 | uncultured Cladophialophora | Asco | SH025802.06FU | 98.22 | 506 | 885 | 1 | 0 | 0 | 0 | 0 | |
| 103 | Cortinariaceae sp. 1 | Alnicola luteolofibrillosa | Basid | SH190719.06FU | 89.93 | 606 | 767 | 0 | 0 | 0 | 0 | 1 | |
| 104 | Cortinariaceae sp. 2 | Alnicola luteolofibrillosa | Basid | JN943976 | 90.26 | 688 | 878 | 1 | 0 | 0 | 0 | 0 | |
| 238 | Cortinariaceae sp. 3 | Cortinariaceae sp. | Basid | SH232566.06FU | 99.02 | 612 | 1096 | 0 | 0 | 0 | 1 | 0 | |
| 207 | Cortinarius casimiri 1 | Cortinarius casimiri | Basid | SH232912.06FU | 96.08 | 510 | 830 | 0 | 0 | 0 | 1 | 0 | |
| 208 | Cortinarius casimiri 2 | Cortinarius casimiri | Basid | SH232830.06FU | 99.41 | 507 | 920 | 0 | 0 | 0 | 1 | 1 | |
| 237 | Cortinarius cephalixu | Cortinarius cephalixu | Basid | AY174784 | 99.29 | 708 | 1279 | 1 | 0 | 0 | 0 | 0 | |
| 199 | Cortinarius sp. 1 | Cortinarius subsertipes | Basid | HQ604709 | 96.75 | 615 | 1020 | 0 | 0 | 2 | 0 | 3 | |
| 183 | Cyathicula microspora | Cyathicula microspora | Asco | SH014066.06FU | 99.36 | 469 | 850 | 0 | 0 | 0 | 0 | 1 | |
| 34 | Geomyces sp. 1 | Geomyces destructans | Asco | EU854569 | 96.73 | 888 | 1476 | 0 | 1 | 0 | 0 | 1 | |
| 129 | Gloeotinia temulenta | Gloeotinia temulenta | Asco | SH235673.06FU | 99.8 | 506 | 928 | 2 | 0 | 0 | 2 | 2 | |
| 349 | Gyoerffyella sp. 1 | Gyoerffyella sp. | Asco | SH234755.06FU | 97.35 | 452 | 765 | 1 | 0 | 0 | 0 | 2 | |
| 1 | Helotiaceae sp. 1 | Leohumicola minima | Asco | AY706329 | 91.95 | 559 | 765 | 0 | 0 | 0 | 2 | 0 | |
| 92 | Helotiaceae sp. 2 | Helotiaceae sp. | Asco | HQ157915 | 93.11 | 537 | 784 | 0 | 0 | 0 | 0 | 1 | |
| 180 | Helotiaceae sp. 3 | uncultured Helotiaceae | Asco | SH030055.06FU | 97.38 | 458 | 774 | 0 | 0 | 0 | 1 | 0 | |
| 377 | Helotiaceae sp. 4 | Helotiales sp. | Asco | SH444216.06FU | 87.31 | 536 | 592 | 2 | 0 | 0 | 0 | 0 | |
| 329 | Helotiales sp. 1 | Leptodontidium sp. | Asco | SH034136.06FU | 99.06 | 531 | 953 | 0 | 0 | 0 | 0 | 1 | |
| 347 | Hyaloscyphaceae sp. 1 | Calycina languida | Asco | SH208263.06FU | 90.74 | 475 | 638 | 0 | 0 | 0 | 1 | 0 | |
| 175 | Hygrophorus pustulatus | Hygrophorus pustulatus | Basid | FJ845412 | 99.8 | 501 | 920 | 0 | 0 | 0 | 1 | 0 | |
| 282 | Inocybaceae sp. 1 | Inocybaceae sp. | Basid | SH240210.06FU | 99.31 | 583 | 1051 | 1 | 0 | 1 | 0 | 0 | |
| 6 | Lachnum sp. 1 | uncultured Lachnum | Asco | SH189775.06FU | 97.44 | 468 | 795 | 3 | 0 | 0 | 3 | 1 | |
| 89 | Lactarius lilacinus | Lactarius lilacinus | Basid | SH238120.06FU | 99.21 | 630 | 1136 | 2 | 0 | 0 | 0 | 0 | |
| 284 | Leohumicola minima | Leohumicola minima | Asco | AY706329 | 97.27 | 549 | 928 | 1 | 0 | 0 | 0 | 0 | |
| 357 | Leptodontidium orchidicola 1 | Leptodontidium orchidicola | Asco | AF486133 | 98.93 | 652 | 1168 | 0 | 0 | 0 | 0 | 5 | |
| 359 | Leptodontidium orchidicola 2 | Leptodontidium orchidicola | Asco | AF486133 | 97.85 | 650 | 1118 | 0 | 0 | 0 | 0 | 1 | |
| 360 | Leptodontidium orchidicola 3 | Leptodontidium orchidicola | Asco | AF486133 | 98.29 | 644 | 1125 | 1 | 0 | 0 | 0 | 0 | |
| 361 | Leptodontidium orchidicola 4 | Leptodontidium orchidicola | Asco | AF486133 | 98.59 | 638 | 1127 | 0 | 0 | 0 | 0 | 1 | |
| 363 | Leptodontidium orchidicola 5 | Leptodontidium orchidicola | Asco | AF486133 | 98.9 | 637 | 1138 | 0 | 0 | 0 | 0 | 1 | |
| 14 | Lycoperdon foetidum | Lycoperdon foetidum | Basid | SH244736.06FU | 97.42 | 658 | 1120 | 1 | 0 | 0 | 0 | 0 | |
| 76 | Meliniomyces bicolor | Meliniomyces bicolor | Asco | SH207170.06FU | 99.79 | 475 | 872 | 1 | 0 | 0 | 0 | 0 | |
| 46 | Phialocephala fortinii 1 | Phialocephala fortinii | Asco | SH213468.06FU | 98.32 | 476 | 833 | 0 | 0 | 0 | 1 | 0 | |
| 47 | Phialocephala fortinii 2 | Phialocephala fortinii | Asco | SH213468.06FU | 98.82 | 592 | 1055 | 2 | 0 | 0 | 0 | 0 | |
| 49 | Phialocephala fortinii 3 | Phialocephala fortinii | Asco | SH213468.06FU | 98.95 | 476 | 850 | 1 | 0 | 0 | 0 | 0 | |
| 339 | Phialocephala fortinii 4 | Phialocephala fortinii | Asco | SH213468.06FU | 97.69 | 477 | 824 | 5 | 0 | 0 | 2 | 1 | |
| 366 | Phialocephala lagerbergii | Phialocephala lagerbergii | Asco | SH204727.06FU | 99.2 | 500 | 900 | 0 | 0 | 0 | 0 | 1 | |
| 320 | Piloderma sp. 1 | Piloderma sp. | Basid | UDB001733 | 100 | 621 | 1147 | 1 | 0 | 0 | 1 | 0 | |
| 128 | Plectosphaerellaceae sp. | Verticillium leptobactrum | Asco | SH235676.06FU | 94.11 | 475 | 715 | 0 | 0 | 0 | 0 | 1 | |
| 61 | Russula cessans | Russula cessans | Basid | FJ845437 | 98.92 | 646 | 1151 | 0 | 0 | 0 | 2 | 0 | |
| 59 | Russula sp. 1 | Russula cessans | Basid | FJ845437 | 93.32 | 704 | 1044 | 0 | 0 | 0 | 1 | 0 | |
| 60 | Russula sp. 2 | Russula cessans | Basid | FJ845437 | 94.64 | 634 | 976 | 1 | 0 | 0 | 0 | 0 | |
| 380 | Russula sp. 3 | Russula cessans | Basid | FJ845437 | 95.04 | 605 | 941 | 0 | 0 | 0 | 1 | 0 | |
| 262 | Russula versicolor | Russula versicolor | Basid | SH224391.06FU | 98.69 | 610 | 1081 | 0 | 0 | 0 | 0 | 2 | |
| 343 | Sebacina dimitica | Sebacina dimitica | Basid | SH231629.06FU | 98.49 | 530 | 931 | 0 | 0 | 0 | 0 | 1 | |
| 85 | Sebacina epigaea | Sebacina epigaea | Basid | EU819519 | 95.69 | 532 | 907 | 0 | 6 | 0 | 1 | 0 | |
| 119 | Sebacina sp. 2 | Sebacina sp. | Basid | SH305078.06FU | 98.3 | 529 | 926 | 0 | 3 | 8 | 0 | 1 | |
| 159 | Sebacina sp. 3 | Sebacina sp. | Basid | SH231609.06FU | 99.25 | 534 | 963 | 1 | 0 | 0 | 0 | 0 | |
| 161 | Sebacina sp. 4 | Sebacina sp. | Basid | SH231664.06FU | 98.71 | 541 | 957 | 3 | 0 | 0 | 0 | 0 | |
| 344 | Sebacina sp. 5 | Sebacina sp. | Basid | SH231595.06FU | 99.26 | 541 | 977 | 0 | 0 | 1 | 0 | 3 | |
| 24 | Tetracladium maxilliforme | Tetracladium maxilliforme | Asco | EU883429 | 97.13 | 593 | 1000 | 2 | 0 | 0 | 0 | 0 | |
| 100 | Thelephora terrestris | Thelephora terrestris | Basid | HM189958 | 93.22 | 708 | 1031 | 3 | 0 | 0 | 0 | 0 | |
| 20 | Thelephoraceae sp. 1 | Thelephoraceae sp. | Basid | SH195974.06FU | 94.75 | 590 | 917 | 0 | 0 | 0 | 0 | 1 | |
| 21 | Thelephoraceae sp. 2 | Thelephoraceae sp. | Basid | SH195974.06FU | 93 | 586 | 848 | 1 | 0 | 0 | 0 | 0 | |
| 99 | Thelephoraceae sp. 3 | Thelephoraceae sp. | Basid | SH219843.06FU | 95.52 | 536 | 856 | 1 | 0 | 0 | 0 | 0 | |
| 132 | Tomentella ferruginea 1 | Tomentella ferruginea | Basid | AF272909 | 98.74 | 555 | 985 | 0 | 1 | 0 | 1 | 0 | |
| 134 | Tomentella ferruginea 2 | Tomentella ferruginea | Basid | AF272909 | 98.17 | 546 | 952 | 0 | 0 | 5 | 0 | 2 | |
| 7 | Tomentella sp. 1 | Tomentella sp. | Basid | SH213382.06FU | 99.65 | 576 | 1055 | 2 | 0 | 0 | 0 | 0 | |
| 239 | Tomentella sp. 10 | Tomentella ellisii | Basid | SH222911.06FU | 96.61 | 590 | 977 | 1 | 0 | 0 | 0 | 0 | |
| 264 | Tomentella sp. 12 | uncultured Tomentella | Basid | SH202475.06FU | 97.74 | 576 | 992 | 0 | 0 | 0 | 0 | 1 | |
| 269 | Tomentella sp. 13 | Tomentella sp. | Basid | U83482 | 96.05 | 683 | 1110 | 1 | 0 | 0 | 0 | 0 | |
| 379 | Tomentella sp. 14 | Tomentella sp. | Basid | SH202538.06FU | 99.15 | 587 | 1055 | 0 | 1 | 0 | 0 | 0 | |
| 8 | Tomentella sp. 2 | uncultured Tomentella | Basid | SH199023.06FU | 98.78 | 572 | 1016 | 0 | 0 | 2 | 3 | 2 | |
| 22 | Tomentella sp. 6 | Tomentella sublilacina | Basid | HM189994 | 95.83 | 695 | 1120 | 18 | 0 | 0 | 1 | 0 | |
| 98 | Tomentella sp. 7 | uncultured Tomentella | Basid | SH021829.06FU | 94.67 | 582 | 896 | 0 | 0 | 0 | 0 | 1 | |
| 223 | Tomentella sp. 9 | uncultured Tomentella | Basid | SH219870.06FU | 98.14 | 590 | 1027 | 0 | 0 | 0 | 0 | 2 | |
| 224 | Tomentella viridula | Tomentella viridula | Basid | AF272914 | 99.66 | 581 | 1061 | 0 | 0 | 0 | 0 | 1 | |
| 280 | Tomentellopsis sp. 1 | Tomentellopsis submollis | Basid | SH199523.06FU | 96.98 | 597 | 1009 | 1 | 0 | 0 | 0 | 0 | |
| 192 | Trichophaea sp. 1 | Trichophaea cf hybrida | Asco | SH227980.06FU | 100 | 522 | 965 | 1 | 0 | 0 | 0 | 0 | |
| 10 | Tuber sp. 1 | uncultured Tuber | Asco | SH204354.06FU | 99.66 | 592 | 1081 | 0 | 3 | 0 | 5 | 0 | |
| 62 | Tuber sp. 2 | Tuber rapaeodorum | Asco | EU784430 | 95.88 | 582 | 937 | 1 | 0 | 0 | 0 | 0 | |
| 384 | Tuber sp. 3 | uncultured Tuber | Asco | SH204354.06FU | 99.66 | 592 | 1081 | 0 | 0 | 0 | 0 | 1 | |
| 41 | Vibrisseaceae sp. 1 | Phialocephala fortinii | Asco | SH016932.06FU | 84.11 | 151 | 147 | 0 | 0 | 0 | 0 | 1 | |
| 150 | Wilcoxina rehmii 2 | Wilcoxina rehmii | Asco | SH023929.06FU | 97.58 | 455 | 778 | 0 | 0 | 0 | 1 | 0 |
References
- Marion, G.M.; Van Cleve, K.; Dyrness, C.T.; Black, C.H. The soil chemical environment along a primary successional sequence on the Tanana River floodplain, interior Alaska. Can. J. For. Res. 1993, 23, 923–927. [Google Scholar] [CrossRef]
- Giesler, R.; Petersson, T.; Hogberg, P. Phosphorus limitation in boreal forests: Effects of aluminum and iron accumulation in the humus layer. Ecosystems 2002, 5, 300–314. [Google Scholar] [CrossRef]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Walker, T.W.; Syers, J.K. The fate of phosphorus during pedogenesis. Geoderma 1976, 1, 1–19. [Google Scholar] [CrossRef]
- Crews, T.E.; Kitayama, K.; Fownes, J.H.; Riley, R.H.; Herbert, D.A.; Mueller-Dombois, D.; Vitousek, P.M. Changes in soil phosphorus fractions and ecosystem dynamics across a long chronosequence in Hawaii. Ecology 1995, 76, 1407–1424. [Google Scholar] [CrossRef]
- Allison, V.J.; Condron, L.M.; Peltzer, D.A.; Richardson, S.J.; Turner, B.L. Changes in enzyme activities and soil microbial community composition along carbon and nutrient gradients at the Franz Josef chronosequence, New Zealand. Soil Biol. Biochem. 2007, 39, 1770–1781. [Google Scholar] [CrossRef]
- Harrison, A.F. Labile organic phosphorus mineralization in relationship to soil properties. Soil Biol. Biochem. 1982, 14, 343–351. [Google Scholar] [CrossRef]
- Turner, B.L.; Condron, L.M.; Richardson, S.J.; Peltzer, D.A.; Allison, V.J. Soil organic phosphorus transformations during pedogenesis. Ecosystems 2007, 10, 1166–1181. [Google Scholar] [CrossRef]
- Ruess, R.W.; Anderson, M.D.; McFarland, J.M.; Kielland, K.; Olson, K.; Taylor, D.E. Ecosystem-level consequences of symbiont partnerships in a N2-fixing shrub from interior Alaskan floodplains. Ecol. Monogr. 2013, 83, 177–194. [Google Scholar] [CrossRef]
- Houlton, B.Z.; Wang, Y.P.; Vitousek, P.M.; Field, C.B. A unifying framework for dinitrogen fixation in the terrestrial biosphere. Nature 2008, 454, 327–334. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Ruess, R.W.; McFarland, J.M.; Trummer, L.M.; Rohrs-Richey, J.K. Disease-mediated declines in N-fixation inputs by Alnus tenuifolia to early-successional floodplains in interior and south-central Alaska. Ecosystems 2009, 12, 489–501. [Google Scholar] [CrossRef]
- Uliassi, D.D.; Ruess, R.W. Limitations to symbiotic nitrogen fixation in primary succession on the Tanana River floodplain, Alaska. Ecology 2002, 83, 88–103. [Google Scholar] [CrossRef]
- Anderson, M.D.; Ruess, R.W.; Uliassi, D.D.; Mitchell, J.S. Estimating N2 fixation in two species of Alnus in interior Alaska using acetylene reduction and 15N2 uptake. Ecoscience 2004, 11, 102–112. [Google Scholar] [CrossRef]
- Giardina, C.P.; Huffman, S.; Binkley, D.; Caldwell, B.A. Alders increase soil phosphorus availability in a Douglas-fir plantation. Can. J. For.Res. 1995, 25, 1652–1657. [Google Scholar] [CrossRef]
- Walker, J.K.M.; Cohen, H.; Higgins, L.M.; Kennedy, P.G. Testing the link between community structure and function for ectomycorrhizal fungi involved in a global tripartite symbiosis. New Phytol. 2014, 202, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Pritsch, K.; Munch, J.C.; Buscot, F. Morphological and anatomical characterisation of black alder Alnus glutinosa (L.) Gaertn. ectomycorrhizas. Mycorrhiza 1997, 7, 201–216. [Google Scholar] [CrossRef]
- Plassard, C.; Louche, J.; Ali, M.A.; Duchemin, M.; Legname, E.; Cloutier-Hurteau, B. Diversity in phosphorus mobilisation and uptake in ectomycorrhizal fungi. Ann. For. Sci. 2011, 68, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Courty, P.E.; Pritsch, K.; Schloter, M.; Hartmann, A.; Garbaye, J. Activity profiling of ectomycorrhiza communities in two forest soils using multiple enzymatic tests. New Phytol. 2005, 167, 309–319. [Google Scholar] [CrossRef]
- Koide, R.T.; Courty, P.E.; Garbaye, J. Research perspectives on functional diversity in ectomycorrhizal fungi. New Phytol. 2007, 174, 240–243. [Google Scholar] [CrossRef]
- Tedersoo, L.; Suvi, T.; Jairus, T.; Kõljalg, U. Forest microsite effects on community composition of ectomycorrhizal fungi on seedlings of Picea abies and Betula pendula. Environ. Microbiol. 2008, 10, 1189–1201. [Google Scholar] [CrossRef]
- Tedersoo, L.; Suvi, T.; Jairus, T.; Ostonen, I.; Polme, S. Revisiting ectomycorrhizal fungi of the genus Alnus: Differential host specificity, diversity and determinants of the fungal community. New Phytol. 2009, 182, 727–735. [Google Scholar] [CrossRef]
- Kennedy, P.G.; Hill, L.T. A molecular and phylogenetic analysis of the structure and specificity of Alnus rubra ectomycorrhizal assemblages. Fungal Ecol. 2010, 3, 195–204. [Google Scholar] [CrossRef]
- Põlme, S.; Bahram, M.; Yamanaka, T.; Nara, K.; Dai, Y.C.; Grebenc, T.; Kraigher, H.; Toivonen, M.; Wang, P.H.; Matsuda, Y.; et al. Biogeography of ectomycorrhizal fungi associated with alders (Alnus spp.) in relation to biotic and abiotic variables at the global scale. New Phytol. 2013, 198, 1239–1249. [Google Scholar] [CrossRef]
- Roy, M.; Rochet, J.; Manzi, S.; Jargeat, P.; Gryta, H.; Moreau, P.A.; Gardes, M. What determines Alnus-associated ectomycorrhizal community diversity and specificity? A comparison of host and habitat effects at a regional scale. New Phytol. 2013, 198, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Kroehler, C.J.; Linkins, A.E. The root surface phosphatases of Eriophorum vaginatum: Effects of temperature, pH, substrate concentration and inorganic phosphorus. Plant Soil 1988, 105, 3–10. [Google Scholar] [CrossRef]
- Colpaert, J.V.; Van Laere, A.; Van Tichelen, K.K.; Van Assche, J.A. The use of inositol hexaphosphate as a phosphorus source by mycorrhizal and non-mycorrhizal Scots Pine (Pinus sylvestris). Funct. Ecol. 1997, 11, 407–415. [Google Scholar] [CrossRef]
- Turner, B.L. Resource partitioning for soil phosphorus: a hypothesis. J. Ecol. 2008, 96, 698–702. [Google Scholar] [CrossRef]
- Louche, J.; Ali, M.A.; Cloutier-Hurteau, B.; Sauvage, F.X.; Quiquampoix, H.; Plassard, C. Efficiency of acid phosphatases secreted from the ectomycorrhizal fungus Hebeloma cylindrosporum to hydrolyse organic phosphorus in podzols. FEMS Microbiol. Ecol. 2010, 73, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, T.N.; Lloyd, A.H.; Nossov, D.; Ruess, R.W.; Charlton, B.A.; Kielland, K. Twenty-five years of vegetation change along a putative successional chronosequence on the Tanana River, Alaska. Can. J. For. Res. 2010, 40, 1273–1287. [Google Scholar] [CrossRef]
- Viereck, L.A.; Vancleve, K.; Adams, P.C.; Schlentner, R.E. Climate of the Tanana River floodplain near Fairbanks, Alaska. Can. J. For. Res. 1993, 23, 899–913. [Google Scholar] [CrossRef]
- Anderson, M.D.; Ruess, R.W.; Taylor, D.L.; Myrold, D.D. Host species and habitat affect nodulation by specific Frankia genotypes in two species of Alnus in interior Alaska. Oecologia 2009, 160, 619–630. [Google Scholar] [CrossRef]
- Anderson, M.D.; Taylor, D.E.; Ruess, R.W. Phylogeny and assemblage composition of Frankia in Alnus tenuifolia nodules across a primary successional sere in interior Alaska. Mol. Ecol. 2013, 22, 3864–3877. [Google Scholar] [CrossRef] [PubMed]
- Pritsch, K.; Raidl, S.; Marksteiner, E.; Blaschke, H.; Agerer, R.; Schloter, M.; Hartmann, A. A rapid and highly sensitive method for measuring enzyme activities in single mycorrhizal tips using 4-methylumbelliferone-labelled fluorogenic substrates in a microplate system. J. Microbiol. Methods 2004, 58, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Giles, C.D.; Cade-Menun, B.J.; Hill, J.E. The inositol phosphates in soils and manures: Abundance, cycling, and measurement. Can. J. Soil Sci. 2011, 91, 397–416. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes - application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Taylor, D.L.; Houston, S. A bioinformatics pipeline for sequence-based analyses of fungal biodiversity. In Methods in Molecular Biology; Xu, J.R., Bluhm, B.H., Eds.; Humana Press: Totowa, NJ, USA, 2011; Volume 722, pp. 141–155. [Google Scholar]
- Timling, I.; Walker, D.A.; Nusbaum, C.; Lennon, N.J.; Taylor, D.L. Rich and cold: Diversity, distribution and drivers of fungal communities in patterned-ground ecosystems of the North American Arctic. Mol. Ecol. 2014, 23, 3258–3272. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Flanagan, P.W.; Van Cleve, K. Nutrient cycling in relation to decomposition and organic matter quality in taiga ecosystems. Can. J. For. Res. 1983, 13, 795–817. [Google Scholar] [CrossRef]
- Ruess, R.W.; Hendrick, R.L.; Bryant, J.P. Regulation of fine root dynamics by mammalian browsers in early successional Alaskan taiga forests. Ecology 1998, 79, 2706–2720, Erratum in 1999, 80, 1101. [Google Scholar] [CrossRef]
- Gentili, F.; Wall, L.G.; Huss-Danell, K. Effects of phosphorus and nitrogen on nodulation are seen already at the stage of early cortical cell divisions in Alnus incana. Ann. Bot. 2006, 98, 309–315. [Google Scholar] [CrossRef]
- Huss-Danell, K.; Gentili, F.; Valverde, C.; Wall, L.G.; Wiklund, A. Phosphorus is important in nodulation of actinorhizal plants and legumes. In Nitrogen Fixation: Global Perspectives; Finan, T., O’Brian, N., Layzell, D., Vessey, K., Newton, W., Eds.; CAB International: Wallingford, UK, 2002; pp. 163–166. [Google Scholar]
- Mitchell, J.S.; Ruess, R.W. N2-fixing alder (Alnus viridis spp. fruticosa) effects on soil properties across a secondary successional chronosequence in interior Alaska. Biogeochemistry 2009, 95, 215–229. [Google Scholar] [CrossRef]
- Molina, R. Ectomycorrhizal specificity in the genus Alnus. Can. J. Bot. 1981, 59, 325–334. [Google Scholar] [CrossRef]
- Kennedy, P.G.; Garibay-Orijel, R.; Higgins, L.M.; Angeles-Arguiz, R. Ectomycorrhizal fungi in Mexican Alnus forests support the host co-migration hypothesis and continental-scale patterns in phylogeography. Mycorrhiza 2011, 21, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Moreau, P.A.; Peintner, U.; Gardes, M. Phylogeny of the ectomycorrhizal mushroom genus Alnicola (Basidiomycota, Cortinariaceae) based on rDNA sequences with special emphasis on host specificity and morphological characters. Mol. Phylogenet. Evolut. 2006, 38, 794–807. [Google Scholar] [CrossRef] [PubMed]
- Rochet, J.; Moreau, P.A.; Manzi, S.; Gardes, M. Comparative phylogenies and host specialization in the alder ectomycorrhizal fungi Alnicola, Alpova and Lactarius (Basidiomycota) in Europe. BMC Evolut. Biol. 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Bogar, L.M.; Kennedy, P.G. New wrinkles in an old paradigm: Neighborhood effects can modify the structure and specificity of Alnus-associated ectomycorrhizal fungal communities. FEMS Microbiol. Ecol. 2013, 83, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Bolan, N.S. A critical review on the role of mycorrhizal fungi in the uptake of phosphorus by plants. Plant Soil 1992, 134, 189–207. [Google Scholar] [CrossRef]
- Cairney, J.W.G. Intraspecific physiological variation: Implications for understanding functional diversity in ectomycorrhizal fungi. Mycorrhiza 1999, 9, 125–135. [Google Scholar] [CrossRef]
- Cairney, J.W.G. Ectomycorrhizal fungi: The symbiotic route to the root for phosphorus in forest soils. Plant Soil 2011, 344, 51–71. [Google Scholar] [CrossRef]
- Wurzburger, N.; Bellenger, J.P.; Kraepiel, A.M.L.; Hedin, L.O. Molybdenum and phosphorus interact to constrain asymbiotic nitrogen fixation in tropical forests. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Agerer, R. Exploration types of ectomycorrhizae: A proposal to classify ectomycorrhizal mycelial systems according to their patterns of differentiation and putative ecological importance. Mycorrhiza 2001, 11, 107–114. [Google Scholar] [CrossRef]
- Alvarez, M.; Huygens, D.; Díaz, L.M.; Villanueva, C.A.; Heyser, W.; Boeckx, P. The spatial distribution of acid phosphatase activity in ectomycorrhizal tissues depends on soil fertility and morphotype, and relates to host plant phosphorus uptake. Plant Cell Environ. 2012, 35, 126–135. [Google Scholar] [CrossRef]
- Colpaert, J.V.; Van Tichelen, K.K.; Van Assche, J.A.; Van Laere, A. Short-term phosphorus uptake rates in mycorrhizal and non-mycorrhizal roots of intact Pinus sylvestris seedlings. New Phytol. 1999, 143, 589–597. [Google Scholar] [CrossRef]
- Rineau, F.; Garbaye, J. Does forest liming impact the enzymatic profiles of ectomycorrhizal communities through specialized fungal symbionts? Mycorrhiza 2009, 19, 493–500. [Google Scholar] [CrossRef] [PubMed]
- van Aarle, I.M.; Plassard, C. Spatial distribution of phosphatase activity associated with ectomycorrhizal plants is related to soil type. Soil Biol. Biochem. 2010, 42, 324–330. [Google Scholar] [CrossRef]
- Turner, B.L.; Papházy, M.J.; Haygarth, P.M.; McKelvie, I.D. Inositol phosphates in the environment. Philos. Trans. Royal Soc. B Biol. Sci. 2002, 357, 449–469. [Google Scholar] [CrossRef] [Green Version]
- Dalai, R.C. Soil Organic Phosphorus. In Advances in Agronomy; Brady, N.C., Ed.; Academic Press: New York, NY, USA, 1977; Volume 29, pp. 83–117. [Google Scholar]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B.; et al. TIGR gene indices clustering tools (TGICL): A software system for fast clustering of large EST datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef]
- Huang, X.; Madan, A. CAP3: A DNA sequence assembly program. Genome Res. 1999, 9, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruess, R.W.; Swanson, M.M.; Kielland, K.; McFarland, J.W.; Olson, K.D.; Taylor, D.L. Phosphorus Mobilizing Enzymes of Alnus-Associated Ectomycorrhizal Fungi in an Alaskan Boreal Floodplain. Forests 2019, 10, 554. https://doi.org/10.3390/f10070554
Ruess RW, Swanson MM, Kielland K, McFarland JW, Olson KD, Taylor DL. Phosphorus Mobilizing Enzymes of Alnus-Associated Ectomycorrhizal Fungi in an Alaskan Boreal Floodplain. Forests. 2019; 10(7):554. https://doi.org/10.3390/f10070554
Chicago/Turabian StyleRuess, Roger W., Michaela M. Swanson, Knut Kielland, Jack W. McFarland, Karl D. Olson, and D. Lee Taylor. 2019. "Phosphorus Mobilizing Enzymes of Alnus-Associated Ectomycorrhizal Fungi in an Alaskan Boreal Floodplain" Forests 10, no. 7: 554. https://doi.org/10.3390/f10070554
APA StyleRuess, R. W., Swanson, M. M., Kielland, K., McFarland, J. W., Olson, K. D., & Taylor, D. L. (2019). Phosphorus Mobilizing Enzymes of Alnus-Associated Ectomycorrhizal Fungi in an Alaskan Boreal Floodplain. Forests, 10(7), 554. https://doi.org/10.3390/f10070554