Endophytic Mycobiota of Jingbai Pear Trees in North China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction, Amplification of ITS rDNA Region and Illumina Sequencing
2.3. Pre-Processing and Analysis of ITS rDNA Sequences
3. Results
3.1. Information on MiSeq Sequencing Data
3.2. Fungal Richness, Diversity and Evenness
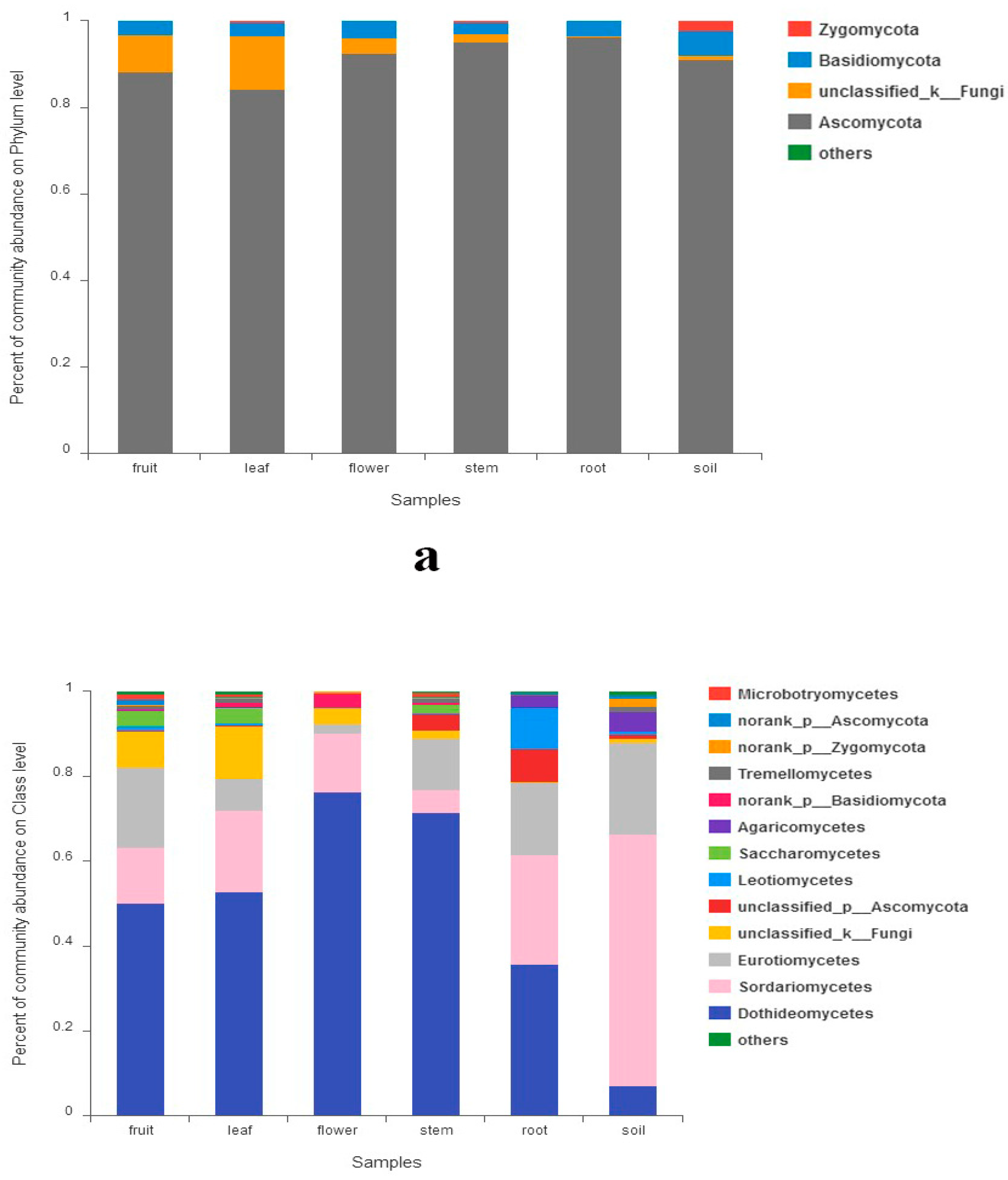
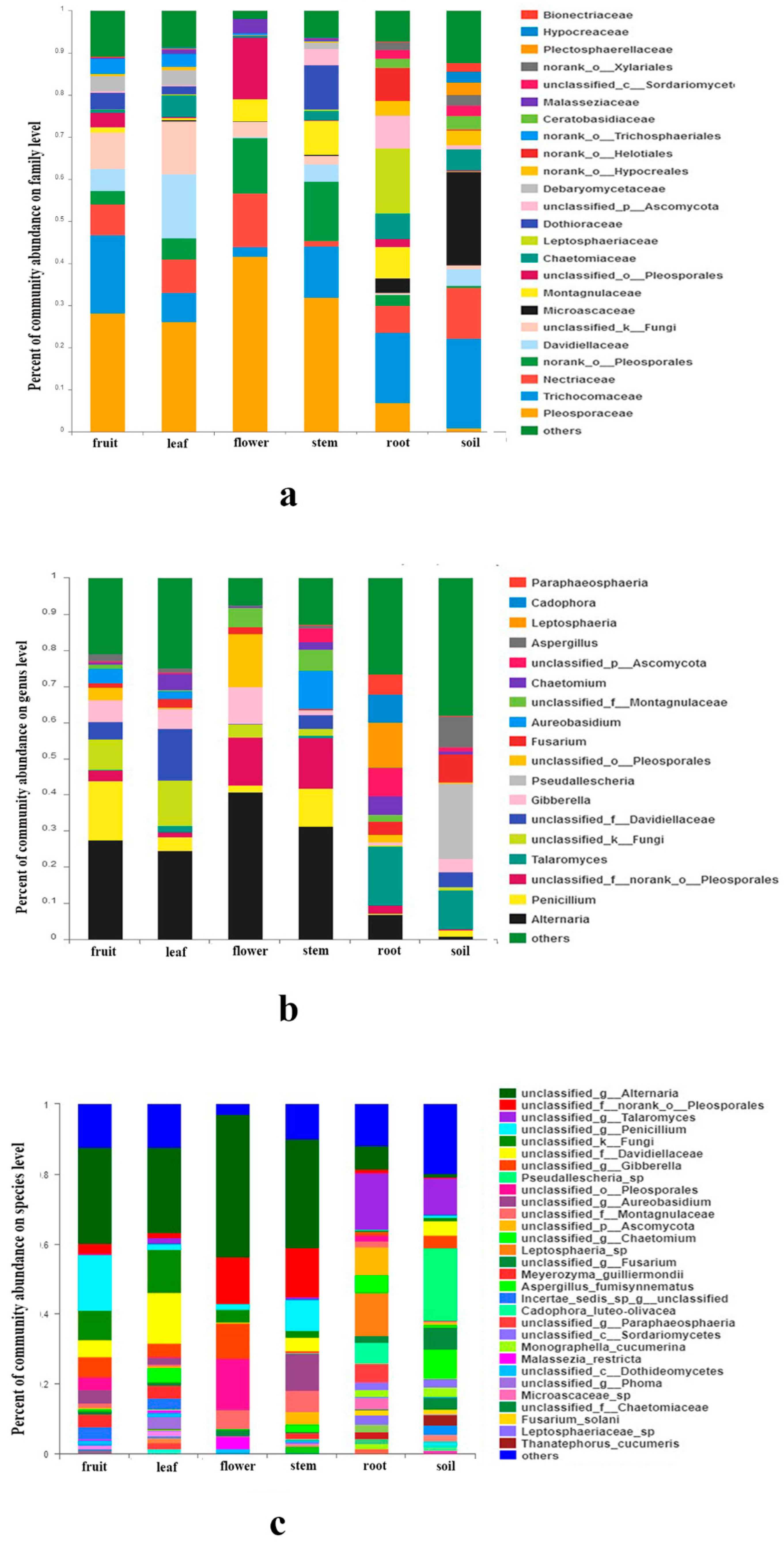
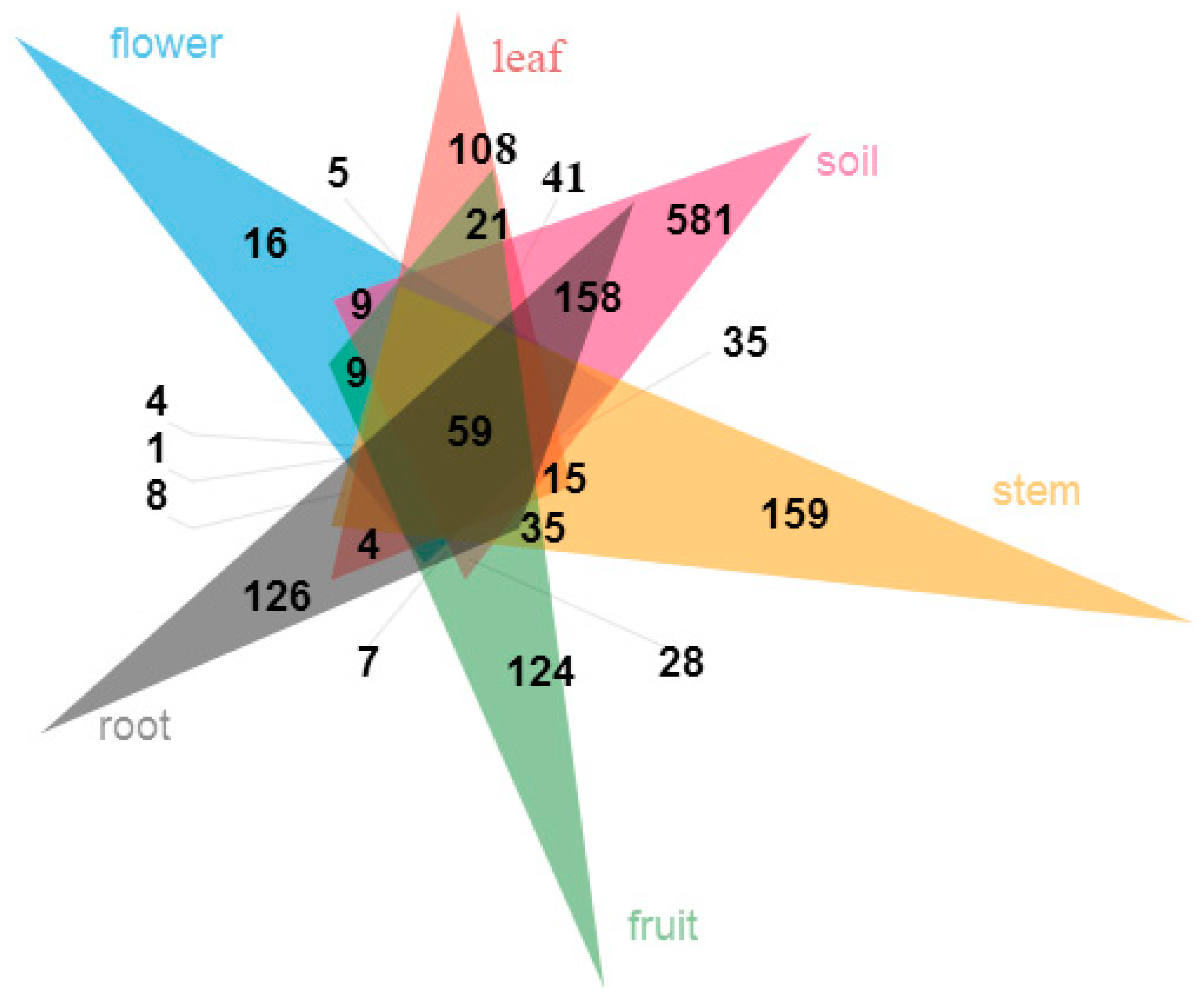
3.3. Fungal Community Composition among Different Organs and Soils
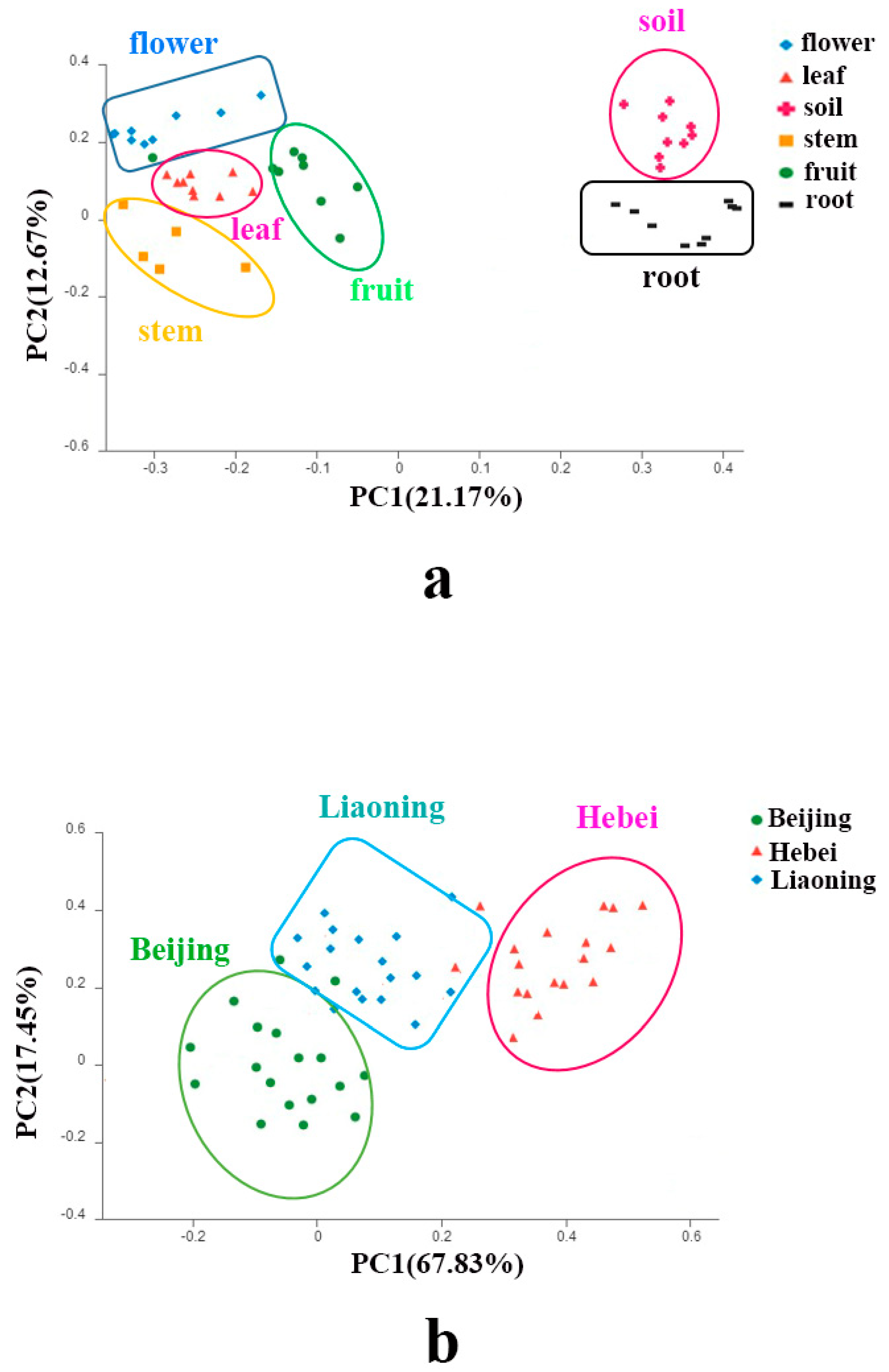
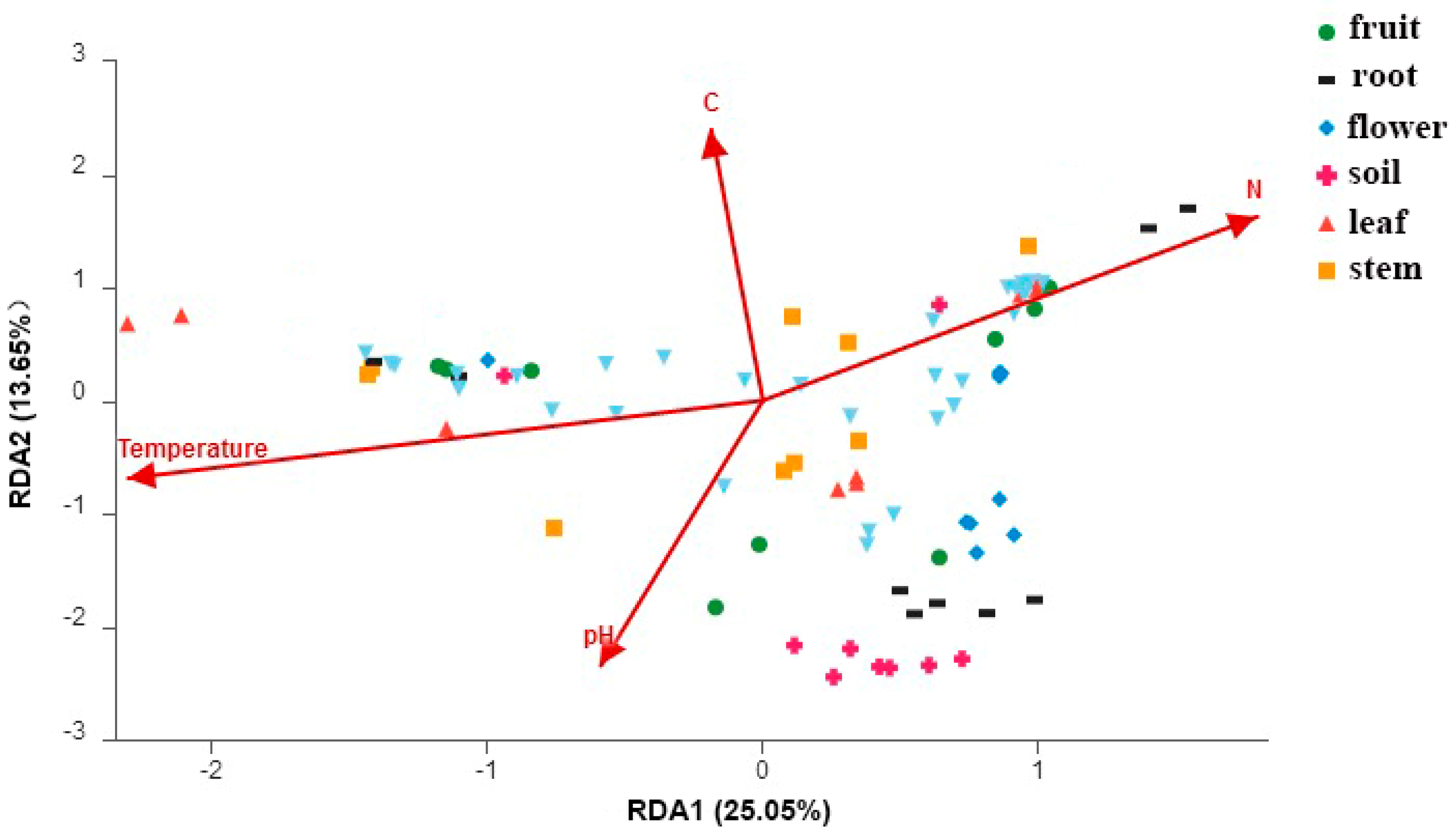
3.4. The Relationship between Fungal Community Structure and Organs, Sampling Sites and Soil Properties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Ethics
Conflicts of Interest
References
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrini, O.; Stone, J.; Carroll, F.E. Endophytic fungi in evergreen shrubs in western Oregon: A preliminary study. Can. J. Bot. 1982, 60, 789–796. [Google Scholar] [CrossRef]
- Sun, X.; Guo, L.D.; Hyde, K.D. Community composition of endophytic fungi in Acer truncatum and their role in decomposition. Fungal Divers. 2011, 47, 85–95. [Google Scholar] [CrossRef]
- Jia, M.; Chen, L.; Xin, H.L.; Zheng, C.J.; Rahman, K.; Han, T.; Qin, L.P. A friendly relationship between endophytic fungi and medicinal plants: A systematic review. Front. Microbiol. 2016, 7, 906. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, J.L.; Maccheroni, W.; Pereira, J.A.; Araujo, W.L. Endophytic microorganisms: A review on insect control and recent advances on tropical plants. Electron. J. Biotechnol. 2000, 3, 15–16. [Google Scholar] [CrossRef]
- Muller, D.B.; Vogel, C.; Bai, Y.; Vorholt, J.A. The plant microbiota: Systems-level insights and perspectives. Annu. Rev. Genet. 2016, 50, 211–234. [Google Scholar] [CrossRef]
- Photita, W.; Lumyong, S.; Lumyong, P.; Hyde, K.D. Endophytic fungi of wild banana (Musa acuminata) at doi Suthep Pui National Park, Thailand. Mycol. Res. 2001, 105, 1508–1513. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Bayman, P. Hidden fungi, emergent properties: Endophytes and microbiomes. Annu. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef]
- Xu, J.; Ebada, S.S.; Proksch, P. Pestalotiopsis a highly creative genus: Chemistry and bioactivity of secondary metabolites. Fungal Divers. 2010, 44, 15–31. [Google Scholar] [CrossRef]
- Tejesvi, M.V.; Kajula, M.; Mattila, S.; Pirttilä, A.M. Bioactivity and genetic diversity of endophytic fungi in Rhododendron tomentosum Harmaja. Fungal Divers. 2011, 47, 97–107. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Q.Y.; Jia, M.; Ming, Q.L.; Yue, W.; Rahman, K.; Han, T. Endophytic fungi with antitumor activities: Their occurrence and anticancer compounds. Crit. Rev. Microbiol. 2016, 42, 454–473. [Google Scholar] [CrossRef] [PubMed]
- Saikkonen, K.; Saari, S.; Helander, M. Defensive mutualism between plants and endophytic fungi? Fungal Divers. 2010, 41, 101–113. [Google Scholar] [CrossRef]
- Yuan, Y.; Feng, H.; Wang, L.; Li, Z.; Shi, Y.; Zhao, L.; Zhu, H. Potential of endophytic fungi isolated from cotton roots for biological control against verticillium wilt disease. PLoS ONE 2017, 12, e0170557. [Google Scholar] [CrossRef] [PubMed]
- Purahong, W.; Hyde, K.D. Effects of fungal endophytes on grass and non-grass litter decomposition rates. Fungal Divers. 2011, 47, 1–7. [Google Scholar] [CrossRef]
- Larrán, S.; Mónaco, C. Status and progress of research in endophytes from agricultural crops in Argentina. In Management of Fungal Plant Pathogens; Arya, A., Perello, A.E., Eds.; CAB International: Cambridge, UK, 2010; Chapter 12; p. 149. [Google Scholar]
- Russo, M.L.; Pelizza, S.A.; Cabello, M.N.; Stenglein, S.A.; Vianna, M.F.; Scorsetti, A.C. Endophytic fungi from selected varieties of soybean (Glycine max L. Merr.) and corn (Zea mays L.) grown in an agricultural area of Argentina. Revista Argentina de Microbiología 2016, 48, 154–160. [Google Scholar] [CrossRef]
- Bezerra, J.D.; Nascimento, C.C.; Barbosa, R.D.N.; da Silva, D.C.; Svedese, V.M.; Silva-Nogueira, E.B.; Souza-Motta, C.M. Endophytic fungi from medicinal plant Bauhinia forficata: Diversity and biotechnological potential. Braz. J. Microbiol. 2015, 46, 49–57. [Google Scholar] [CrossRef]
- Giordano, L.; Gonthier, P.; Varese, G.C.; Miserere, L.; Nicolotti, G. Mycobiota inhabiting sapwood of healthy and declining Scots pine (Pinus sylvestris L.) trees in the Alps. Fungal Divers. 2009, 38, 69–83. [Google Scholar]
- Kovalchuk, A.; Mukrimin, M.; Zeng, Z.; Raffaello, T.; Liu, M.X.; Kasanen, R.; Sun, H.; Asiegbu, F.O. Mycobiome analysis of asymptomatic and symptomatic Norway spruce trees naturally infected by the conifer pathogens Heterobasidion spp. Environ. Microbiol. Rep. 2018, 10, 532–541. [Google Scholar] [CrossRef]
- Johnston, P.R. Endophytes of apple and kiwifruit. In Proceedings of the 47th New Zealand Plant Protection Conference, Waitangi, New Zealand, 9–11 August 1994; New Zealand Plant Protection Society Inc.: Rotorua, New Zealand, 1994; p. 353. [Google Scholar]
- Camatti-Sartori, V.; da Silva-Ribeiro, R.T.; Valdebenito-Sanhueza, R.M.; Pagnocca, F.C.; Echeverrigaray, S.; Azevedo, J.L. Endophytic yeasts and filamentous fungi associated with southern Brazilian apple (Malus domestica) orchards subjected to conventional, integrated or organic cultivation. J. Basic Microb. 2005, 45, 397–402. [Google Scholar] [CrossRef]
- Photita, W.; Lumyong, S.; Lumyong, P.; McKenzie, E.H.C.; Hyde, K.D. Are some endophytes of Musa acuminata latent pathogens? Fungal Divers. 2004, 16, 131–140. [Google Scholar]
- Hortova, B.; Novotny, D. Endophytic fungi in branches of sour cherry trees: A preliminary study. Czech Mycol. 2011, 63, 77–82. [Google Scholar]
- Janisiewicz, W.J.; Jurick, W.M.; Peter, K.A.; Kurtzman, C.P.; Buyer, J.S. Yeasts associated with plums and their potential for controlling brown rot after harvest. Yeast 2014, 31, 207–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novotny, D. Studium endofytickych hub zemedelsky vyznamnch rostlin. Sbornik prispevku z workshopu Micromyco 2007, 4, 9. [Google Scholar]
- Sun, X.; Guo, L.D. Endophytic fungal diversity: Review of traditional and molecular techniques. Mycology 2012, 3, 65–76. [Google Scholar]
- Pinto, C.; Pinho, D.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Unravelling the diversity of grapevine microbiome. PLoS ONE 2014, 9, e85622. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Rybakova, D.; Grube, M.; Koberl, M. The plant microbiome explored: Implications for experimental botany. J. Exp. Bot. 2016, 67, 995–1002. [Google Scholar] [CrossRef]
- Dong, G.L. Cultivation Technology of Jing Bai Pear; Capital Normal University Press: Beijing, China, 2010; pp. 1–187. (In Chinese) [Google Scholar]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttila, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Abdelfattah, A.; Norelli, J.; Burchard, E.; Schena, L.; Droby, S.; Wisniewski, M. Apple endophytic microbiota of different rootstock/scion combinations suggests a genotype–specific influence. Microbiome 2018, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Vepštaitė-Monstavičė, I.; Lukša, J.; Stanevičienė, R.; Strazdaitė-Žielienė, Ž.; Yurchenko, V.; Serva, S.; Servienė, E. Distribution of apple and blackcurrant microbiota in Lithuania and the Czech Republic. Microbiol. Res. 2018, 206, 1–8. [Google Scholar] [CrossRef]
- Ren, F.; Dong, W.; Yan, D.H. Endophytic bacterial communities of Jingbai Pear trees in north China analyzed with Illumina sequencing of 16S rDNA. Arch. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis Part 3: Chemical Methods (Methodsofsoilan3); Sparks, D.L., Ed.; Soil Science Society of America Publisher Inc.: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Bremner, J.M. Total Nitrogen. In Methods of Soil Analysis Part 2: Chemical and Microbial Properties; Black, C.A., Ed.; American Society of Agronomy Publisher Inc.: Madison, WI, USA, 1965; pp. 1049–1178. [Google Scholar]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant. Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Lanzén, A.; Epelde, L.; Blanco, F.; Martín, I.; Artetxe, U.; Garbisuet, C. Multi–targeted metagenetic analysis of the influence of climate and environmental parameters on soil microbial communities along an elevational gradient. Sci. Rep. 2016, 6, e28257. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Gevers, D.; Westcott, S.L. Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA–based studies. PLoS ONE 2011, 6, e27310. [Google Scholar] [CrossRef] [PubMed]
- UNITE Community. UNITE Mothur Release. Version 01.12.2017. UNITE Community 2017. [Google Scholar] [CrossRef]
- Mothur Calculators. Available online: https://www.mothur.org/wiki/Calculators (accessed on 1 December 2018).
- R Core Team. R: A Language and environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Sinclair, J.B.; Cerkauskas, R.F. Latent infection vs. endophytic colonization by fungi. In Endophytic Fungi in Grasses and Woody Plants: Systematics, Ecology and Evolution; APS Press: St Paul, MN, USA, 1996; pp. 3–29. [Google Scholar]
- Botella, L.; Diez, J.J. Phylogenic diversity of fungal endophytes in Spanish stands of Pinus halepensis. Fungal Divers. 2011, 47, 9–18. [Google Scholar] [CrossRef]
- Pawłowska, J.; Wilk, M.; Śliwińska-Wyrzychowska, A.; Mętrak, M.; Wrzosek, M. The diversity of endophytic fungi in the above–ground tissue of two Lycopodium species in Poland. Symbiosis 2014, 63, 87–97. [Google Scholar] [CrossRef]
- Nelsen, M.P.; Lücking, R.; Grube, M.; Mbatchou, J.S.; Muggia, L.; Plata, E.R.; Lumbsch, H.T. Unravelling the phylogenetic relationships of lichenised fungi in Dothideomyceta. Stud. Mycol. 2009, 64, 135–144. [Google Scholar] [CrossRef]
- Robbertse, B.; Reeves, J.B.; Schoch, C.L.; Spatafora, J.W. A phylogenomic analysis of the Ascomycota. Fungal Genet. Biol. 2006, 43, 715–725. [Google Scholar] [CrossRef]
- Higgins, K.L.; Arnold, A.E.; Miadlikowska, J.; Sarvate, S.D.; Lutzoni, F. Phylogenetic relationships, host affinity, and geographic structure of boreal and arctic endophytes from three major plant lineages. Mol. Phylogenet. Evol. 2007, 42, 543–555. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.; Hyde, K.D.; Jones, E.G.; McKenzie, E.H.; Huang, S.K.; Abdel-Wahab, M.A.; Hongsanan, S. Towards a natural classification and backbone tree for Sordariomycetes. Fungal Divers. 2015, 72, 199–301. [Google Scholar] [CrossRef]
- Abutaha, N.; Mashaly, A.M.; Al-Mekhlafi, F.A.; Farooq, M.; Al-shami, M.; Wadaan, M.A. Larvicidal activity of endophytic fungal extract of Cochliobolus spicifer (Pleosporales: Pleosporaceae) on Aedes caspius and Culex pipiens (Diptera: Culicidae). Appl. Entomol. Zool. 2015, 50, 405–414. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Thambugala, K.M.; Manamgoda, D.S.; Jayawardena, R.; Camporesi, E.; Boonmee, S.; Chukeatirote, E. Towards a natural classification and backbone tree for Pleosporaceae. Fungal Divers. 2015, 71, 85–139. [Google Scholar] [CrossRef]
- McGee, P.A.; Markovina, A.L.; Jeong, G.C.; Cooper, E.D. Trichocomaceae in bark survive high temperatures and fire. FEMS Microbiol. Ecol. 2006, 56, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Leite, D.P.J.; Yamamoto, A.C.; Amadio, J.V.; Martins, E.R.; do Santos, F.A.; Simões Sde, A.; Hahn, R.C. Trichocomaceae: Biodiversity of Aspergillus spp. and Penicillium spp. residing in libraries. J. Infect. Dev. Ctries. 2012, 6, 734–743. [Google Scholar]
- Dugan, F.M.; Roberts, R.G. Etiology of preharvest colonization of Bing cherry fruit by fungi. Phytopathology 1994, 84, 1031–1036. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, H.X.; Chen, Y.C.; Sun, Z.H.; Li, H.H.; Li, S.N.; Yan, M.L.; Zhang, W.M. Two new metabolites from the endophytic fungus Alternaria sp. A744 derived from Morinda officinalis. Molecules 2017, 22, 765. [Google Scholar] [CrossRef]
- Xiang, L.; Gong, S.; Yang, L.; Hao, J.; Xue, M.; Zeng, F.; Yu, D. Biocontrol potential of endophytic fungi in medicinal plants from Wuhan Botanical Garden in China. Biol. Control 2016, 94, 47–55. [Google Scholar] [CrossRef]
- Yilmaz, N.; Visagie, C.M.; Frisvad, J.C.; Houbraken, J.; Jacobs, K.; Samson, R.A. Taxonomic re–evaluation of species in Talaromyces section Islandici, using a polyphasic approach. Persoonia 2016, 36, 37–56. [Google Scholar] [CrossRef]
- Nicoletti, R.; Salvatore, M.M.; Andolfi, A. Secondary Metabolites of Mangrove–Associated Strains of Talaromyces. Mar. Drugs 2018, 16, 12. [Google Scholar] [CrossRef]
- Coda, R.; Rizzello, C.G.; Cagno, R.D.; Trani, A.; Cardinali, G.; Gobbetti, M. Antifungal activity of Meyerozyma guilliermondii: Identification of active compounds synthesized during dough fermentation and their effect on long-term storage of wheat bread. Food Microbiol. 2013, 33, 243–251. [Google Scholar] [CrossRef]
- Nakayan, P.; Hameed, A.; Singh, S.; Young, L.S.; Hung, M.H.; Young, C.C. Phosphate-solubilizing soil yeast Meyerozyma guilliermondii cc1 improves maize (Zea mays L.) productivity and minimizes requisite chemical fertilization. Plant Soil 2013, 373, 301–315. [Google Scholar] [CrossRef]
- Kwon, Y.J.; Sohn, M.J.; Zheng, C.J.; Kim, W.G. Fumimycin: A peptide deformylase inhibitor with an unusual skeleton produced by Aspergillus fumisynnematus. Org. Lett. 2007, 9, 2449–2451. [Google Scholar] [CrossRef]
- Takemoto, K.; Kamisuki, S.; Chia, P.T.; Kuriyama, I.; Mizushina, Y.; Sugawara, F. Bioactive dihydronaphthoquinone derivatives from Fusarium solani. J. Nat. Prod. 2014, 77, 1992–1996. [Google Scholar] [CrossRef]
- Bezos, D.; Martínez-Álvarez, P.; Sanz-Ros, A.V.; MartínGarcía, J.; Fernandez, M.M.; Diez, J.J. Fungal Communities Associated with BarkBeetles in Pinus radiata Plantations in Northern Spain Affected by Pine Pitch Canker, with Special Focus on Fusarium Species. Forests 2018, 9, 698. [Google Scholar] [CrossRef]
- Martín-Pinto, P.; Pajares, J.A.; Nanos, N.; Diez, J.J. Site and seasonal influences on the fungal community on leaves and stems of Pinus and Quercus seedlings in forest nurseries. Sydowia 2004, 56, 243–257. [Google Scholar]
- Pinto, C.; Pinho, D.; Cardoso, R.; Custodio, V.; Fernandes, J.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Wine fermentation microbiome: A landscape from different Portuguese wine appellations. Front. Microbiol. 2015, 6, 905. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Thorngate, J.H.; Richardson, P.M.; Mills, D.A. Microbial biogeography of wine grapes is conditioned by cultivar, vintage, and climate. Proc. Natl. Acad. Sci. USA 2014, 111, E139–E148. [Google Scholar] [CrossRef]
- Bezemer, T.M.; Lawson, C.S.; Hedlund, K.; Edwards, A.R.; Brook, A.J.; Igual, J.M.; Mortimer, S.R.; Van Der Putten, W.H. Plant species and functional group effects on abiotic and microbial soil properties and plant–soil feedback responses in two grasslands. J. Ecol. 2006, 94, 893–904. [Google Scholar] [CrossRef]
- Neumann, G.; Bott, S.; Ohler, M.; Mock, H.P.; Lippmann, R.; Grosch, R.; Smalla, K. Root exudation and root development of lettuce (Lactuca sativa L. cv. Tizian) as affected by different soils. Front. Microbiol. 2014, 5, 2. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Sobs (Richness) | Invsimpson (Diversity) | Simpsoneven (Evenness) |
|---|---|---|---|
| leaf | 109.78 ± 7.37 | 5.92 ± 0.27 | 0.074 ± 0.006 |
| flower | 64.22 ± 1.55 | 2.83 ± 0.08 | 0.047 ± 0.002 |
| fruit | 115.56 ± 6.03 | 5.98 ± 0.35 | 0.057 ± 0.003 |
| stem | 120.00 ± 6.84 | 4.58 ± 0.19 | 0.048 ± 0.004 |
| root | 159.33 ± 6.27 | 8.95 ± 0.54 | 0.057 ± 0.003 |
| soil | 487.44 ± 15.82 | 9.95 ± 1.24 | 0.018 ± 0.002 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, F.; Dong, W.; Sun, H.; Yan, D.-H. Endophytic Mycobiota of Jingbai Pear Trees in North China. Forests 2019, 10, 260. https://doi.org/10.3390/f10030260
Ren F, Dong W, Sun H, Yan D-H. Endophytic Mycobiota of Jingbai Pear Trees in North China. Forests. 2019; 10(3):260. https://doi.org/10.3390/f10030260
Chicago/Turabian StyleRen, Fei, Wei Dong, Hui Sun, and Dong-Hui Yan. 2019. "Endophytic Mycobiota of Jingbai Pear Trees in North China" Forests 10, no. 3: 260. https://doi.org/10.3390/f10030260
APA StyleRen, F., Dong, W., Sun, H., & Yan, D. -H. (2019). Endophytic Mycobiota of Jingbai Pear Trees in North China. Forests, 10(3), 260. https://doi.org/10.3390/f10030260