Bird Communities in Urban Riparian Areas: Response to the Local- and Landscape-Scale Environmental Variables

Abstract
:1. Introduction
- (1)
- Clarify the avian biodiversity in urban riparian areas of the Tsing River.
- (2)
- Identify the influence of environmental variables on avian biodiversity.
- (3)
- Obtain a species-specific understanding of (2) by including biological characteristics.
2. Methods
2.1. Study Area
2.2. Bird Surveys and Biodiversity Metrics
2.3. Environmental Variables and Data Collection
2.4. Preselection of Variables and Statistical Analyses
3. Results
3.1. Observed Birds and the Selected Variables
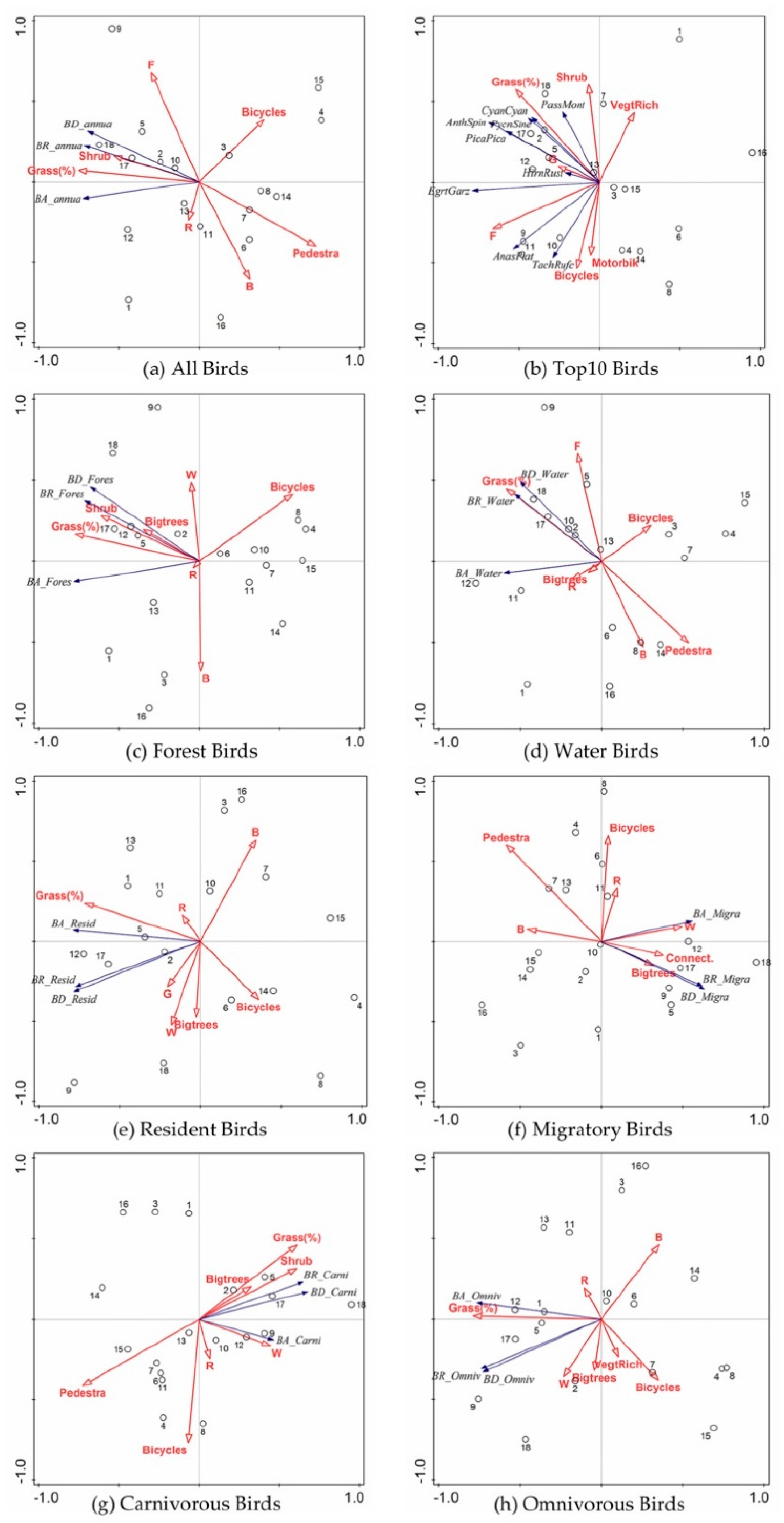
3.2. Overall Responses of Birds
3.3. Species Specific Responses of Birds
4. Discussion
4.1. The Scale of Effect of Landscape-Scale Variables
4.2. What Can Be Concluded from the Responses of Migratory Birds?
4.3. The Dispersal Distance of Birds and Landscape Connectivity
4.4. Significant Variables and Implication for Bird Conservation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Humphrey, J.W.; Watts, K.; Fuentes-Montemayor, E.; Macgregor, N.A.; Peace, A.J.; Park, K.J. What can studies of woodland fragmentation and creation tell us about ecological networks? A literature review and synthesis. Landsc. Ecol. 2015, 30, 21–50. [Google Scholar] [CrossRef]
- Ortega-Álvarez, R.; MacGregor-Fors, I. Living in the big city: Effects of urban land-use on bird community structure, diversity, and composition. Landsc. Urban Plan. 2009, 90, 189–195. [Google Scholar] [CrossRef]
- Haedo, J.; Gioia, A.; Aráoz, E.; Paolini, L.; Malizia, A. Primary productivity in cities and their influence over subtropical bird assemblages. Urban For. Urban Green. 2017, 26, 57–64. [Google Scholar] [CrossRef]
- Jokimäki, J.; Suhonen, J.; Kaisanlahti-Jokimäki, M.L.; Jukka, S.; Marja-Liisa, K.J. Urban core areas are important for species conservation: A European-level analysis of breeding bird species. Landsc. Urban Plan. 2018, 178, 73–81. [Google Scholar] [CrossRef]
- Liang, J.; Xing, W.; Zeng, G.; Li, X.; Peng, Y.; Li, X.; Gao, X.; He, X. Where will threatened migratory birds go under climate change? Implications for China’s national nature reserves. Sci. Total Environ. 2018, 645, 1040–1047. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, R.; Lindenmayer, D.; Barton, P.; Ikin, K.; Crane, M.; Michael, D.; Okada, S.; Gibbons, P.; Stein, J. Cross-sectional and temporal relationships between bird occupancy and vegetation cover at multiple spatial scales. Ecol. Appl. 2014, 24, 1275–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, E.E.; Maron, M. Does the response of bird assemblages to fire mosaic properties vary among spatial scales and foraging guilds? Landsc. Ecol. 2016, 31, 687–699. [Google Scholar] [CrossRef]
- Guttery, M.R.; Ribic, C.A.; Sample, D.W.; Paulios, A.; Trosen, C.; Dadisman, J.; Horton, J.A. Scale-specific habitat relationships influence patch occupancy: Defining neighborhoods to optimize the effectiveness of landscape-scale grassland bird conservation. Landsc. Ecol. 2017, 32, 515–529. [Google Scholar] [CrossRef]
- Mu, R.; Kumaraswamy, K.; Mohanraj, R. Environmental Management of River Basin Ecosystems; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Suri, J.; Anderson, P.M.; Charles-Dominique, T.; Hellard, E.; Cumming, G.S. More than just a corridor: A suburban river catchment enhances bird functional diversity. Landsc. Urban Plan. 2017, 157, 331–342. [Google Scholar] [CrossRef]
- Pennington, D.N.; Hansel, J.; Blair, R.B. The conservation value of urban riparian areas for landbirds during spring migration: Land cover, scale, and vegetation effects. Biol. Conserv. 2008, 141, 1235–1248. [Google Scholar] [CrossRef]
- Tiwary, N.; Urfi, A. Spatial variations of bird occupancy in Delhi: The significance of woodland habitat patches in urban centres. Urban For. Urban Green. 2016, 20, 338–347. [Google Scholar] [CrossRef]
- Threlfall, C.G.; Williams, N.S.; Hahs, A.K.; Livesley, S.J. Approaches to urban vegetation management and the impacts on urban bird and bat assemblages. Landsc. Urban Plan. 2016, 153, 28–39. [Google Scholar] [CrossRef]
- Huang, Y.; Zhao, Y.; Li, S.; Von Gadow, K. The Effects of habitat area, vegetation structure and insect richness on breeding bird populations in Beijing urban parks. Urban For. Urban Green. 2015, 14, 1027–1039. [Google Scholar] [CrossRef]
- Yang, G.; Xu, J.; Wang, Y.; Wang, X.; Pei, E.; Yuan, X.; Li, H.; Ding, Y.; Wang, Z. Evaluation of microhabitats for wild birds in a Shanghai urban area park. Urban For. Urban Green. 2015, 14, 246–254. [Google Scholar] [CrossRef]
- Sasaki, T.; Imanishi, J.; Fukui, W.; Morimoto, Y. Fine-scale characterization of bird habitat using airborne LiDAR in an urban park in Japan. Urban For. Urban Green. 2016, 17, 16–22. [Google Scholar] [CrossRef]
- Estevo, C.A.; Nagy-Reis, M.B.; Silva, W.R. Urban parks can maintain minimal resilience for Neotropical bird communities. Urban For. Urban Green. 2017, 27, 84–89. [Google Scholar] [CrossRef]
- Morelli, F.; Benedetti, Y.; Su, T.; Zhou, B.; Moravec, D.; Šímová, P.; Liang, W. Taxonomic diversity, functional diversity and evolutionary uniqueness in bird communities of Beijing’s urban parks: Effects of land use and vegetation structure. Urban For. Urban Green. 2017, 23, 84–92. [Google Scholar] [CrossRef]
- Steel, Z.L.; Steel, A.E.; Williams, J.N.; Viers, J.H.; Marquet, P.A.; Barbosa, O. Patterns of bird diversity and habitat use in mixed vineyard-matorral landscapes of Central Chile. Ecol. Indic. 2017, 73, 345–357. [Google Scholar] [CrossRef]
- Morelli, F. High nature value farmland increases taxonomic diversity, functional richness and evolutionary uniqueness of bird communities. Ecol. Indic. 2018, 90, 540–546. [Google Scholar] [CrossRef]
- Canedoli, C.; Manenti, R.; Padoa-Schioppa, E. Birds biodiversity in urban and periurban forests: Environmental determinants at local and landscape scales. Urban Ecosyst. 2018, 21, 779–793. [Google Scholar] [CrossRef]
- Banville, M.J.; Bateman, H.L.; Earl, S.R.; Warren, P.S. Decadal declines in bird abundance and diversity in urban riparian zones. Landsc. Urban Plan. 2017, 159, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Barton, P.S.; Ikin, K.; Smith, A.L.; MacGregor, C.; Lindenmayer, D.B. Vegetation structure moderates the effect of fire on bird assemblages in a heterogeneous landscape. Landsc. Ecol. 2014, 29, 703–714. [Google Scholar] [CrossRef]
- Mammides, C.; Kadis, C.; Coulson, T. The effects of road networks and habitat heterogeneity on the species richness of birds in Natura 2000 sites in Cyprus. Landsc. Ecol. 2015, 30, 67–75. [Google Scholar] [CrossRef]
- Chambers, C.L.; Cushman, S.A.; Medina-Fitoria, A.; Martínez-Fonseca, J.; Chávez-Velásquez, M. Influences of scale on bat habitat relationships in a forested landscape in Nicaragua. Landsc. Ecol. 2016, 31, 1299–1318. [Google Scholar] [CrossRef]
- Fartmann, T.; Kämpfer, S.; Brüggeshemke, J.; Juchem, M.; Klauer, F.; Weking, S.; Löffler, F. Landscape-scale effects of Christmas-tree plantations in an intensively used low-mountain landscape–Applying breeding bird assemblages as indicators. Ecol. Indic. 2018, 94, 409–419. [Google Scholar] [CrossRef]
- Salgueiro, P.A.; Mira, A.; Rabaça, J.E.; Santos, S.M. Identifying critical thresholds to guide management practices in agro-ecosystems: Insights from bird community response to an open grassland-to-forest gradient. Ecol. Indic. 2018, 88, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Galitsky, C.; Lawler, J.J. Relative influence of local and landscape factors on bird communities vary by species and functional group. Landsc. Ecol. 2015, 30, 287–299. [Google Scholar] [CrossRef]
- Taylor, J.J.; Lepczyk, C.A.; Brown, D.G. Patch and matrix level influences on forest birds at the rural–urban interface. Landsc. Ecol. 2016, 31, 1005–1020. [Google Scholar] [CrossRef]
- O’Neill, R.V.; Deangelis, D.L.; Waide, J.B.; Allen, T.F.; Allen, G.E. A Hierarchical Concept of Ecosystems (No. 23); Princeton University Press: Princeton, NJ, USA, 1986. [Google Scholar]
- Rukke, B.A.; Midtgaard, F. The importance of scale and spatial variables for the fungivorous beetle Bolitophagus reticulatus (Coleoptera, Tenebrionidae) in a fragmented forest landscape. Ecography 1998, 21, 561–572. [Google Scholar] [CrossRef]
- Dorresteijn, I.; Teixeira, L.; Von Wehrden, H.; Loos, J.; Hanspach, J.; Stein, J.A.R.; Fischer, J. Impact of land cover homogenization on the Corncrake (Crex crex) in traditional farmland. Landsc. Ecol. 2015, 30, 1483–1495. [Google Scholar] [CrossRef]
- Reijnen, R.; Foppen, R.; Ter Braak, C.; Thissen, J. The Effects of Car Traffic on Breeding Bird Populations in Woodland. III. Reduction of Density in Relation to the Proximity of Main Roads. J. Appl. Ecol. 1995, 32, 187. [Google Scholar] [CrossRef]
- Forman, R.T.T.; Reineking, B.; Hersperger, A.M. Road Traffic and Nearby Grassland Bird Patterns in a Suburbanizing Landscape. Environ. Manag. 2002, 29, 782–800. [Google Scholar] [CrossRef]
- Uuemaa, E.; Mander, U.; Marja, R. Trends in the use of landscape spatial metrics as landscape indicators: A review. Ecol. Indic. 2013, 28, 100–106. [Google Scholar] [CrossRef]
- Vihervaara, P.; Mononen, L.; Auvinen, A.P.; Virkkala, R.; Lü, Y.; Pippuri, I.; Valkama, J. How to integrate remotely sensed data and biodiversity for ecosystem assessments at landscape scale. Landsc. Ecol. 2015, 30, 501–516. [Google Scholar] [CrossRef]
- Tannier, C.; Foltête, J.C.; Girardet, X. Assessing the capacity of different urban forms to preserve the connectivity of ecological habitats. Landsc. Urban Plan. 2012, 105, 128–139. [Google Scholar] [CrossRef]
- Foltête, J.C.; Clauzel, C.; Vuidel, G. A software tool dedicated to the modelling of landscape networks. Environ. Model. Softw. 2012, 38, 316–327. [Google Scholar] [CrossRef]
- Buelow, C.A.; Baker, R.; Reside, A.E.; Sheaves, M. Spatial dynamics of coastal forest bird assemblages: The influence of landscape context, forest type, and structural connectivity. Landsc. Ecol. 2017, 32, 547–561. [Google Scholar] [CrossRef]
- Sahraoui, Y.; Foltête, J.C.; Clauzel, C. A multi-species approach for assessing the impact of land-cover changes on landscape connectivity. Landsc. Ecol. 2017, 32, 1819–1835. [Google Scholar] [CrossRef]
- Xu, X.; Xie, Y.; Qi, K.; Luo, Z.; Wang, X. Detecting the response of bird communities and biodiversity to habitat loss and fragmentation due to urbanization. Sci. Total Environ. 2018, 624, 1561–1576. [Google Scholar] [CrossRef]
- Forman, R.T.; Collinge, S.K. Nature conserved in changing landscapes with and without spatial planning. Landsc. Urban Plan. 1997, 37, 129–135. [Google Scholar] [CrossRef]
- Mimet, A.; Clauzel, C.; Foltête, J.C. Locating wildlife crossings for multispecies connectivity across linear infrastructures. Landsc. Ecol. 2016, 31, 1955–1973. [Google Scholar] [CrossRef]
- Lambeck, R.J. Focal Species: A Multi-Species Umbrella for Nature Conservation. Especies Focales: Una Sombrilla Multiespecifica para Conservar la Naturaleza. Conserv. Biol. 1997, 11, 849–856. [Google Scholar] [CrossRef] [Green Version]
- Silvano, A.L.; Guyer, C.; Steury, T.D.; Grand, J.B. Selecting focal species as surrogates for imperiled species using relative sensitivities derived from occupancy analysis. Ecol. Indic. 2017, 73, 302–311. [Google Scholar] [CrossRef]
- Loman, Z.G.; Deluca, W.V.; Harrison, D.J.; Loftin, C.S.; Rolek, B.W.; Wood, P.B. Landscape capability models as a tool to predict fine-scale forest bird occupancy and abundance. Landsc. Ecol. 2018, 33, 77–91. [Google Scholar] [CrossRef]
- Miguet, P.; Jackson, H.B.; Jackson, N.D.; Martin, A.E.; Fahrig, L. What determines the spatial extent of landscape effects on species? Landsc. Ecol. 2016, 31, 1177–1194. [Google Scholar] [CrossRef]
- MacKinnon, J.R.; MacKinnon, J.; Phillipps, K.; He, F.Q. A Field Guide to the Birds of China; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Shih, W.Y. Bird diversity of greenspaces in the densely developed city centre of Taipei. Urban Ecosyst. 2018, 21, 379–393. [Google Scholar] [CrossRef]
- Alexandrino, E.R.; Buechley, E.R.; Piratelli, A.J.; de Barros Ferraz, K.M.P.M.; Moral, R.D.A.; Şekercioğlu, Ç.H.; Silva, W.R.; do Couto, H.T.Z. Bird sensitivity to disturbance as an indicator of forest patch conditions: An issue in environmental assessments. Ecol. Indic. 2016, 66, 369–381. [Google Scholar] [CrossRef] [Green Version]
- Kettig, R.; Landgrebe, D. Classification of Multispectral Image Data by Extraction and Classification of Homogeneous Objects. IEEE Trans. Geosci. Electron. 1976, 14, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Vieira, M.A.; Formaggio, A.R.; Rennó, C.D.; Atzberger, C.; Aguiar, D.A.; Mello, M.P. Object Based Image Analysis and Data Mining applied to a remotely sensed Landsat time-series to map sugarcane over large areas. Remote Sens. Environ. 2012, 123, 553–562. [Google Scholar] [CrossRef]
- Saura, S.; Pascual-Hortal, L. A new habitat availability index to integrate connectivity in landscape conservation planning: Comparison with existing indices and application to a case study. Landsc. Urban Plan. 2007, 83, 91–103. [Google Scholar] [CrossRef]
- Saura, S.; Torné, J. Conefor Sensinode 2.2: A software package for quantifying the importance of habitat patches for landscape connectivity. Environ. Model. Softw. 2009, 24, 135–139. [Google Scholar] [CrossRef]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Grafius, D.R.; Corstanje, R.; Siriwardena, G.M.; Plummer, K.E.; Harris, J.A. A bird’s eye view: Using circuit theory to study urban landscape connectivity for birds. Landsc. Ecol. 2017, 32, 1771–1787. [Google Scholar] [CrossRef]
- Moudrý, V.; Komárek, J.; Šímová, P. Which breeding bird categories should we use in models of species distribution? Ecol. Indic. 2017, 74, 526–529. [Google Scholar] [CrossRef]
- Hostetler, M. Scale, birds, and human decisions: A potential for integrative research in urban ecosystems. Landsc. Urban Plan. 1999, 45, 15–19. [Google Scholar] [CrossRef]
- Bernard, G.E.; Van Dongen, W.F.; Guay, P.J.; Symonds, M.R.; Robinson, R.W.; Weston, M.A. Bicycles evoke longer flight-initiation distances and higher intensity escape behaviour of some birds in parks compared with pedestrians. Landsc. Urban Plan. 2018, 178, 276–280. [Google Scholar] [CrossRef]
- Guo, S.; Saito, K.; Yin, W.; Su, C. Landscape Connectivity as a Tool in Green Space Evaluation and Optimization of the Haidan District, Beijing. Sustainability 2018, 10, 1979. [Google Scholar] [CrossRef]
- Pei, N.; Wang, C.; Jin, J.; Jia, B.; Chen, B.; Qie, G.; Qiu, E.; Gu, L.; Sun, R.; Li, J.; et al. Long-term afforestation efforts increase bird species diversity in Beijing, China. Urban For. Urban Green. 2018, 29, 88–95. [Google Scholar] [CrossRef]
- Groffman, P.M.; Baron, J.S.; Blett, T.; Gold, A.J.; Goodman, I.; Gunderson, L.H.; Levinson, B.M.; Palmer, M.A.; Paerl, H.W.; Peterson, G.D.; et al. Ecological Thresholds: The Key to Successful Environmental Management or an Important Concept with No Practical Application? Ecosystems 2006, 9, 1–13. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Explains % | Pseudo-F | p | |
|---|---|---|---|---|
| All birds | Grass (%) | 48.5 | 14.1 | 0.034 |
| Pedestrians | 11.7 | 4.1 | 0.074 | |
| Bicycles | 10.0 | 6.2 | 0.098 | |
| F | 5.5 | 4.6 | 0.080 | |
| Top 10 birds | F | 22.7 | 4.4 | 0.034 |
| Grass (%) | 18.2 | 4.3 | 0.054 | |
| Shrub | 11.3 | 3.1 | 0.022 | |
| Bicycles | 8.0 | 2.4 | 0.038 | |
| Veg. Richness | 6.2 | 2.0 | 0.066 | |
| Motorbikes/scooters | 7.3 | 2.8 | 0.052 | |
| Resident birds | Grass (%) | 45.4 | 12.5 | 0.034 |
| Bicycles | 13.9 | 7.9 | 0.038 | |
| B | 11.2 | 3.6 | 0.020 | |
| W | 6.6 | 5.0 | 0.052 | |
| Migratory birds | Pedestrians | 30.7 | 6.7 | 0.034 |
| W | 18.3 | 5.0 | 0.064 | |
| Connect.5km | 10.7 | 4.4 | 0.044 | |
| Big trees | 8.3 | 5.5 | 0.070 | |
| Bicycles | 3.1 | 2.3 | 0.074 | |
| Forest birds | Grass (%) | 49.1 | 14.5 | 0.034 |
| Bicycles | 12.9 | 4.8 | 0.020 | |
| Shrub | 9.6 | 4.4 | 0.052 | |
| R | 4.5 | 2.5 | 0.082 | |
| B | 6.8 | 5.4 | 0.052 | |
| Water birds | Grass (%) | 26.9 | 5.5 | 0.078 |
| F | 6.1 | 2.1 | 0.068 | |
| Pedestrians | 9.8 | 4.7 | 0.074 | |
| Carnivorous birds | Pedestrians | 39.7 | 9.9 | 0.034 |
| Bicycles | 6.2 | 2.5 | 0.096 | |
| Omnivorous birds | Grass (%) | 54.1 | 17.7 | 0.034 |
| B | 11.4 | 4.6 | 0.050 | |
| Bicycles | 9.9 | 6.9 | 0.038 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, S.; Su, C.; Saito, K.; Cheng, J.; Terada, T. Bird Communities in Urban Riparian Areas: Response to the Local- and Landscape-Scale Environmental Variables. Forests 2019, 10, 683. https://doi.org/10.3390/f10080683
Guo S, Su C, Saito K, Cheng J, Terada T. Bird Communities in Urban Riparian Areas: Response to the Local- and Landscape-Scale Environmental Variables. Forests. 2019; 10(8):683. https://doi.org/10.3390/f10080683
Chicago/Turabian StyleGuo, Shiyi, Chang Su, Kaoru Saito, Jiexin Cheng, and Toru Terada. 2019. "Bird Communities in Urban Riparian Areas: Response to the Local- and Landscape-Scale Environmental Variables" Forests 10, no. 8: 683. https://doi.org/10.3390/f10080683
APA StyleGuo, S., Su, C., Saito, K., Cheng, J., & Terada, T. (2019). Bird Communities in Urban Riparian Areas: Response to the Local- and Landscape-Scale Environmental Variables. Forests, 10(8), 683. https://doi.org/10.3390/f10080683