Does Ozone Alter the Attractiveness of Japanese White Birch Leaves to the Leaf Beetle Agelastica coerulea via Changes in Biogenic Volatile Organic Compounds (BVOCs): An Examination with the Y-Tube Test



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Insects
2.3. Olfactory Response Test (Y-Tube Test)
2.4. Sampling of BVOCs
2.5. Gas Chromatograph Mass Spectrometer (GC-MS) Analyses
2.6. Lifetime of BVOCs with O3
2.7. Statistical Analysis
3. Results
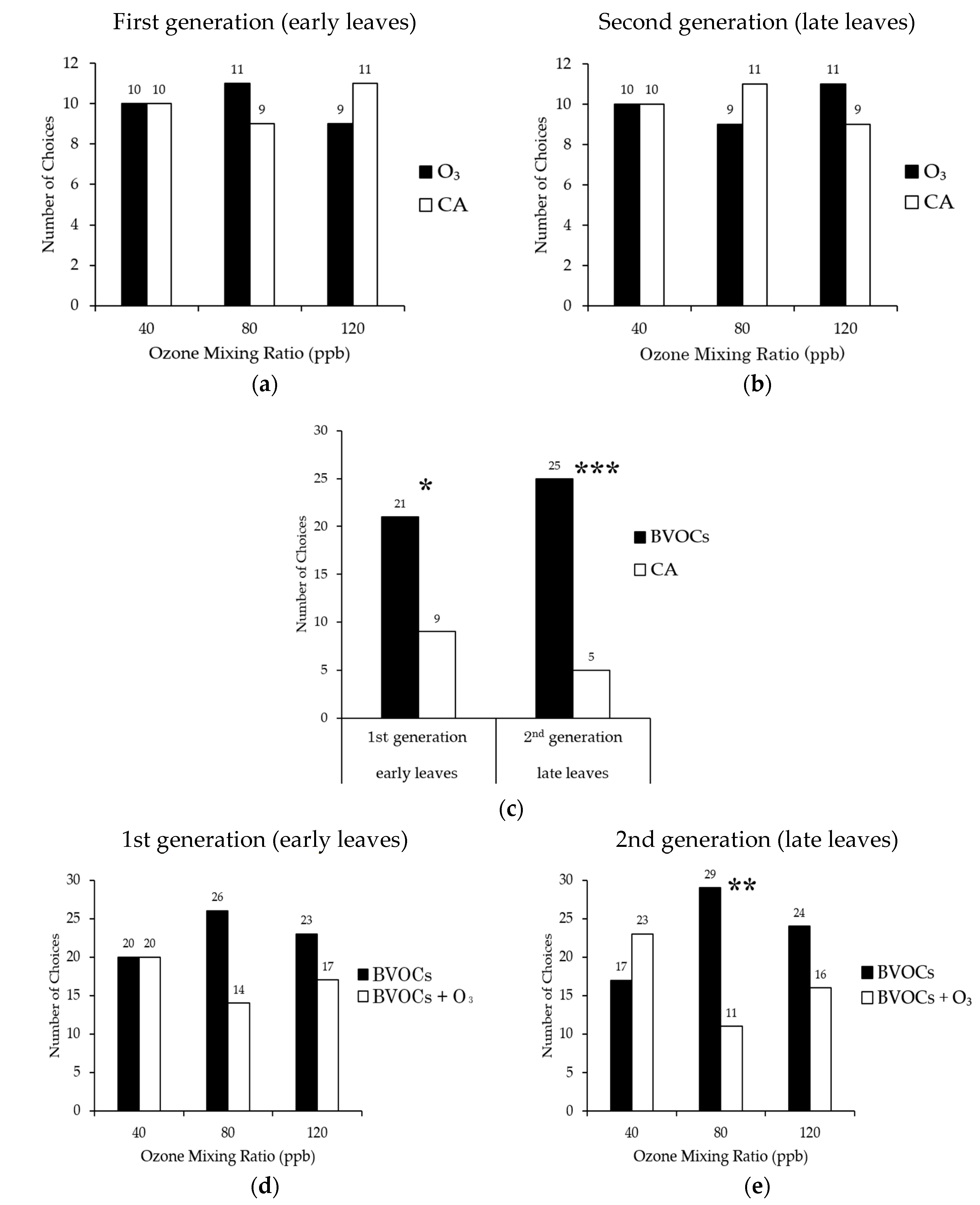
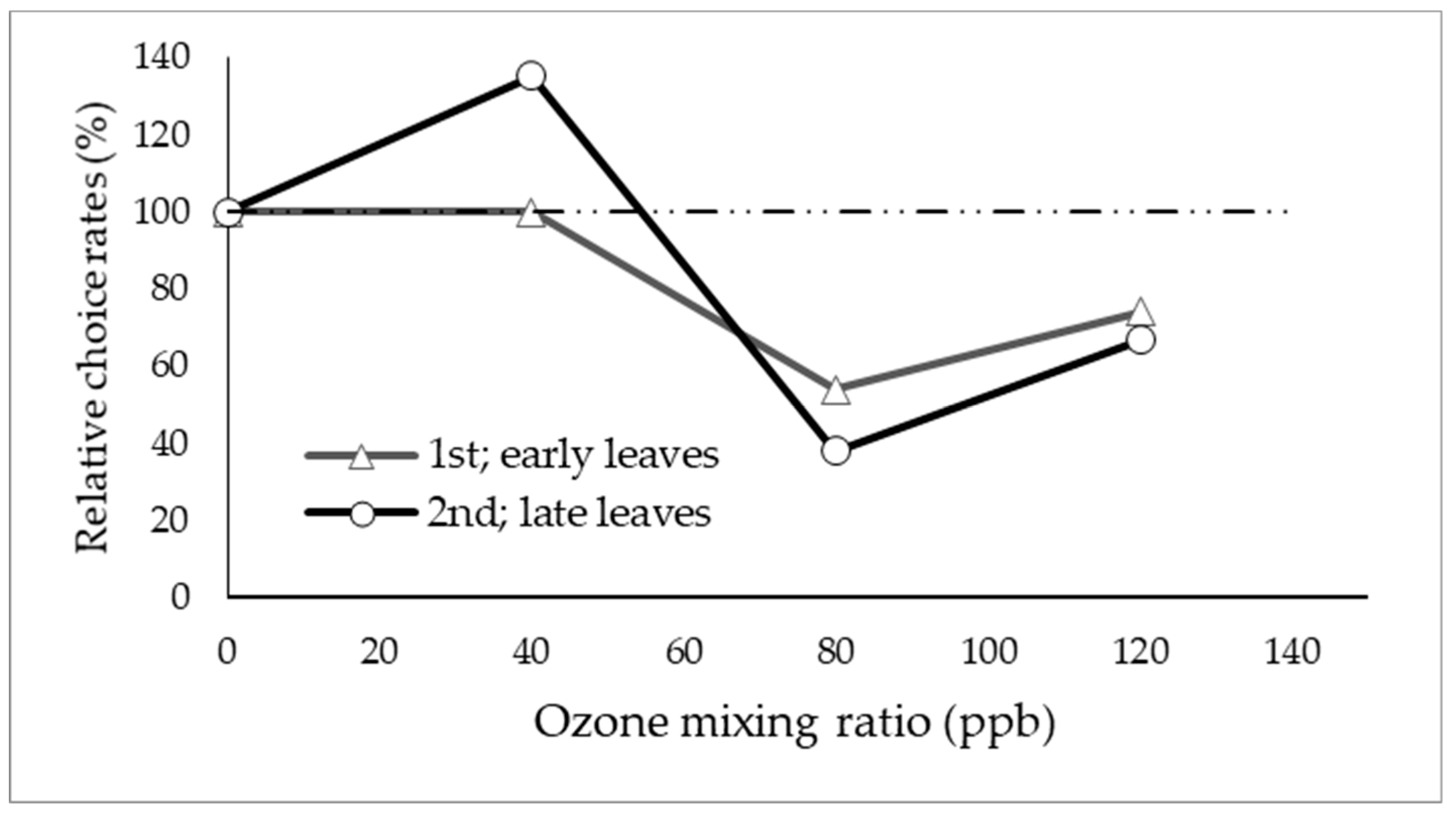
3.1. Olfactory Response Test (Y-Tube Test)
3.2. Sampling of BVOCs
3.2.1. BVOCs’ Emission Rates and Composition
3.2.2. O3 Reactive Compounds
4. Discussion
4.1. Behavior of Alder Leaf Beetles in High O3 Environment
4.2. Multiplicity of BVOCs
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Koike, T.; Watanabe, M.; Hoshika, Y.; Kitao, M.; Matsumura, H.; Funada, R.; Izuta, T. Effects of Ozone on Forest Ecosystems in East and Southeast Asia. Dev. Environ. Sci. 2013, 13, 371–390. [Google Scholar] [CrossRef]
- Akimoto, H.; Mori, Y.; Sasaki, K.; Nakanishi, H.; Ohizumi, T.; Itano, Y. Analysis of monitoring data of ground-level ozone in Japan for long- term trend during 1990–2010: Causes of temporal and spatial variation. Atmos. Environ. 2015, 102, 302–310. [Google Scholar] [CrossRef]
- Nagashima, T.; Sudo, K.; Akimoto, H.; Kurokawa, J.; Ohara, T. Long-term change in the source contribution to surface ozone over Japan. Atmos. Chem. Phys. 2017, 17, 8231–8246. [Google Scholar] [CrossRef] [Green Version]
- Matyssek, R.; Wieser, G.; Calfapietra, C.; De Vries, W.; Dizengremel, P.; Ernst, D.; Jolivet, Y.; Mikkelsen, T.N.; Mohren, G.M.J.; Le Thiec, D.; et al. Forests under climate change and air pollution: gaps in understanding and future directions for research. Environ. Pollut. 2012, 160, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Agathokleous, E.; Kitao, M.; Komatsu, M.; Tamai, Y.; Saito, H.; Harayama, H.; Uemura, A.; Tobita, H.; Koike, T. Effects of soil nutrient availability and ozone on container-grown Japanese larch seedlings and role of soil microbes. J. Forestry Res. 2019, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Matyssek, R.; Wieser, G.; Fleischmann, F.; Grűnhage, L. Ozone Research, Quo Vadis? Lessons from the Free-Air Canopy Fumigation Experiment at Kranzberg Forest. Dev. Environ. Sci. 2013, 13, 103–129. [Google Scholar] [CrossRef]
- Chappelka, A.H.; Grulke, N.E. Disruption of the ‘disease triangle’ by chemical and physical environmental change. Plant Biol. 2016, 18, 5–12. [Google Scholar] [CrossRef]
- Lindroth, R.L. Impacts of Elevated Atmospheric CO2 and O3 on forests: Phytochemistry, Trophic Interactions, and Ecosystem Dynamics. J. Chem. Ecol. 2010, 36, 2–21. [Google Scholar] [CrossRef]
- Gregg, J.W.; Jones, C.G.; Dawson, T.E. Urbanization effects on tree growth in the vicinity of New York City. Nature 2003, 424, 183–187. [Google Scholar] [CrossRef]
- Sakikawa, T.; Shi, C.; Nakamura, M.; Watanabe, M.; Oikawa, M.; Satoh, F.; Koike, T. Leaf phenology and insect grazing of Japanese white birch saplings grown under free-air ozone exposure. J. Agric. Meteorol. 2011, 72, 80–84. [Google Scholar] [CrossRef] [Green Version]
- Abu ElEla, S.; Agathokleous, E.; Ghazawy, N.A.; Amin, T.R.; ElSayed, W.M.; Koike, T. Enzyme activity modification in adult beetles (Agelastica coerulea) inhabiting birch trees in an ozone-enriched atmosphere. Environ. Sci. Pollut. Res. 2018, 25, 32675–32683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugai, T.; Okamoto, S.; Agathokleous, E.; Masui, N.; Satoh, F.; Koike, T. Leaf defense capacity of Japanese elm (Ulmus davidiana var. japonica) seedlings subjected to a nitrogen loading and insect herbivore dynamics in a free air ozone-enriched environment. Environ. Sci. Pollut. Res. 2019, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Agathokleous, E.; Sakikawa, T.; Abu ElEla, S.; Mochizuki, T.; Nakamura, M.; Watanabe, M.; Kawamura, K.; Koike, T. Ozone alters the feeding behavior of the leaf beetle Agelastica coerulea (Coleoptera: Chrysomelidae) into leaves of Japanese white birch (Betula platyphylla var. japonica). Environ. Sci. Pollut. Res. 2017, 24, 17577–17583. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, R.; Arey, J. Gas-phase tropospheric chemistry of biogenic volatile organic compounds: A review. Atmos. Environ. 2003, 37, S197–S219. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Blande, J.D. Where do herbivore-induced plant volatiles go? Front. Plant Sci. 2013, 4, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blande, J.D.; Holopainen, J.K.; Niinemets, Ü. Plant volatiles in polluted atmospheres: Stress responses and signal degradation. Plant. Cell. Environ. 2014, 37, 1892–1904. [Google Scholar] [CrossRef] [Green Version]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P. Protective and defensive roles of non-glandular trichomes against multiple stresses: Structure–function coordination. J. Forestry Res. 2019. [Google Scholar] [CrossRef] [Green Version]
- Roeland, S.; Moretti, M.; Amorim, J.H.; Branquinho, C.; Fares, S.; Morelli, F.; Niinemets, Ü.; Paoletti, E.; Pinho, P.; Sgrigna, G.; et al. Towards an integrative approach to evaluate the environmental ecosystem services provided by urban forest. J. Forestry Res. 2019, 30, 1981–1996. [Google Scholar] [CrossRef] [Green Version]
- Pinto, D.M.; Blande, J.D.; Souza, S.R.; Nerg, A.-M.; Holopainen, J.K. Plant Volatile Organic Compounds (VOCs) in Ozone (O3) Polluted Atmospheres: The Ecological Effects. J. Chem. Ecol. 2010, 36, 22–34. [Google Scholar] [CrossRef]
- Li, T.; Blande, J.D.; Holopainen, J.K. Atmospheric transformation of plant volatiles disrupts host plant finding. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Fuentes, J.D.; Lerdau, M.; Atkinson, R.; Baldocchi, D.; Bottenheim, J.W.; Ciccioli, P.; Lamb, B.; Geron, C.; Gu, L.; Guenther, A.; et al. Biogenic Hydrocarbons in the Atmospheric Boundary Layer: A Review. Bull. Amer. Meteor. 2000, 81, 1537–1575. [Google Scholar] [CrossRef] [Green Version]
- McFrederick, Q.S.; Kathilankal, J.C.; Fuentes, J.D. Air pollution modifies floral scent trails. Atmos. Environ. 2008, 42, 2336–2348. [Google Scholar] [CrossRef]
- Fuentes, J.D.; Roulston, T.H.; Zenker, J. Ozone impedes the ability of a herbivore to find its host. Environ. Res. Lett. 2013, 8, 014048. [Google Scholar] [CrossRef]
- Koike, T. Physiological ecology of the growth characteristics of Japanese mountain birch in Northern Japan: A comparison with Japanese white birch. In Vegetation Sciences in Forestry: Global Perspective Based on Forest Ecosystems of East and Southeast Asia (Handbook of Vegetation Science); Box, E.O., Peet, R.K., Masuzawa, T., Yamada, I., Fujiwara, K., Maycock, P.F., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; Volume 12, pp. 409–422. [Google Scholar]
- Park, I.K.; Lee, S.G.; Shin, S.C.; Kim, C.S.; Ahn, Y.J. Feeding and attraction of Agelastica coerulea. (Coleoptera: Chrysomelidae) to Betulaceae plants. J. Econ. Entomol. 2004, 97, 1978–1982. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.H.; Han, K.S.; Boo, K.S. Identification of Volatiles Responsive to the Alder Leaf Beetle (Agelastica coerulea) from its Host Plant, the Japanese Alder (Alnus glutinosa). J. Asia Pac. Entomol. 2000, 3, 33–40. [Google Scholar] [CrossRef]
- Tani, A.; Kawawata, Y. Isoprene emission from the major native Quercus spp. in Japan. Atmos. Environ. 2008, 42, 4540–4550. [Google Scholar] [CrossRef]
- Shimoda, T.; Ozawa, R.; Arimura, G.; Takabayashi, J.; Nishioka, T. Olfactory responses of two specialist insect predators of spider mites toward plant volatiles from lima bean leaves induced by jasmonic acid and/or methyl salicylate. Appl. Entomol. Zool. 2002, 37, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Brilli, F.; Ciccioli, P.; Frattoni, M.; Prestininzi, M.; Spanedda, A.F.; Loreto, F. Constitutive and herbivore-induced monoterpenes emitted by Populus × euroamericana leaves are key volatiles that orient Chrysomela populi beetles. Plant. Cell. Environ. 2009, 32, 542–552. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends. Plant. Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef]
- Reissell, A.; Aschmann, S.M.; Atkinson, R.; Arey, J. Products of the OH radical- and O3-initiated reactions of myrcene and ocimene. J. Geophys. Res. 2002, 107. [Google Scholar] [CrossRef]
- Agathokleous, E. Environmental hormesis, a fundamental non-monotonic biological phenomenon with implications in ecotoxicology and environmental safety. Ecotox. Environ. Saf. 2018, 148, 1042–1053. [Google Scholar] [CrossRef] [Green Version]
- Copolovici, L.; Kännaste, A.; Remmel, T.; Vislap, V.; Niinemets, Ü. Volatile emissions from Alnus glutionosa induced by herbivory are quantitatively related to the extent of damage. J. Chem. Ecol. 2011, 37, 18–28. [Google Scholar] [CrossRef]
- Kessler, A.; Baldwin, I.T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef] [PubMed]
- De Moraes, C.M.; Lewis, W.J.; Paré, P.W.; Alborn, H.T.; Tumlinson, J.H. Herbivore-infested plants selectively attract parasitoids. Nature 1998, 393, 570–573. [Google Scholar] [CrossRef]
- Bolter, C.J.; Dicke, M.; Van Loon, J.J.A.; Visser, J.H.; Posthumus, M.A. Attraction of Colorado potato beetle to herbivore-damaged plants during herbivory and after its termination. J. Chem. Ecol. 1997, 23, 1003–1023. [Google Scholar] [CrossRef]
- Dicke, M. Behavioural and community ecology of plants that cry for help. Plant. Cell. Environ. 2009, 32, 654–665. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| k2 | [O3] 28.4 ppb | [O3] 80.0 ppb | |||
|---|---|---|---|---|---|
| Monoterpenes | |||||
| Camphene | 9.2 × 10−19 | 18.0 | d | 6.4 | d |
| 2-Carene | 2.3 × 10−16 | 1.7 | h | 36.2 | m |
| 3-Carene | 3.6 × 10−17 | 11.0 | h | 3.9 | h |
| Limonene | 2.0 × 10−16 | 2.0 | h | 42.6 | m |
| Myrcene | 4.8 × 10−16 | 50.0 | m | 17.7 | m |
| cis-/trans-Ocimene | 5.4 × 10−16 | 44.0 | m | 15.7 | m |
| α-Phellandrene | 3.0 × 10−15 | 8.0 | m | 2.8 | m |
| β-Phellandrene | 4.7 × 10−17 | 8.4 | h | 3.0 | h |
| α-Pinene | 8.6 × 10−17 | 4.6 | h | 1.6 | h |
| β-Pinene | 1.5 × 10−17 | 1.1 | d | 0.4 | d |
| Sabinene | 8.3 × 10−17 | 4.8 | h | 1.7 | h |
| α-Terpinene | 2.4 × 10−14 | 1.0 | m | 0.4 | m |
| γ-Terpinene | 1.4 × 10−16 | 2.8 | h | 59.6 | m |
| Terpinolene | 1.8 × 10−15 | 13.0 | m | 4.6 | m |
| Sesquiterpenes | |||||
| β-Caryophyllene | 1.2 × 10−14 | 2.0 | m | 0.7 | m |
| α-Cedrene | 2.8 × 10−17 | 14.0 | h | 5.0 | h |
| α-Copaene | 1.6 × 10−16 | 2.5 | h | 53.2 | m |
| α-Humulene | 1.2 × 10−14 | 2.0 | m | 0.7 | m |
| Oxygenates | |||||
| 1,8-Cineole | - | >110 | d | - | |
| cis-3-Hexen-1-ol | 6.4 × 10−17 | 6.2 | h | 2.2 | h |
| cis-3-Hexenyl acetate | 5.4 × 10−17 | 7.3 | h | 2.6 | h |
| Linalool | 4.3 × 10−16 | 55.0 | m | 19.7 | m |
| 2-Methyl-3-buten-2-ol | 9.7 × 10−18 | 1.7 | d | 0.6 | d |
| 6-Methyl-5-hepten-2-one | 4.0 × 10−16 | 1.0 | h | 21.3 | m |
| Group | Emission Rate (%) | F-Value | T-test | ||
|---|---|---|---|---|---|
| Early Leaves | Late Leaves | ||||
| June | September | Type | p-Value | ||
| MT (monoterpene) | 65.252 ± 2.637 | 96.569 ± 0.272 | 93.79 | W | ** |
| Oxygenated-MT | 17.119 ± 2.028 | 2.366 ± 0.092 | 481.95 | W | ** |
| SQT (sesquiterpene) | 7.971 ± 3.955 | 0.217 ± 0.050 | 6186.57 | W | n.s |
| N-compound | 3.272 ± 0.558 | 0.173 ± 0.045 | 156.94 | W | ** |
| GLV (green leaf volatile) | 6.386 ± 0.978 | 0.613 ± 0.245 | 15.97 | W | ** |
| Organic acid-ester | - | 0.062 ± 0.024 | - | - | - |
| June | September | F Value | T-test | ||||
|---|---|---|---|---|---|---|---|
| Number of Compounds | 30 | 27 | Type | p Value | |||
| Total Emission rate (μmol m−2s−1) | 0.983 ± 0.182 | 2.115 ± 0.134 | 1.85 | S | ** | ||
| Name of compounds | RT | Group | |||||
| hexanal | 16.53 | GLV | 0.724 ± 0.195 | 0.151 ± 0.044 | 20.10 | W | * |
| cis-3-hexen-1-ol | 18.82 | GLV | 1.059 ± 0.111 | 0.462 ± 0.211 | 3.64 | S | * |
| α-thujene | 21.72 | MT | 1.485 ± 0.133 | 1.566 ± 0.188 | 1.99 | S | n.s |
| α-pinene | 22.13 | MT | 7.495 ± 0.259 | 7.786 ± 0.229 | 1.27 | S | n.s |
| camphene | 22.84 | MT | 0.243 ± 0.027 | 0.297 ± 0.032 | 1.40 | S | n.s |
| sabinene | 23.56 | MT | 40.980 ± 2.212 | 50.107 ± 1.804 | 1.50 | S | * |
| β-pinene | 23.88 | MT | 3.299 ± 0.088 | 3.459 ± 0.143 | 2.64 | S | n.s |
| trans-3-hexenyl acetate | 24.32 | GLV | 4.009 ± 0.831 | — | — | — | — |
| cis-2-hexenyl acetate | 24.54 | GLV | 0.594 ± 0.134 | — | — | — | — |
| 2-carene | 25.13 | MT | 0.600 ± 0.081 | 0.610 ± 0.125 | 2.41 | S | n.s |
| o-cymene | 25.39 | MT | 1.789 ± 0.635 | 1.623 ± 0.440 | 2.08 | S | n.s |
| trans-β-ocimene | 25.48 | MT | 0.992 ± 0.174 | 4.334 ± 0.325 | 3.47 | S | ** |
| limonene | 25.58 | MT | 0.454 ± 0.059 | 0.669 ± 0.041 | 2.08 | S | * |
| β-phellandrene | 25.71 | MT | 1.006 ± 0.098 | 1.022 ± 0.144 | 2.16 | S | n.s |
| p-Cineole | 25.78 | Oxy-MT | 1.170 ± 0.441 | 0.238 ± 0.011 | 1552.13 | W | n.s |
| cis-β-ocimene | 25.89 | MT | 4.249 ± 0.797 | 17.920 ± 1.349 | 2.87 | S | ** |
| γ-terpinene | 26.53 | MT | 1.474 ± 0.203 | 1.467 ± 0.322 | 2.52 | S | n.s |
| trans-sabinene-hydrate | 27.01 | Oxy-MT | 0.911 ± 0.117 | 1.437 ± 0.049 | 5.60 | S | ** |
| linalool oxide | 27.48 | Oxy-MT | 4.706 ± 0.770 | 0.510 ± 0.044 | 307.73 | W | ** |
| β-linalool | 27.71 | Oxy-MT | 10.333 ± 1.929 | 0.181 ± 0.036 | 2829.78 | W | ** |
| geranyl nitrile | 28.11 | N | 3.272 ± 0.558 | 0.173 ± 0.045 | 156.94 | W | ** |
| neo-allo-ocimene | 28.65 | MT | 1.185 ± 0.215 | 5.709 ± 0.426 | 3.93 | S | ** |
| methyl salicylate | 31.14 | Oa-est | — | 0.062 ± 0.024 | — | — | — |
| ylangene | 36.40 | SQT | 0.900 ± 0.427 | 0.035 ± 0.010 | 1666.55 | W | n.s |
| α-copaene | 36.57 | SQT | 0.731 ± 0.336 | 0.022 ± 0.006 | 3104.84 | W | n.s |
| β-bourbonene | 36.88 | SQT | 1.470 ± 0.622 | 0.074 ± 0.016 | 1451.45 | W | n.s |
| β-caryophyllene | 37.93 | SQT | 0.743 ± 0.317 | 0.034 ± 0.008 | 1409.37 | W | n.s |
| β-copaene | 38.13 | SQT | 0.547 ± 0.276 | — | — | — | — |
| aristolene | 38.29 | SQT | 2.461 ± 1.448 | — | — | — | — |
| α-farnesene | 39.35 | SQT | — | 0.030 ± 0.005 | — | — | — |
| germacrene | 39.48 | SQT | 0.807 ± 0.437 | — | — | — | — |
| α-guaiene | 39.73 | SQT | 0.312 ± 0.138 | 0.023 ± 0.006 | 482.87 | W | n.s |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masui, N.; Mochizuki, T.; Tani, A.; Matsuura, H.; Agathokleous, E.; Watanabe, T.; Koike, T. Does Ozone Alter the Attractiveness of Japanese White Birch Leaves to the Leaf Beetle Agelastica coerulea via Changes in Biogenic Volatile Organic Compounds (BVOCs): An Examination with the Y-Tube Test. Forests 2020, 11, 58. https://doi.org/10.3390/f11010058
Masui N, Mochizuki T, Tani A, Matsuura H, Agathokleous E, Watanabe T, Koike T. Does Ozone Alter the Attractiveness of Japanese White Birch Leaves to the Leaf Beetle Agelastica coerulea via Changes in Biogenic Volatile Organic Compounds (BVOCs): An Examination with the Y-Tube Test. Forests. 2020; 11(1):58. https://doi.org/10.3390/f11010058
Chicago/Turabian StyleMasui, Noboru, Tomoki Mochizuki, Akira Tani, Hideyuki Matsuura, Evgenios Agathokleous, Toshihiro Watanabe, and Takayoshi Koike. 2020. "Does Ozone Alter the Attractiveness of Japanese White Birch Leaves to the Leaf Beetle Agelastica coerulea via Changes in Biogenic Volatile Organic Compounds (BVOCs): An Examination with the Y-Tube Test" Forests 11, no. 1: 58. https://doi.org/10.3390/f11010058
APA StyleMasui, N., Mochizuki, T., Tani, A., Matsuura, H., Agathokleous, E., Watanabe, T., & Koike, T. (2020). Does Ozone Alter the Attractiveness of Japanese White Birch Leaves to the Leaf Beetle Agelastica coerulea via Changes in Biogenic Volatile Organic Compounds (BVOCs): An Examination with the Y-Tube Test. Forests, 11(1), 58. https://doi.org/10.3390/f11010058