Warming Effects on Topsoil Organic Carbon and C:N:P Stoichiometry in a Subtropical Forested Landscape


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sampling Design and Plant Census
2.3. Soil Sampling and Laboratory Determination
2.4. Understory Radiation Measurements
2.5. Potential Direct Incident Radiation
2.6. Warming Gradient and Warming Index
2.7. Statistical Analyses
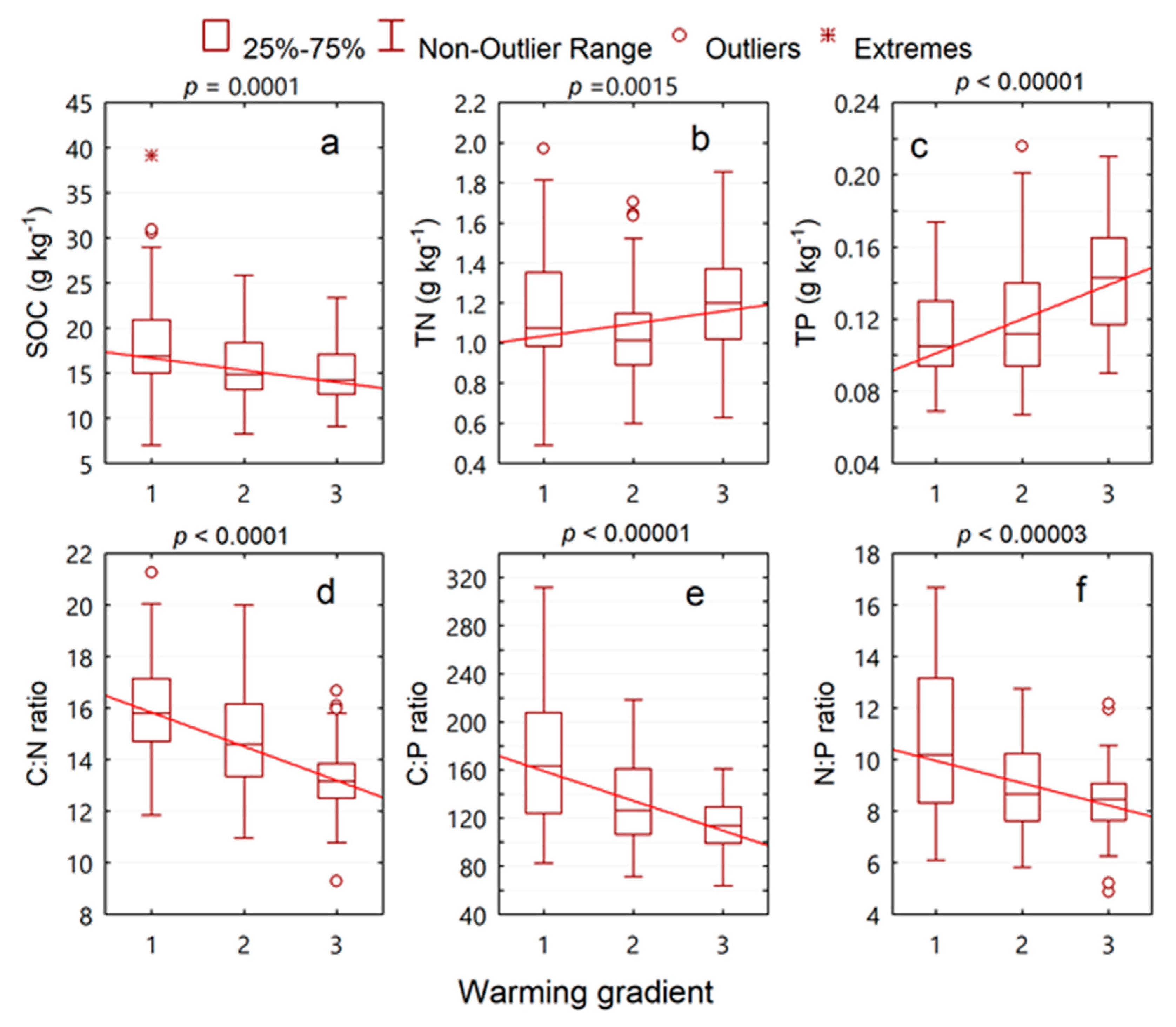
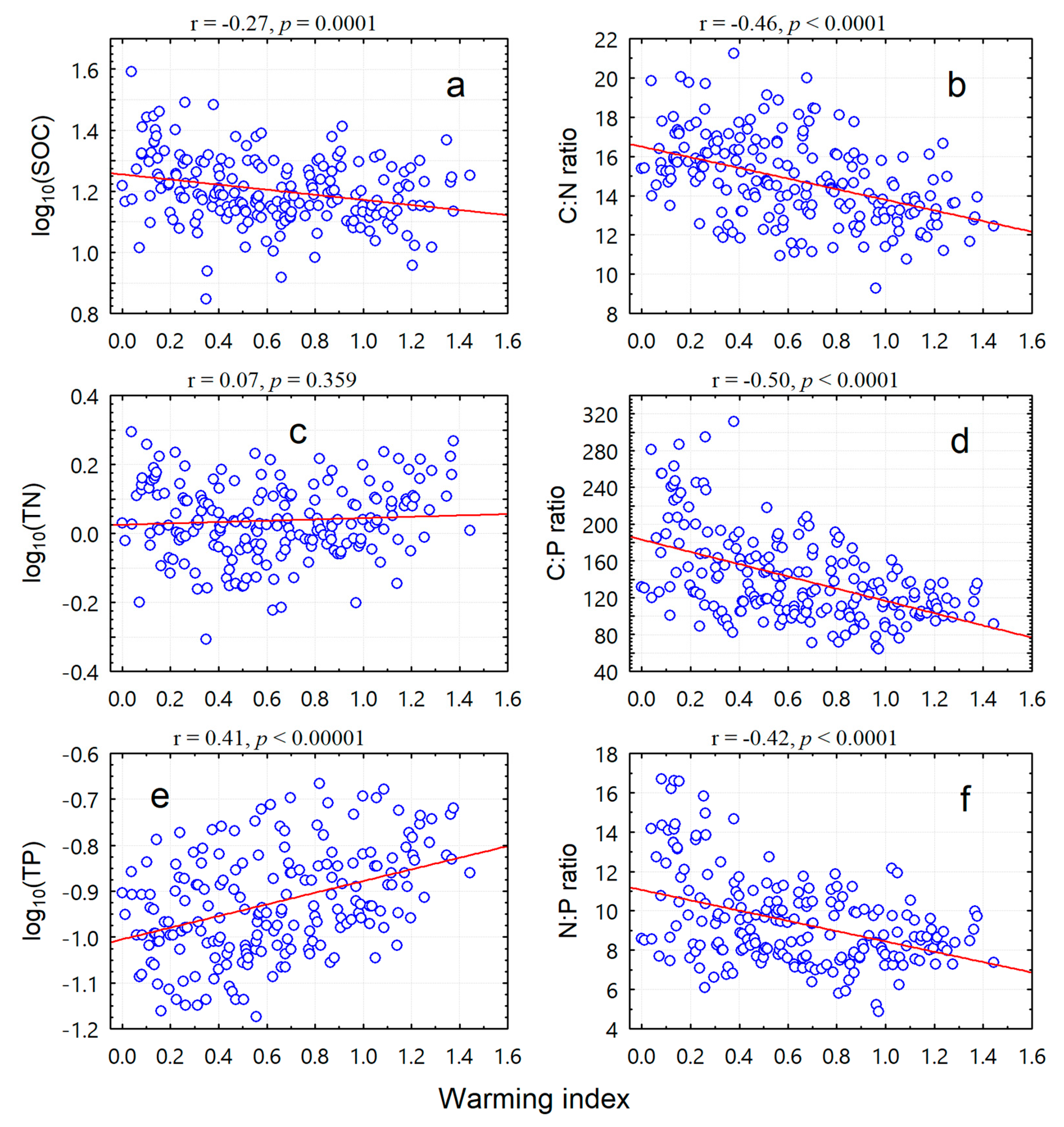
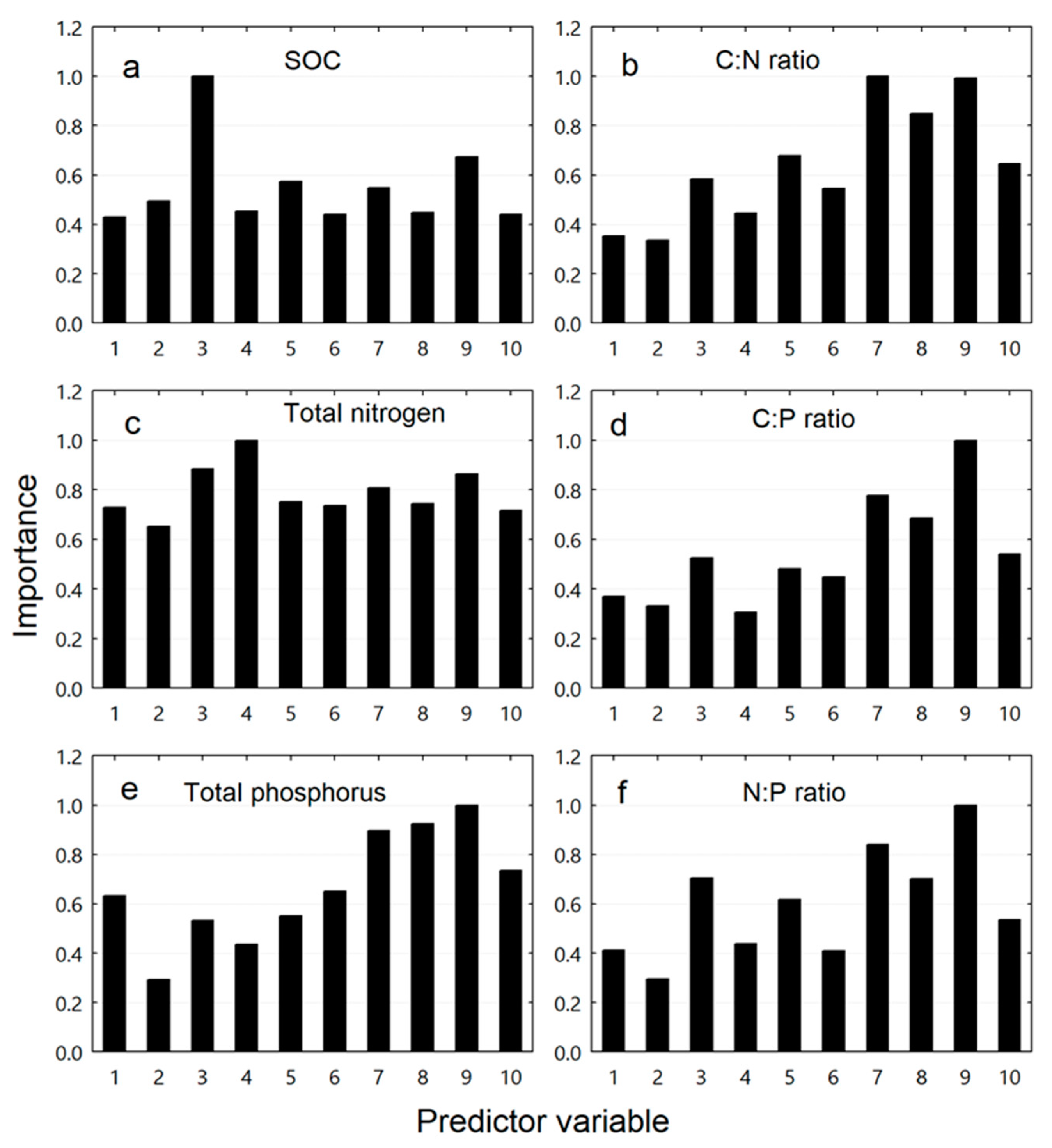
3. Results
3.1. Spatial Variability
3.2. Inverse Elevation-Dependent Warming Effects
3.3. Partial Correlations
3.4. Relative Importance of Predictor Variables
4. Discussion
4.1. Warming Effects on SOC and C:N:P Stoichiometry
4.2. Warming Effects on the Relationships of SOC and C:N:P Stoichiometry to Habitat Factors
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: New York, NY, USA, 2013. [Google Scholar]
- Cheesman, A.W.; Winter, K. Elevated night-time temperatures increase growth in seedlings of two tropical pioneer tree species. New Phytol. 2013, 197, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.H.; Fu, Y.L.; Zhou, L.Y.; Li, B.; Luo, Y.Q. An imperative need for global change research in tropical forests. Tree Physiol. 2013, 33, 903–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IPCC. Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson-Delmotte, V., Zhai, P., Pörtner, H.-O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2018. [Google Scholar]
- Klutse, N.A.B.; Ajayi, V.O.; Gbobaniyi, E.O.; Egbebiyi, T.S.; Kouadio, K.; Nkrumah, F.; Quagraine, K.A.; Olusegun, C.; Diasso, U.; Abiodun, B.J.; et al. Potential impact of 1.5 degrees C and 2 degrees C global warming on consecutive dry and wet days over West Africa. Environ. Res. Lett. 2018, 13, 055013. [Google Scholar] [CrossRef]
- Mba, W.P.; Longandjo, G.N.T.; Moufouma-Okia, W.; Bell, J.P.; James, R.; Vondou, D.A.; Haensler, A.; Fotso-Nguemo, T.C.; Guenang, G.M.; Tchotchou, A.L.D.; et al. Consequences of 1.5 degrees C and 2 degrees C global warming levels for temperature and precipitation changes over Central Africa. Environ. Res. Lett. 2018, 13, 055011. [Google Scholar] [CrossRef] [Green Version]
- Nikulin, G.; Lennard, C.; Dosio, A.; Kjellstrom, E.; Chen, Y.M.; Hansler, A.; Kupiainen, M.; Laprise, R.; Mariotti, L.; Maule, C.F.; et al. The effects of 1.5 and 2 degrees of global warming on Africa in the CORDEX ensemble. Environ. Res. Lett. 2018, 13, 065003. [Google Scholar] [CrossRef]
- Li, C.L.; Cao, Z.Y.; Chang, J.J.; Zhang, Y.; Zhu, G.L.; Zong, N.; He, Y.T.; Zhang, J.J.; He, N.P. Elevational gradient affect functional fractions of soil organic carbon and aggregates stability in a Tibetan alpine meadow. Catena 2017, 156, 139–148. [Google Scholar] [CrossRef]
- Sierra, J.; Causeret, F. Changes in soil carbon inputs and outputs along a tropical altitudinal gradient of volcanic soils under intensive agriculture. Geoderma 2018, 320, 95–104. [Google Scholar] [CrossRef]
- Werner, C.; Schmidt, H.P.; Gerten, D.; Lucht, W.; Kammann, C. Biogeochemical potential of biomass pyrolysis systems for limiting global warming to 1.5 degrees C. Environ. Res. Lett. 2018, 13, 044036. [Google Scholar] [CrossRef]
- Scharlemann, J.P.; Tanner, E.V.; Hiederer, R.; Kapos, V.J.C.M. Global soil carbon: Understanding and managing the largest terrestrial carbon pool. Carbon Manag. 2014, 5, 81–91. [Google Scholar] [CrossRef]
- Bazzaz, F.A. Tropical forests in a future climate: Changes in biological diversity and impact on the global carbon cycle. Clim. Chang. 1998, 39, 317–336. [Google Scholar] [CrossRef]
- Miles, L.; Grainger, A.; Phillips, O. The impact of global climate change on tropical forest biodiversity in Amazonia. Glob. Ecol. Biogeogr. 2004, 13, 553–565. [Google Scholar] [CrossRef]
- Billings, S.A.; Ballantyne, F. How interactions between microbial resource demands, soil organic matter stoichiometry, and substrate reactivity determine the direction and magnitude of soil respiratory responses to warming. Glob. Chang. Biol. 2013, 19, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Kirkby, C.A.; Richardson, A.E.; Wade, L.J.; BattenB, G.D.; Blanchard, C.; Kirkegaard, J.A. Carbon-nutrient stoichiometry to increase soil carbon sequestration. Soil Biol. Biochem. 2013, 60, 77–86. [Google Scholar] [CrossRef]
- Bojko, O.; Kabala, C. Organic carbon pools in mountain soils—Sources of variability and predicted changes in relation to climate and land use changes. Catena 2017, 149, 209–220. [Google Scholar] [CrossRef]
- Chen, D.F.; Yu, M.; Gonzalez, G.; Zou, X.M.; Gao, Q. Climate Impacts on soil carbon processes along an elevation gradient in the tropical Luquillo Experimental Forest. Forests 2017, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Simon, A.; Dhendup, K.; Rai, P.B.; Gratzer, G. Soil carbon stocks along elevational gradients in Eastern Himalayan mountain forests. Geoderma Reg. 2018, 12, 28–38. [Google Scholar] [CrossRef]
- Hu, C.; Li, F.; Xie, Y.H.; Deng, Z.M.; Chen, X.S. Soil carbon, nitrogen, and phosphorus stoichiometry of three dominant plant communities distributed along a small-;scale elevation gradient in the East Dongting Lake. Phys. Chem. Earth 2018, 103, 28–34. [Google Scholar] [CrossRef]
- Zhang, K.; Su, Y.Z.; Yang, R. Variation of soil organic carbon, nitrogen, and phosphorus stoichiometry and biogeographic factors across the desert ecosystem of Hexi Corridor, northwestern China. J. Soil Sediment 2019, 19, 49–57. [Google Scholar] [CrossRef]
- Hansen, K.; Butzeck, C.; Eschenbach, A.; Grongroft, A.; Jensen, K.; Pfeiffer, E.M. Factors influencing the organic carbon pools in tidal marsh soils of the Elbe estuary (Germany). J. Soil Sediment 2017, 17, 47–60. [Google Scholar] [CrossRef]
- Singh, P.; Bengtsson, L. Hydrological sensitivity of a large Himalayan basin to climate change. Hydrol. Process. 2004, 18, 2363–2385. [Google Scholar] [CrossRef]
- Hu, Y.; Su, Z.; Li, W.; Li, J.; Ke, X.J. Influence of tree species composition and community structure on carbon density in a subtropical forest. PLoS ONE 2015, 10, e0136984. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Zhong, Y.; Sun, Y.; Su, Z.; Jia, X.; Hu, Y.; Zhou, Q. Topography-associated thermal gradient predicts warming effects on woody plant structural diversity in a subtropical forest. Sci. Rep. 2017, 7, 40387. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.; Mi, X.; Ren, H.; Ma, K.J. Species-habitat associations change in a subtropical forest of China. J. Veg. Sci. 2009, 20, 415–423. [Google Scholar] [CrossRef]
- Condit, R. Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots; Springer: Berlin, Germany, 1998. [Google Scholar]
- Bao, S. Soil and Agricultural Chemistry Analysis; China Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Frazer, G.W.; Canham, C.D.; Lertzman, K.P. Gap Light Analyzer (GLA), Version 2.0: Imaging Software to Extract Canopy Structure and Gap Light Transmission Indices from True-Colour Fisheye Photographs, Users Manual and Program Documentation; Simon Fraser University: Burnaby, BC, Canada; The Institute of Ecosystem Studies: Millbrook, NY, USA, 1999. [Google Scholar]
- McCune, B. Improved estimates of incident radiation and heat load using non-parametric regression against topographic variables. J. Veg. Sci. 2007, 18, 751–754. [Google Scholar] [CrossRef]
- Santos, R.M.; Oliveira, A.T.; Eisenlohr, P.V.; Queiroz, L.P.; Cardoso, D.B.O.S.; Rodal, M.J.N. Identity and relationships of the Arboreal Caatinga among other floristic units of seasonally dry tropical forests (SDTFs) of north-eastern and Central Brazil. Ecol. Evol. 2012, 2, 409–428. [Google Scholar] [CrossRef]
- Zhou, Y.; Su, Y.; Zhong, Y.; Xie, P.; Xu, M.; Su, Z. Community attributes predict the relationship between habitat invasibility and land use types in an agricultural and forest landscape. Forests 2019, 10, 867. [Google Scholar] [CrossRef] [Green Version]
- Breiman, L. Random forest. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Breiman, L.; Last, M.; Rice, J. Random forests: Finding quasars. In Statistical Challenges in Astronomy; Springer: New York, NY, USA, 2003; pp. 243–254. [Google Scholar]
- Sun, X. Soil Science; China Forestry Press: Beijing, China, 2008. [Google Scholar]
- Fang, Y.; Mo, J.; Sandra, B.; Zhou, G.; Zhang, Q.; Li, D.J. Storage and distribution of soil organic carbon in Dinghushan Biosphere Reserve. Acta Ecol. Sin. 2004, 24, 135–142. [Google Scholar]
- Rial, M.; Cortizas, A.M.; Rodriguez-Lado, L. Understanding the spatial distribution of factors controlling topsoil organic carbon content in European soils. Sci. Total. Environ. 2017, 609, 1411–1422. [Google Scholar] [CrossRef]
- Song, G.H.; Li, L.Q.; Pan, G.X.; Zhang, Q. Topsoil organic carbon storage of China and its loss by cultivation. Biogeochemistry 2005, 74, 47–62. [Google Scholar] [CrossRef]
- Boddy, E.; Hill, P.W.; Farrar, J.; Jones, D.L. Fast turnover of low molecular weight components of the dissolved organic carbon pool of temperate grassland field soils. Soil Biol. Biochem. 2007, 39, 827–835. [Google Scholar] [CrossRef]
- Zhang, Z.; Cai, X.; Tang, C.; Guo, J. Priming effect of dissolved organic matter in the surface soil of a Cunninghamia lanceolata plantation. Acta Ecol. Sin. 2017, 37, 7660–7667. [Google Scholar]
- Jobbagy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Chen, C.R.; Xu, Z.H.; Zhang, S.L.; Keay, P. Soluble organic nitrogen pools in forest soils of subtropical Australia. Plant Soil 2005, 277, 285–297. [Google Scholar] [CrossRef]
- Alberti, G.; Vicca, S.; Inglima, I.; Belelli-Marchesini, L.; Genesio, L.; Miglietta, F.; Marjanovic, H.; Martinez, C.; Matteucci, G.; D’Andrea, E.; et al. Soil C:N stoichiometry controls carbon sink partitioning between above-ground tree biomass and soil organic matter in high fertility forests. Iforest 2015, 8, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Li, D.J.; Cheng, X.L.; Ruan, H.H.; Luo, Y.Q. Carbon: Nitrogen stoichiometry following afforestation: A global synthesis. Sci. Rep. 2016, 6, 19117. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Chen, Y.M. Ecosystem C:N:P stoichiometry and carbon storage in plantations and a secondary forest on the Loess Plateau, China. Ecol. Eng. 2017, 105, 125–132. [Google Scholar] [CrossRef]
- Zhao, F.Z.; Kang, D.; Han, X.H.; Yang, G.H.; Yang, G.H.; Feng, Y.Z.; Ren, G.X. Soil stoichiometry and carbon storage in long-term afforestation soil affected by understory vegetation diversity. Ecol. Eng. 2015, 74, 415–422. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, G.X.; Shen, H.H.; Yang, Y.; Cui, H.J.; Liu, Q. Dynamics of carbon and nitrogen accumulation and C:N stoichiometry in a deciduous broadleaf forest of deglaciated terrain in the eastern Tibetan Plateau. For. Ecol. Manag. 2014, 312, 10–18. [Google Scholar] [CrossRef]
- Gutierrez-Giron, A.; Diaz-Pines, E.; Rubio, A.; Gavilan, R.G. Both altitude and vegetation affect temperature sensitivity of soil organic matter decomposition in Mediterranean high mountain soils. Geoderma 2015, 237, 1–8. [Google Scholar] [CrossRef]
- Fissore, C.; Giardina, C.P.; Kolka, R.K.; Trettin, C.C.; King, G.M.; Jurgensen, M.F.; Barton, C.D.; Mcdowell, S.D. Temperature and vegetation effects on soil organic carbon quality along a forested mean annual temperature gradient in North America. Glob. Chang. Biol. 2008, 14, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.M.; Johnston, E.R.; Li, L.H.; Konstantinidis, K.T.; Han, X.G. Experimental warming reveals positive feedbacks to climate change in the Eurasian Steppe. ISME J. 2017, 11, 885–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, F.; You, Q.G.; Xu, M.H.; Zhou, X.H.; Wang, T.; Guo, J.; Xue, X. Effects of experimental warming on soil respiration and its components in an alpine meadow in the permafrost region of the Qinghai-Tibet Plateau. Eur. J. Soil. Sci. 2015, 66, 145–154. [Google Scholar] [CrossRef]
- Schindlbacher, A.; Rodler, A.; Kuffner, M.; Kitzler, B.; Sessitsch, A.; Zechmeister-Boltenstern, S. Experimental warming effects on the microbial community of a temperate mountain forest soil. Soil Biol. Biochem. 2011, 43, 1417–1425. [Google Scholar] [CrossRef] [Green Version]
- Mau, R.L.; Dijkstra, P.; Schwartz, E.; Koch, B.J.; Hungate, B.A. Warming induced changes in soil carbon and nitrogen influence priming responses in four ecosystems. Appl. Soil. Ecol. 2018, 124, 110–116. [Google Scholar] [CrossRef]
- Li, Y.Q.; Qing, Y.X.; Lyu, M.K.; Chen, S.D.; Yang, Z.J.; Lin, C.F.; Yang, Y.S. Effects of artificial warming on different soil organic carbon and nitrogen pools in a subtropical plantation. Soil Biol. Biochem. 2018, 124, 161–167. [Google Scholar] [CrossRef]
- Karhu, K.; Auffret, M.D.; Dungait, J.A.; Hopkins, D.W.; Prosser, J.I.; Singh, B.K.; Subke, J.A.; Wookey, P.A.; Agren, G.I.; Sebastia, M.T.; et al. Temperature sensitivity of soil respiration rates enhanced by microbial community response. Nature 2014, 513, 81–84. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef]
- Bradford, M.A.; Davies, C.A.; Frey, S.D.; Maddox, T.R.; Melillo, J.M.; Mohan, J.E.; Reynolds, J.F.; Treseder, K.K.; Wallenstein, M.D. Thermal adaptation of soil microbial respiration to elevated temperature. Ecol. Lett. 2008, 11, 1316–1327. [Google Scholar] [CrossRef]
- Rustad, L.; Campbell, J.; Marion, G.; Norby, R.; Mitchell, M.; Hartley, A.; Cornelissen, J.; Gurevitch, J.; Gcte, N. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 2001, 126, 543–562. [Google Scholar] [CrossRef]
- Wang, N.; Fu, F.Z.; Wang, B.T.; Wang, R.J. Carbon, nitrogen and phosphorus stoichiometry in Pinus tabulaeformis forest ecosystems in warm temperate Shanxi Province, north China. J. For. Res. 2018, 29, 1665–1673. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Attribute | Mean | Median | Minimum | Maximum | Standard Deviation | Coefficient of Variation (%) |
|---|---|---|---|---|---|---|
| SOC (g kg−1) | 16.55 | 15.46 | 7.04 | 39.12 | 4.55 | 27.50 |
| TN (g kg−1) | 1.12 | 1.07 | 0.49 | 1.97 | 0.28 | 24.58 |
| TP (g kg−1) | 0.12 | 0.12 | 0.07 | 0.22 | 0.03 | 26.71 |
| C:N ratio | 14.82 | 14.67 | 9.29 | 21.25 | 2.15 | 14.48 |
| C:P ratio | 141.99 | 129.12 | 64.16 | 311.66 | 48.10 | 33.88 |
| N:P ratio | 9.44 | 8.80 | 4.88 | 16.69 | 2.28 | 24.15 |
| Response Variable | Variables to Be Controlled for | ||
|---|---|---|---|
| PDIR Only | TransTot Only | PDIR and TransTot | |
| SOC | −0.27 * | −0.26 * | −0.26 * |
| TN | 0.07 | 0.06 | 0.07 |
| TP | 0.44 ** | 0.40 ** | 0.44 ** |
| C:N ratio | −0.49 ** | −0.45 ** | −0.48 ** |
| C:P ratio | −0.54 ** | −0.50 ** | −0.54 ** |
| N:P ratio | −0.45 ** | −0.42 ** | −0.44 ** |
| Predictor Variable | Type of Coefficient | Response Variable | |||||
|---|---|---|---|---|---|---|---|
| SOC | TN | TP | C:N Ratio | C:P Ratio | N:P Ratio | ||
| PDIR | r | 0.02a | −0.09a | −0.26 **a | 0.21 *a | 0.23 *a | 0.20 *a |
| partial r | 0.04a | −0.10a | −0.32 **a | 0.28 **a | 0.32 **a | 0.26 **a | |
| TransTot | r | 0.10a | −0.05a | −0.07a | 0.18 *a | 0.10a | 0.03a |
| partial r | 0.07a | −0.04a | −0.03a | 0.16 *a | 0.06a | −0.01a | |
| Nunder | r | −0.07a | −0.37 ***a | −0.20 *a | 0.20 *a | −0.07a | −0.19 *a |
| partial r | −0.14a | −0.36 ***a | −0.12a | 0.12a | −0.22 *a | −0.32 **a | |
| Sunder | r | −0.10a | 0.05a | 0.33 **a | −0.36 ***a | −0.41 ***a | −0.34 **a |
| partial r | 0.04a | 0.02a | 0.17 *b | −0.17 *b | −0.21 *b | −0.17 *b | |
| H’under | r | −0.07a | 0.24 **a | 0.37 **a | −0.41 ***a | −0.29 **a | −0.17 *a |
| partial r | 0.12a | 0.25 **a | 0.19 *b | −0.19 *b | 0.01b | 0.10b | |
| Axis1under | r | −0.04a | 0.16 *a | 0.32 **a | −0.29 **a | −0.26 **a | −0.19 *a |
| partial r | 0.064a | 0.146 *a | 0.203 *a | −0.159 *a | −0.104b | −0.053a | |
| Axis2under | r | 0.13a | −0.18 *a | −0.44 ***a | 0.45 ***a | 0.39 ***a | 0.30 **a |
| partial r | 0.06a | −0.16 *a | −0.37 ***a | 0.37 ***a | 0.29 **a | 0.20 *a | |
| Axis1tree | r | −0.23 *a | 0.22 *a | 0.54 ***a | −0.57 ***a | −0.50 ***a | −0.38 ***a |
| partial r | −0.10a | 0.22 *a | 0.41 ***b | −0.43 ***b | −0.30 **b | −0.19 *b | |
| Axis2tree | r | 0.08a | 0.11a | 0.03a | 0.12a | 0.09a | 0.07a |
| partial r | 0.14 *a | 0.10a | −0.07a | 0.25 **a | 0.24 **a | 0.19 *a | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Y.; Wu, Z.; Xie, P.; Zhang, L.; Chen, H. Warming Effects on Topsoil Organic Carbon and C:N:P Stoichiometry in a Subtropical Forested Landscape. Forests 2020, 11, 66. https://doi.org/10.3390/f11010066
Su Y, Wu Z, Xie P, Zhang L, Chen H. Warming Effects on Topsoil Organic Carbon and C:N:P Stoichiometry in a Subtropical Forested Landscape. Forests. 2020; 11(1):66. https://doi.org/10.3390/f11010066
Chicago/Turabian StyleSu, Yuqiao, Zhuoling Wu, Peiyun Xie, Lu Zhang, and Hui Chen. 2020. "Warming Effects on Topsoil Organic Carbon and C:N:P Stoichiometry in a Subtropical Forested Landscape" Forests 11, no. 1: 66. https://doi.org/10.3390/f11010066
APA StyleSu, Y., Wu, Z., Xie, P., Zhang, L., & Chen, H. (2020). Warming Effects on Topsoil Organic Carbon and C:N:P Stoichiometry in a Subtropical Forested Landscape. Forests, 11(1), 66. https://doi.org/10.3390/f11010066