Flux-Based Ozone Risk Assessment for a Plant Injury Index (PII) in Three European Cool-Temperate Deciduous Tree Species

 ,
,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Plant Material
2.2. Assessment of Ozone Visible Injury
2.3. Modeling of Stomatal Conductance
2.4. Calculation of Ozone Indices
2.5. Data Analysis
3. Results
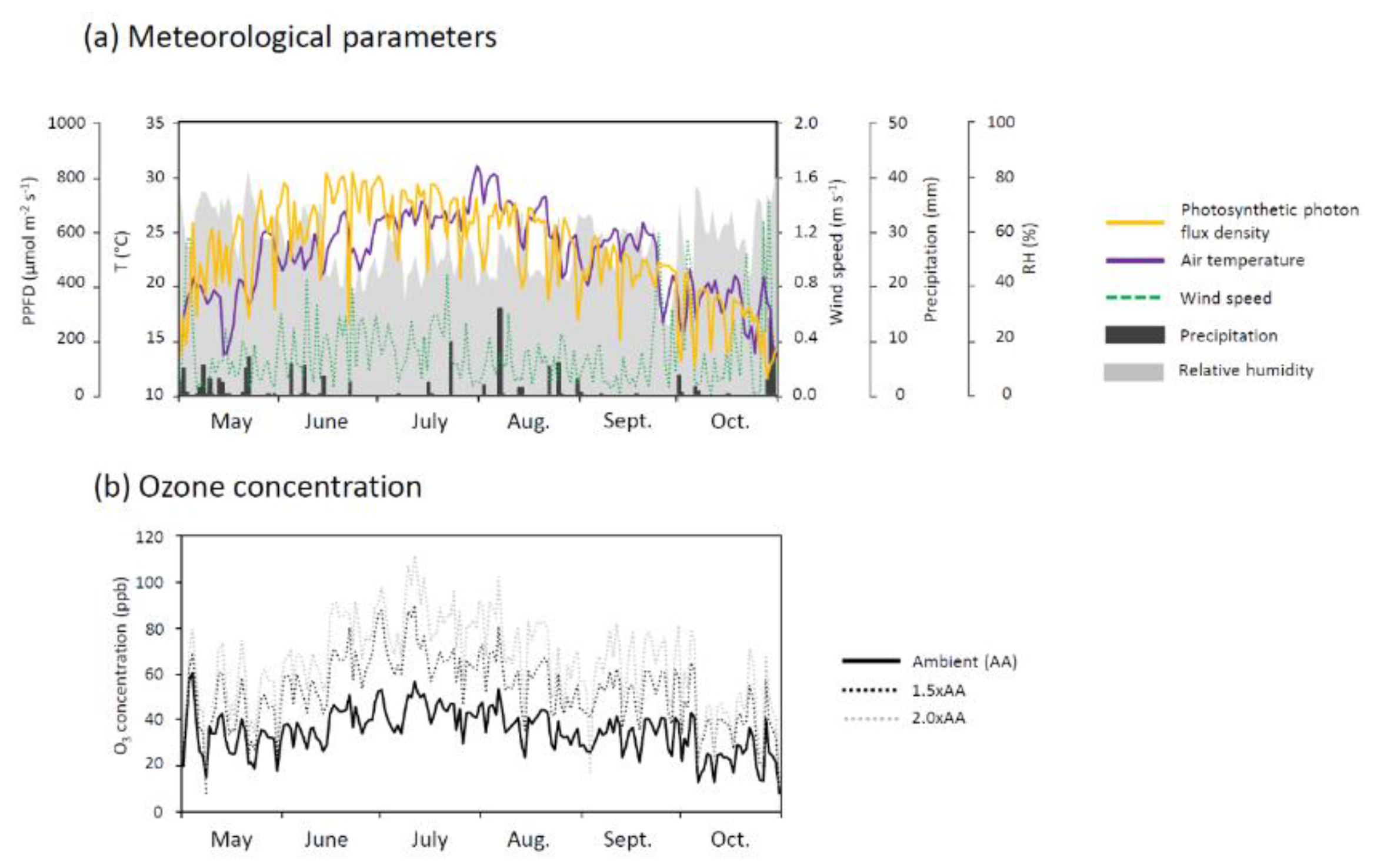
3.1. Ozone Concentration and Meteorological Factors
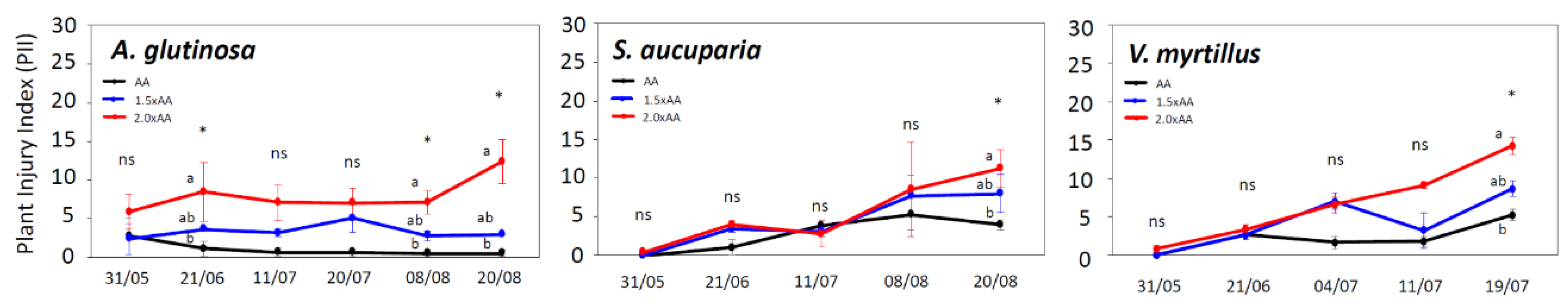
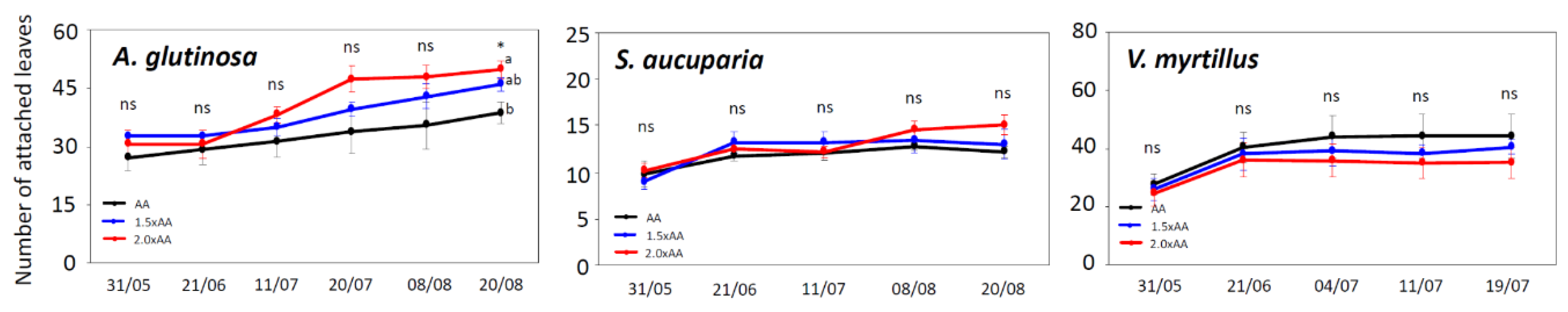
3.2. Ozone Visible Injury
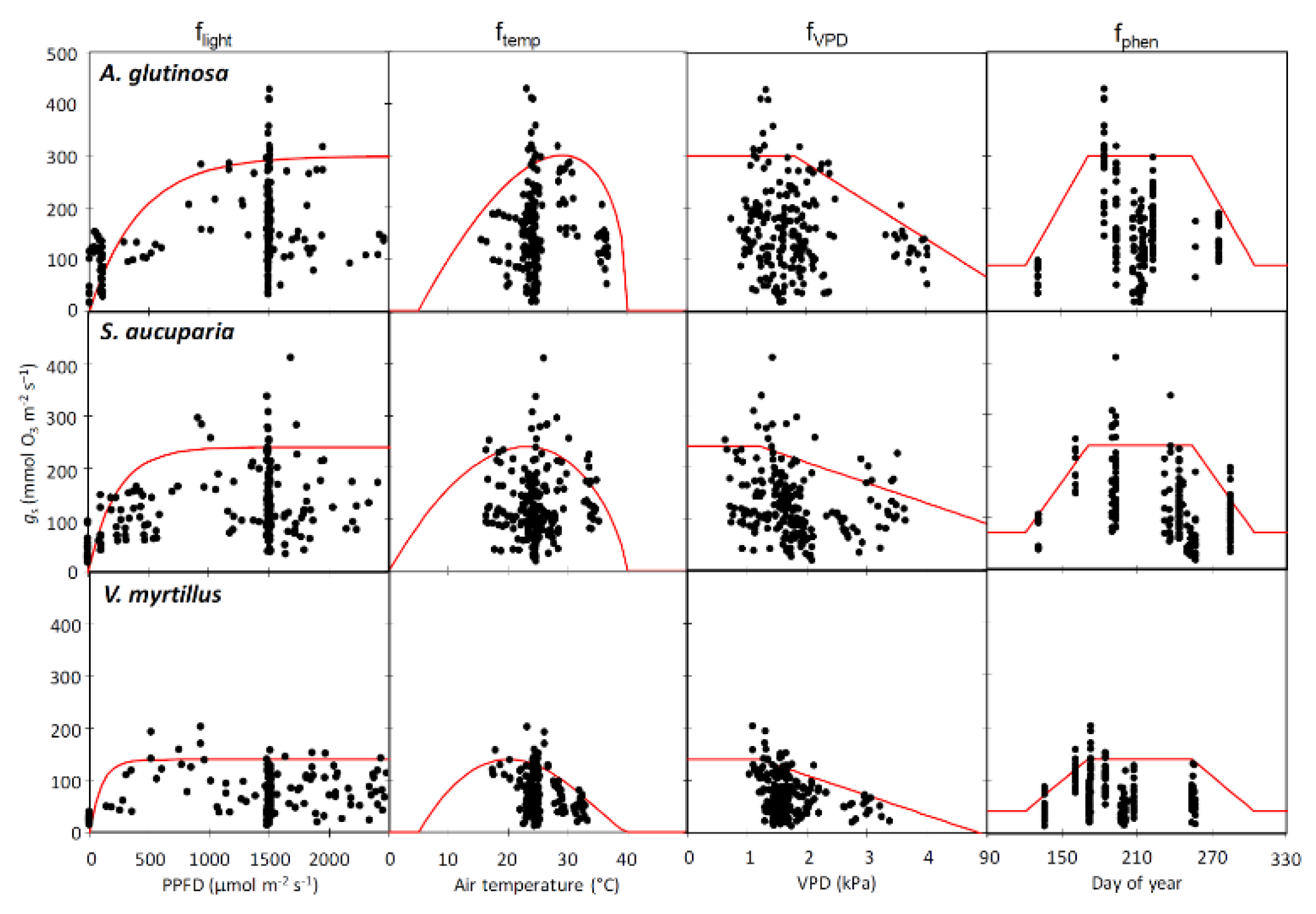
3.3. Parameterization of Stomatal Conductance Model
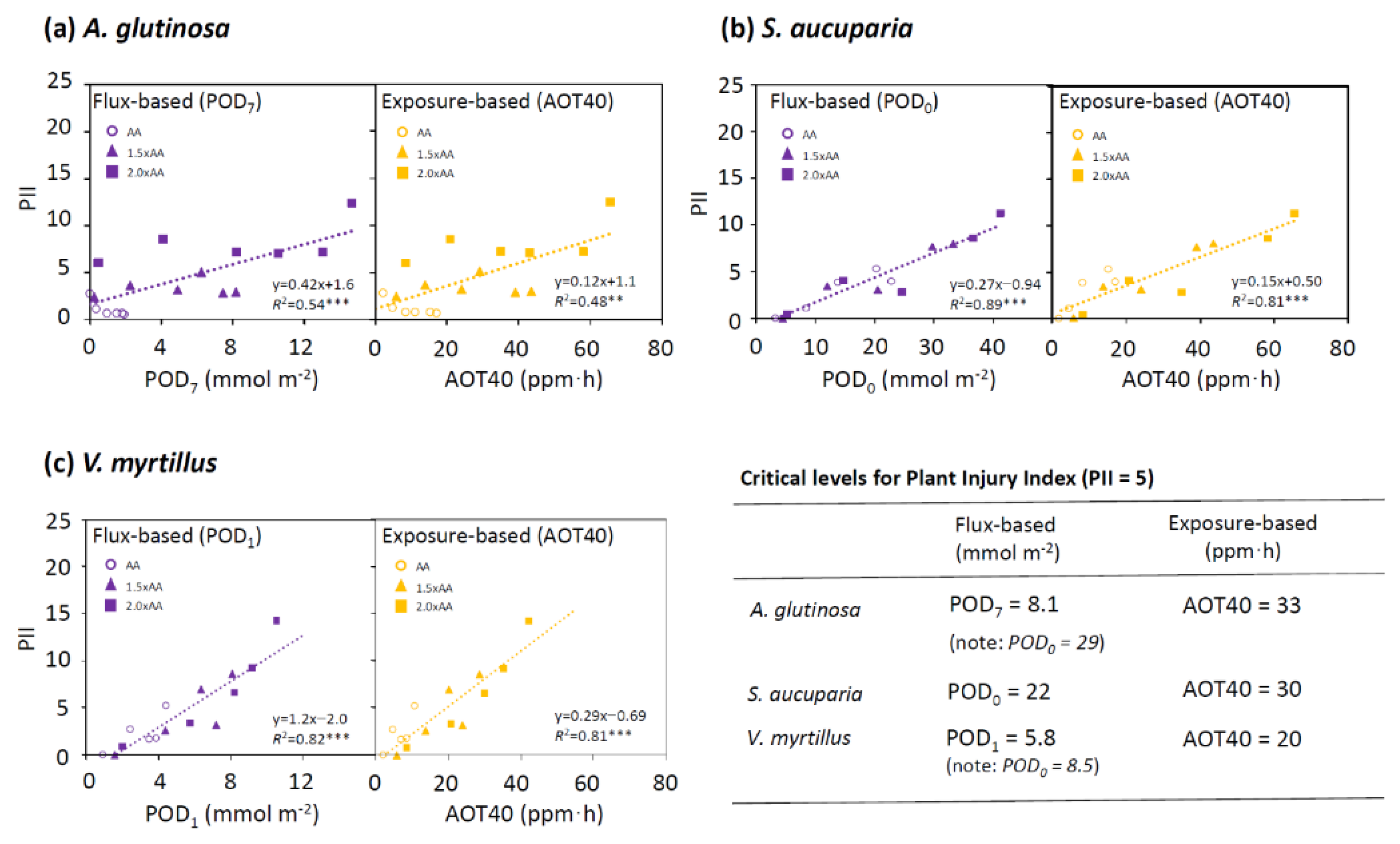
3.4. Dose-Response Relationship for Plant Injury Index
4. Discussion
4.1. New DO3SE Parameterization in Three Deciduous Tree Species
4.2. Flux-Based Assessment of Ozone Visible Injury
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grulke, N.E.; Heath, R.L. Ozone effects on plants in natural ecosystems. Plant Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lefohn, A.S.; Malley, C.S.; Smith, L.; Wells, B.; Hazucha, M.; Simon, H.; Naik, V.; Mills, G.; Schultz, M.G.; Paoletti, E.; et al. Tropospheric ozone assessment report: Global ozone metrics for climate change, human health, and crop/ecosystem research. Elem. Sci. Anthr. 2018, 6, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, G.; Pleijel, H.; Malley, C.S.; Sinha, B.; Cooper, O.; Schultz, M.; Neufeld, H.S.; Simpson, D.; Sharps, K.; Feng, Z.; et al. Tropospheric Ozone Assessment Report: Present day tropospheric ozone distribution and trends relevant to vegetation. Elem. Sci. Anthr. 2018, 6, 47. [Google Scholar] [CrossRef]
- Paoletti, E.; Ferrara, A.M.; Calatayud, V.; Cerveró, J.; Giannetti, F.; Sanz, M.J.; Manning, W.J. Deciduous shrubs for ozone bioindication: Hibiscus syriacus as an example. Environ. Pollut. 2009, 157, 865–870. [Google Scholar] [CrossRef]
- Hoshika, Y.; Omasa, K.; Paoletti, E. Whole-tree water use efficiency is decreased by ambient ozone and not affected by O3-induced stomatal sluggishness. PLoS ONE 2012, 7, e39270. [Google Scholar] [CrossRef] [Green Version]
- Innes, J.L.; Skelly, J.M.; Schaub, M. Ozone and broadleaved species. In A Guide to the Identification of Ozone-Induced Foliar Injury; Paul Haupt Verlag: Bern, Switzerland, 2001. [Google Scholar]
- Feng, Z.; Sun, J.; Wan, W.; Hu, E.; Calatayud, V. Evidence of widespread ozone-induced visible injury on plants in Beijing, China. Environ. Pollut. 2014, 193, 296–301. [Google Scholar] [CrossRef]
- Paoletti, E.; Alivernini, A.; Anav, A.; Badea, O.; Carrari, E.; Chibulescu, S.; Conte, A.; Ciriani, M.L.; Dalstein-Richier, L.; De Marco, A.; et al. Toward stomatal–flux based forest protection against ozone: The MOTTLES approach. Sci. Tot. Environ. 2019, 691, 516–527. [Google Scholar] [CrossRef]
- Fuhrer, J.; Skärby, L.; Ashmore, M.R. Critical levels for ozone effects on vegetation in Europe. Environ. Pollut. 1997, 97, 91–106. [Google Scholar] [CrossRef]
- Hůnová, I.; Schreiberová, M. Ambient ozone phytotoxic potential over the Czech forests as assessed by AOT40. iForest Biogeosci. For. 2012, 5, 153–162. [Google Scholar] [CrossRef]
- Baumgarten, M.; Werner, H.; Häberle, K.-H.; Emberson, L.D.; Fabian, P.; Matyssek, R. Seasonal ozone response of mature beech trees (Fagus sylvatica) at high altitude in the Bavarian forest (Germany) in comparison with young beech grown in the field and in phytotrons. Environ. Pollut. 2000, 109, 431–442. [Google Scholar] [CrossRef]
- Van der Heyden, D.; Skelly, J.; Innes, J.; Hug, C.; Zhang, J.; Landolt, W.; Bleuler, P. Ozone exposure thresholds and foliar injury on forest plants in Switzerland. Environ. Pollut. 2001, 111, 321–331. [Google Scholar] [CrossRef]
- Novak, K.; Skelly, J.M.; Schaub, M.; Kräuchi, N.; Hug, C.; Landolt, W.; Bleuler, P. Ozone air pollution and foliar injury development on native plants of Switzerland. Environ. Pollut. 2003, 125, 41–52. [Google Scholar] [CrossRef]
- Calatayud, V.; Cerveró, J.; Sanz, M.J. Foliar, physiological and growth responses of four maple species exposed to ozone. Wat. Air Soil Pollut. 2007, 185, 239–254. [Google Scholar] [CrossRef]
- Paoletti, E. Ozone impacts on forests. In CAB Reviews: Perspectives in Agriculture, Veterinary Science, Nutrition and Natural Resources; CABI: Wallingford, UK, 2007. [Google Scholar]
- CLRTAP. Mapping Critical Levels for Vegetation, Chapter III of Manual on methodologies and criteria for modelling and mapping critical loads and levels and air pollution effects, risks and trends, 2017. In UNECE Convention on Long-Range Transboundary Air Pollution; Programme Coordinating Centre, Federal Research Centre for Forestry and Forest Products: Hamburg, Germany, 2017; Available online: www.icpmapping.org (accessed on 8 November 2019).
- Sicard, P.; De Marco, A.; Dalstein-Richier, L.; Tagliaferro, F.; Paoletti, E. An epidemiological assessment of stomatal ozone flux-based critical levels for visible ozone injury in Southern European forests. Sci. Tot. Environ. 2016, 541, 729–741. [Google Scholar] [CrossRef]
- Paoletti, E. Impact of ozone on Mediterranean forests: A review. Environ. Pollut. 2006, 144, 463–474. [Google Scholar] [CrossRef]
- Schaub, M.; Calatayud, V. Assessment of visible foliar injury induced by ozone. In Forest Monitoring: Methods for Terrestrial Investigations in Europe with an Overview of North America and Asia; Ferretti, M., Fischer, R., Eds.; Elsevier: London, UK, 2013; pp. 205–221. [Google Scholar]
- Pell, E.J.; Brendley, B.W.; Sinn, J.P. Ozone-induced accelerated foliar senescence: Implications for toxicity and compensation. In Proceedings 1995 Meeting of the Northern Global Change Program; Hom, J., Birdsey, R., O’Brian, K., Eds.; Gen. Tech. Rep. NE-214; U.S. Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Radnor, PA, USA, 1996; pp. 13–19. [Google Scholar]
- Hoshika, Y.; De Carlo, A.; Baraldi, R.; Neri, L.; Carrari, E.; Agathokleous, E.; Zhang, L.; Fares, S.; Paoletti, E. Ozone-induced impairment of night-time stomatal closure in O3-sensitive poplar clone is affected by nitrogen but not by phosphorus enrichment. Sci. Tot. Environ. 2019, 692, 713–722. [Google Scholar] [CrossRef]
- Vogel, C.S.; Curtis, P.S.; Thomas, R.B. Growth and nitrogen accretion of dinitrogen-fixing Alnus glutinosa (L.) Gaertn. under elevated carbon dioxide. Plant Ecol. 1997, 130, 63–70. [Google Scholar] [CrossRef]
- Heide, O.M. Temperature rather than photoperiod controls growth cessation and dormancy in Sorbus species. J. Exp. Bot. 2011, 62, 5397–5404. [Google Scholar] [CrossRef]
- Selås, V.; Sønsteby, A.; Heide, O.M.; Opstad, N. Climatic and seasonal control of annual growth rhythm and flower formation in Vaccinium myrtillus (Ericaceae), and the impact on annual variation in berry production. Plant Ecol. Evolut. 2015, 148, 350–360. [Google Scholar] [CrossRef]
- Paoletti, E.; Materassi, A.; Fasano, G.; Hoshika, Y.; Carriero, G.; Silaghi, D.; Badea, O. A new-generation 3D ozone FACE (Free Air Controlled Exposure). Sci. Tot. Environ. 2017, 575, 1407–1414. [Google Scholar] [CrossRef]
- Alonso, R.; Elvira, S.; Sanz, M.J.; Gerosa, G.; Emberson, L.D.; Bermejo, B.; Gimeno, B.S. Sensitivity analysis of a parameterization of the stomatal component of the DO3SE model for Quercus ilex to estimate ozone fluxes. Environ. Pollut. 2008, 155, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Hoshika, Y.; Carrari, E.; Zhang, L.; Carriero, G.; Pignatelli, S.; Fasano, G.; Materassi, A.; Paoletti, E. Testing a ratio of photosynthesis to O3 uptake as an index for assessing O3-induced foliar visible injury in poplar trees. Environ. Sci. Pollut. Res. 2018, 25, 8113–8124. [Google Scholar] [CrossRef] [PubMed]
- Emberson, L.D.; Büker, P.; Ashmore, M.R. Assessing the risk caused by ground level ozone to European forest trees: A case study in pine, beech and oak across different climate regions. Environ. Pollut. 2007, 147, 454–466. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shang, B.; Yuan, X.; Feng, Z.; Calatayud, V. Relationships of CO2 assimilation rates with exposure- and flux-based O3 metrics in three urban tree species. Sci. Tot. Environ. 2018, 613–614, 233–239. [Google Scholar] [CrossRef]
- Oleksyn, J.; Karolewski, P.; Giertych, M.J.; Zytkowiak, R.; Reich, P.B.; Tjoelker, M.G. Primary and secondary host plants differ in leaf-level photosynthetic response to herbivory: Evidence from Alnus and Betula grazed by the alder beetle, Agelastica alni. New Phytol. 1998, 140, 239–249. [Google Scholar] [CrossRef]
- Kučerová, A.; Pokorný, J.; Radoux, M.; Nemcova, M.; Cadelli, D.; Dušek, J. Evapotranspiration of small-scale constructed wetlands planted with ligneous species. In Transformations of Nutrients in Natural and Constructed Wetlands; Vymazal, J., Ed.; Backhuys Publishers: Kerkwerve, The Netherlands, 2001; pp. 413–427. [Google Scholar]
- Kupper, P.; Ivanova, H.; Söber, A.; Rohula-Okunev, G.; Sellin, A. Night and daytime water relations in five fast-growing tree species: Effects of environmental and endogenous variables. Ecohydrology 2017, 11, e1927. [Google Scholar] [CrossRef]
- Marzuoli, R.; Gerosa, G.; Desotgiu, R.; Bussotti, F.; Ballarin-Denti, A. Ozone fluxes and foliar injury development in the ozone-sensitive poplar clone Oxford (Populus maximowiczii x Populus berolinensis). Tree Physiol. 2009, 29, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Hoshika, Y.; Watanabe, M.; Inada, N.; Koike, T. Modeling of stomatal ozone conductance for estimating ozone uptake of Fagus crenata under experimentally enhanced free-air ozone exposure. Wat. Air Soil Pollut. 2012, 223, 3893–3901. [Google Scholar] [CrossRef]
- Hoshika, Y.; Watanabe, M.; Inada, N.; Koike, T. Effects of ozone-induced stomatal closure on ozone uptake and ist changes due to leaf age in sun and shade leaves of Siebold’s beech. J. Agric. Meteorol. 2015, 71, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, P.S. Niche differentiation with respect to light utilization among coexisting dwarf shrubs in a subarctic woodland. Polar Biol. 1987, 8, 35–39. [Google Scholar] [CrossRef]
- Gerdol, R.; Iacumin, P.; Marchesini, R.; Bragazza, L. Water- and nutrient-use efficiency of a deciduous species, Vaccinium myrtillus, and an evergreen species, V. vitis-idaea, in a subalpine dwarf shrub heath in the southern Alps, Italy. Oikos 2000, 88, 19–32. [Google Scholar] [CrossRef]
- Tonteri, T.; Salemaa, M.; Rauto, P.; Hallikainen, V.; Korpela, L.; Merilä, P. Forest management regulates temporal change in the cover of boreal plant species. For. Ecol. Manag. 2016, 381, 115–124. [Google Scholar] [CrossRef]
- Claessens, H.; Oosterbaan, A.; Savill, P.; Rondeux, J. A review of the characteristics of black alder (Alnus glutinosa (L.) Gaertn.) and their implications for silvicultural practices. Forestry 2010, 83, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Giertych, M.; Karolewski, P.; Oleksyn, J. Carbon allocation in seedlings of deciduous tree species depends on their shade tolerance. Acta Physiol. Plant. 2015, 37, 216. [Google Scholar] [CrossRef] [Green Version]
- Larcher, W. Physiological Plant Ecology, 4th ed.; Springer: New York, NY, USA, 2003. [Google Scholar]
- Matyssek, R.; Clarke, N.; Cudlin, P.; Mikkelsen, T.N.; Tuovinen, J.P.; Wieser, G.; Paoletti, E. Climate Change, Air Pollution and Global Challenges: Understanding and persPectives from Forest Research. In Developments in Environmental Science; Elsevier: Amsterdam, The Netherlands, 2013; 622p. [Google Scholar]
- Pellegrini, E.; Hoshika, Y.; Paoletti, E. Ozone effects on the quality of Common Bilberry fruits. Unpublished work.
- Vandermeiren, K.; De Temmerman, L.; Hookham, N. Ozone sensitivity of Phaseolus vulgaris in relation to cultivar differences, growth stage and growing conditions. Wat. Air Soil Pollut. 1995, 85, 1455–1460. [Google Scholar] [CrossRef]
- Morgan, P.B.; Ainsworth, E.A.; Long, S.P. How does elevated ozone impact soybean? A meta-analysis of photosynthesis, growth and yield. Plant Cell Environ. 2003, 26, 1317–1328. [Google Scholar] [CrossRef]
- Feng, Z.; Pang, J.; Nouchi, I.; Kobayashi, K.; Yamakawa, T.; Zhu, J. Apoplastic ascorbate contributes to the differential ozone sensitivity in two varieties of winter wheat under fully open-air field conditions. Environ. Pollut. 2010, 158, 3539–3545. [Google Scholar] [CrossRef]
- Braun, S.; Schindler, C.; Rihm, B. Growth trends of beech and Norway spruce in Switzerland: The role of nitrogen deposition, ozone, mineral nutrition and climate. Sci. Tot. Environ. 2017, 599–600, 637–646. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | A. glutinosa | S. aucuparia | V. myrtillus |
|---|---|---|---|
| Onset date | 18 May | 21 May | 26 May |
| POD0 (mmol m−2) | 3.2 | 4.3 | 3.5 |
| AOT40 (ppm·h) | 5.6 | 6.3 | 8.9 |
| Parameter | Unit | A. glutinosa | S. aucuparia | V. myrtillus | |
|---|---|---|---|---|---|
| gmax | (mmol O3 m−2 PLA s−1) | 300 | 240 | 140 | |
| fmin | (fraction) | 0.13 | 0.17 | 0.17 | |
| Astart | (day of year) | 121 | 121 | 121 | |
| Aend | (day of year) | 304 | 304 | 304 | |
| fphen | fphen_a | (days) | 50 | 50 | 50 |
| fphen_b | (days) | 50 | 50 | 50 | |
| fphen_c | (fraction) | 0.3 | 0.3 | 0.3 | |
| fphen_d | (fraction) | 0.3 | 0.3 | 0.3 | |
| flight | a | (constant) | 0.0024 | 0.0043 | 0.0104 |
| ftemp | Topt | (°C) | 29 | 23 | 20 |
| Tmin | (°C) | 5 | 0 | 5 | |
| Tmax | (°C) | 40 | 40 | 40 | |
| fVPD | VPDmax | (kPa) | 1.8 | 1.2 | 1.2 |
| VPDmin | (kPa) | 5.7 | 7.0 | 4.7 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoshika, Y.; Carrari, E.; Mariotti, B.; Martini, S.; De Marco, A.; Sicard, P.; Paoletti, E. Flux-Based Ozone Risk Assessment for a Plant Injury Index (PII) in Three European Cool-Temperate Deciduous Tree Species. Forests 2020, 11, 82. https://doi.org/10.3390/f11010082
Hoshika Y, Carrari E, Mariotti B, Martini S, De Marco A, Sicard P, Paoletti E. Flux-Based Ozone Risk Assessment for a Plant Injury Index (PII) in Three European Cool-Temperate Deciduous Tree Species. Forests. 2020; 11(1):82. https://doi.org/10.3390/f11010082
Chicago/Turabian StyleHoshika, Yasutomo, Elisa Carrari, Barbara Mariotti, Sofia Martini, Alessandra De Marco, Pierre Sicard, and Elena Paoletti. 2020. "Flux-Based Ozone Risk Assessment for a Plant Injury Index (PII) in Three European Cool-Temperate Deciduous Tree Species" Forests 11, no. 1: 82. https://doi.org/10.3390/f11010082
APA StyleHoshika, Y., Carrari, E., Mariotti, B., Martini, S., De Marco, A., Sicard, P., & Paoletti, E. (2020). Flux-Based Ozone Risk Assessment for a Plant Injury Index (PII) in Three European Cool-Temperate Deciduous Tree Species. Forests, 11(1), 82. https://doi.org/10.3390/f11010082