Variability and Plasticity in Cuticular Transpiration and Leaf Permeability Allow Differentiation of Eucalyptus Clones at an Early Age

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Stomatal Characteristics
2.3. Cuticular Transpiration
2.4. Data Analysis
3. Results
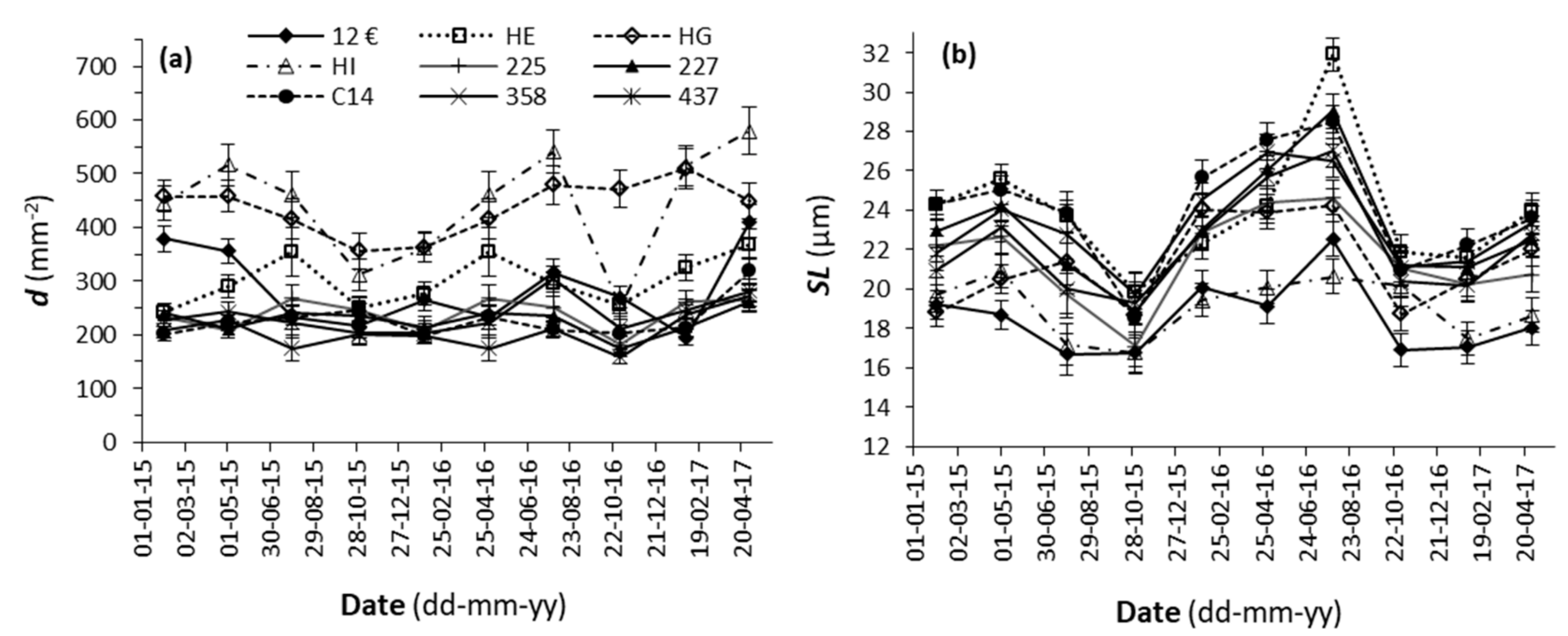
3.1. Stomatal Characteristics
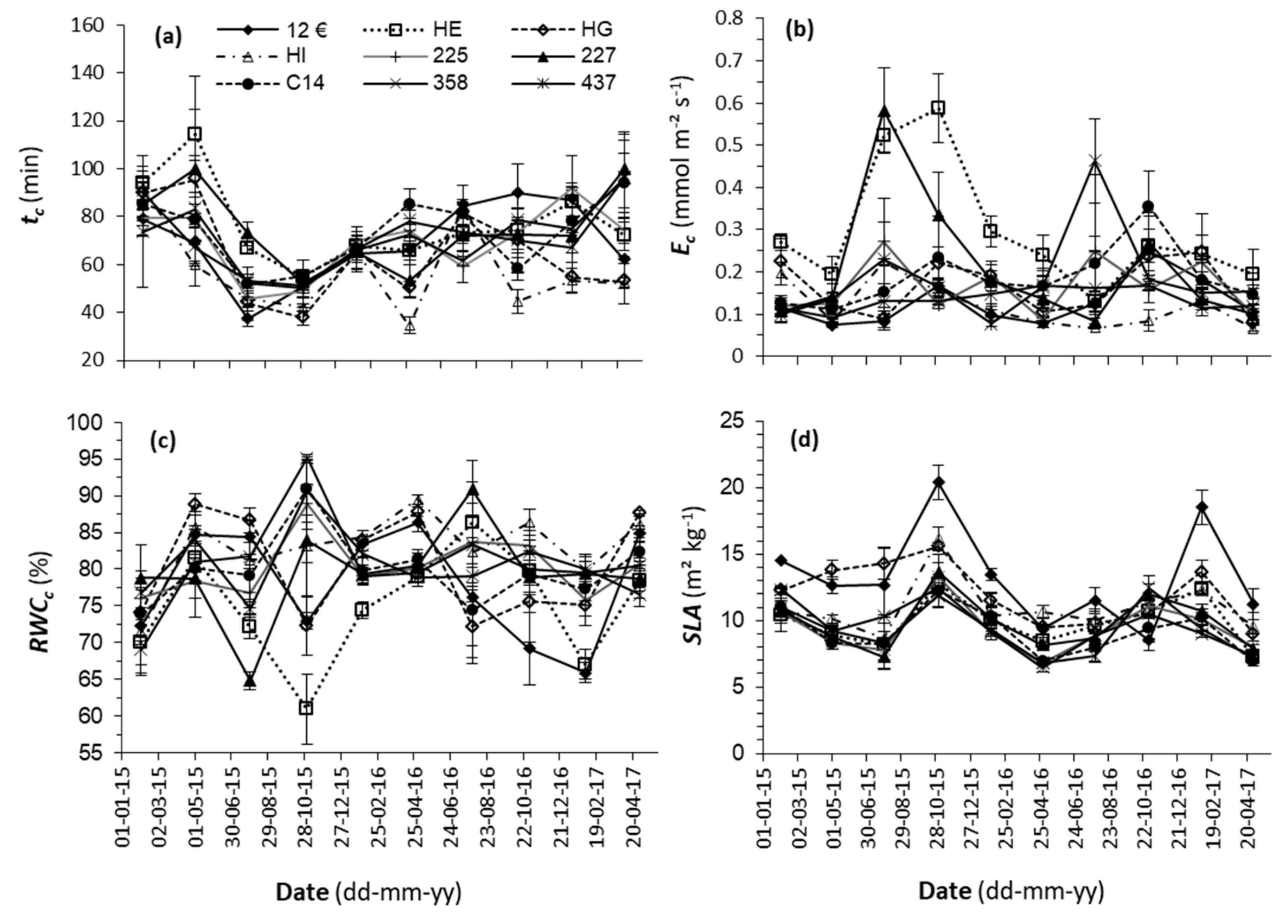
3.2. Cuticular Transpiration
4. Discussion
4.1. Stomatal Characteristics
4.2. Cuticular Transpiration and SLA
5. Conclusions
- -
- All the clones under study showed seasonal variations in d and stomatal size, SLA, and Ec, as well as in RWCc and the tc, despite the substrate being constantly humidified to field capacity and fertilized, in response to stimuli such as light radiation, photoperiod, temperature, and/or the air’s evaporative demand to acclimatize to the environmental conditions.
- -
- Each clone adjusted its own d and size values to acclimatize its stomata to the growth conditions. The maximum amount of water transpired with fully open stomata might depend on other internal and external factors. Thus, the criteria of size and d alone are not sufficient to differentiate between clones, at least for this study.
- -
- The HE and HG clones presented poor stomatal control as they showed high transpiration rates when Ψ was between −1.0 and −2.0 MPa, which represent a risk in cases of prolonged drought. The HI clone, on the other hand, conserved internal water very efficiently because it closed stomata sooner and at a lesser degree of dehydration.
- -
- Taking into account only the morpho-physiological parameters measured in this study and the genotypes considered, the clone known to resist to droughts, C14, was characterized by low values of SLA and Ec, high RWCc, low seasonal plasticity regarding d, and good stomatal control. These characteristics were shared by the other E. globulus clones, though the clones of hybrids differed as they presented less favorable properties for drought resistance such as lower epidermis impermeability (HE, HG, 12€), higher SLA (12€, HG), and lower stomatal control under conditions of moderate water stress (HE, HG).
- -
- These analyzed eucalyptus clones showed they had genetic variability for drought resistance. Gains could thus potentially be obtained through the selection and implantation of improved populations. However, in situations of water stress, other morpho-physiological properties should be studied (e.g., WUE, xylem anatomy and cavitation vulnerability, osmotic adjustment capacity, etc.) together with the characteristics studied here.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schlichting, C.D. The evolution of phenotypic plasticity in plants. Annu. Rev. Ecol. Syst. 1986, 17, 677–693. [Google Scholar] [CrossRef]
- Bussis, D.; Von Groll, U.; Fisahn, J.; Altman, T. Stomatal aperture can compensate altered stomatal density in Arabidopsis thaliana at growth light conditions. Funct. Plant Biol. 2006, 33, 1037–1043. [Google Scholar] [CrossRef]
- Poorter, L.; Rozendaal, D.M.A. Leaf size and leaf display of thirty-eight tropical tree species. Oecologia 2008, 158, 35–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejía de Tafur, M.S.; Burbano Diaz, R.A.; García Díaz, M.A.; Baena García, D. Respuesta fotosintética de Eucalyptus grandis W. Hill a la disponibilidad de agua en el suelo y a la intensidad de luz. Acta Agron. 2014, 63, 311–317. [Google Scholar] [CrossRef]
- Haworth, M.; Scutt, C.P.; Douthe, C.; Marino, G.; Gomes, M.T.G.; Loreto, F.; Flexas, J.; Centritto, M. Allocation of the epidermis to stomata relates to stomatal physiological control: Stomatal factors involved in the diversification of the angiosperms and development of amphistomaty. Environ. Exp. Bot. 2018, 151, 55–63. [Google Scholar] [CrossRef]
- Haworth, M.; Marino, G.; Cosentino, S.L.; Brunetti, C.; De Carlo, A.; Avola, G.; Riggi, E.; Loreto, F.; Centritto, M. Increased free abscisic acid during drought enhances stomatal sensitivity and modifies stomatal behaviour in fast growing giant reed (Arundo donax L.). Environ. Exp. Bot. 2018, 147, 116–124. [Google Scholar] [CrossRef]
- García-Mata, C.; Lamattina, L. Nitric oxide induces stomatal closure and enhances the adaptive plant responses against drought stress. Plant Physiol. 2001, 126, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Shimazaki, K.; Doi, M.; Assmann, S.M.; Kinoshita, T. Light regulation of stomatal movement. Annu. Rev. Plant Biol. 2007, 58, 219–247. [Google Scholar] [CrossRef] [Green Version]
- Gerardin, T.; Douthe, C.; Flexas, J.; Brendel, O. Shade and drought growth conditions strongly impact dynamic responses of stomata to variations in irradiance in Nicotiana tabacum. Environ. Exp. Bot. 2018, 153, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Dai, Z.; Edwards, G.E.; Ku, M.S.B. Control of photosynthesis and stomatal conductance in Ricinus communis L. (castor bean) by leaf to air vapor pressure deficit. Plant Physiol. 1992, 99, 1426–1434. [Google Scholar]
- Mott, K.A.; Peak, D. Testing a vapour-phase model of stomatal responses to humidity. Plant Cell Environ. 2013, 36, 936–944. [Google Scholar] [CrossRef] [PubMed]
- Haworth, M.; Elliott-Kingston, C.; McElwain, J.C. Stomatal control as a driver of plant evolution. J. Exp. Bot. 2011, 62, 2419–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haworth, M.; Killi, D.; Materassi, A.; Raschi, A. Co-ordination of stomatal physiological behavior and morphology with carbon dioxide determines stomatal control. Am. J. Bot. 2015, 102, 677–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Givnish, T.J. Adaptation to sun and shade: A whole-plant perspective. Funct. Plant Biol. 1988, 15, 63–92. [Google Scholar] [CrossRef] [Green Version]
- Woodward, F.I.; Kelly, C. The influence of CO2 concentration on stomatal density. New Phytol. 1995, 131, 311–327. [Google Scholar] [CrossRef]
- Hetherington, A.M.; Woodward, F.I. The role of stomata in sensing and driving environmental change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef]
- Lammertsma, E.I.; de Boer, H.J.; Dekker, S.C.; Dilcher, D.L.; Lotter, A.F.; Wagner-Cremer, F. Global CO2 rise leads to reduced maximum stomatal conductance in Florida vegetation. Proc. Natl. Acad. Sci. USA 2011, 108, 4035–4040. [Google Scholar] [CrossRef] [Green Version]
- Nouvellon, Y.; Laclau, J.P.; Epron, D.; Kinana, A.; Mabiala, A.; Roupsard, O.; Bonnefond, J.M.; le Marie, G.; Marsden, C.; Bontemps, J.D.; et al. Within-stand and seasonal variations of specific leaf area in a clonal Eucalyptus plantation in the Republic of Congo. For. Ecol. Manag. 2010, 259, 1796–1807. [Google Scholar] [CrossRef]
- Canny, M.J.; Huang, C.X. Leaf water content and palisade cell size. New Phytol. 2006, 170, 75–85. [Google Scholar] [CrossRef]
- Ali, I.; Abbas, S.Q.; Hameed, M.; Naz, N.; Zafar, S.; Kanwal, S. Leaf anatomical adaptations in some exotic species of Eucalyptus L’Hér. (Myrtaceae). Pak. J. Bot. 2009, 41, 2717–2727. [Google Scholar]
- Oteros, J.; García-Mozo, H.; Vázquez, L.; Mestre, A.; Domínguez-Vilches, E.; Galán, C. Modelling olive phenological response to weather and topography. Agric. Ecosyst. Environ. 2013, 179, 62–68. [Google Scholar] [CrossRef]
- Navarrete-Campos, D.; Bravo, L.A.; Rubilar, R.A.; Emhart, V.; Sanhueza, R. Drought effects on water use efficiency, freezing tolerance and survival of Eucalyptus globulus and Eucalyptus globulus × nitens cuttings. New For. 2013, 44, 119–134. [Google Scholar] [CrossRef]
- FAO. El Estado de los Bosques del Mundo 2016. Los Bosques y la Agricultura: Desafíos y Oportunidades en Relación con el uso de la Tierra; FAO: Rome, Italy, 2016; 119p. [Google Scholar]
- Nunes, B.H.S.; Rezende, G.D.S.P.; Ramalho, M.A.P.; Santos, J.B. Implicações da interação genótipo × ambientes na seleção de clones de eucalipto. Cerne 2002, 8, 49–58. [Google Scholar]
- Berger, R.; Schneider, P.R.; Finger, C.A.G.; Haseilen, C.R. Efeito do espaçamento e da adubação no crescimento de um clone de Eucalyptus saligna Smith. Cienc. Florest. 2002, 12, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Leslie, A.D.; Mencuccini, M.; Perks, M.P.; Wilson, E.R. A review of the suitability of eucalypts for short rotation forestry for energy in the UK. New For. 2019, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Almeida, A.C.; Smethurst, P.J.; Siggins, A.; Cavalcante, R.B.L.; Borges, N.J. Quantifying the effects of Eucalyptus plantations and management on water resources at plot and catchment scales. Hydrol. Process. 2016, 30, 4687–4703. [Google Scholar] [CrossRef]
- Silva, P.; Campoe, O.; Paula, R.; Lee, D. Seedling growth and physiological responses of sixteen eucalypt taxa under controlled water regime. Forests 2016, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- Costa e Silva, F.; Shvaleva, A.; Broetto, F.; Ortuño, M.F.; Rodrigues, M.L.; Almeida, M.H.; Chaves, M.M.; Pereira, J.S. Acclimation to short-term low temperatures in two Eucalyptus globulus clones with contrasting drought resistance. Tree Physiol. 2008, 29, 77–86. [Google Scholar] [CrossRef]
- Silva, C.D.; Nascimento, J.S.; Scarpinati, E.A.; Paula, R.C. Classification of Eucalyptus urograndis hybrids under different water availability based on biometric traits. For. Syst. 2014, 23, 209–215. [Google Scholar] [CrossRef]
- Silva, I.M.A.; de Souza, M.W.R.; Rodrigues, A.C.P.; Correia, L.P.S.; Veloso, R.V.S.; dos Santos, J.B.; Titon, M.; Gonçalves, J.F.; de Laia, M.L. Determination of parameters for selection of Eucalyptus clones tolerant to drought. Afr. J. Agric. Res. 2016, 11, 3940–3949. [Google Scholar]
- Li, C. Population differences in water-use efficiency of Eucalyptus microtheca seedlings under different watering regimes. Physiol. Plant. 2000, 108, 134–139. [Google Scholar] [CrossRef]
- Jiménez, E.; Vega, J.A.; Pérez-Gorostiaga, P.; Fonturbel, T.; Cuiñas, P.; Fernández, C. Evaluación de la transpiración de Eucalyptus globulus mediante la densidad de flujo de savia y su relación con variables meteorológicas y dendrométricas. Bol. Inf. CIDEU 2007, 119–138. Available online: https://www.uhu.es/cideu/PreWeb/Boletin/Boletin3/BolInf3CIDEU119-138.pdf (accessed on 18 March 2019).
- Almeida, A.C.; Soares, J.V.; Landsberg, J.J.; Rezende, G.D. Growth and water balance of Eucalyptus grandis hybrid plantations in Brazil during a rotation for pulp production. For. Ecol. Manag. 2007, 251, 10–21. [Google Scholar] [CrossRef]
- Bhusal, N.; Bhusal, S.J.; Yoon, T.M. Comparisons of physiological and anatomical characteristics between two cultivars in bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2018, 231, 73–81. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.G.; Yoon, T.M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- McDowell, N.G.; Beerling, D.J.; Breshears, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef]
- Pita, P.; Cañas, I.; Soria, F.; Ruiz, F.; Toval, G. Use of physiological traits in tree breeding for improved yield in drought-prone environments. The case of Eucalyptus globulus. For. Syst. 2005, 14, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Fernández, M.; Tapias, R.; Alesso, P. Adaptación a la sequía y necesidades hídricas de Eucalyptus globulus Labill. en Huelva. Bol. Inf. CIDEU 2010, 8, 31–41. Available online: http://rabida.uhu.es/dspace/handle/10272/4560 (accessed on 20 March 2019).
- Moraes, C.B.; Freitas, T.C.M.; Pieroni, G.B.; Resende, M.D.V.; Zimback, L.; Mori, E.S. Estimativas de parâmetros genéticos para seleção precoce de clones de Eucalyptus para região com ocorrência de geadas. Sci. For. 2014, 42, 219–227. [Google Scholar]
- Flores, T.B.; Alvares, C.A.; Souza, V.C.; Stape, J.L. Eucalyptus no Brasil—Zoneamento Climático e Guia Para Identificação; IPEF: Piracicaba, Brazil, 2016; 447p. [Google Scholar]
- Héroult, A.; Lin, Y.S.; Bourne, A.; Medlyn, B.E.; Ellsworth, D.S. Optimal stomatal conductance in relation to photosynthesis in climatically contrasting Eucalyptus species under drought. Plant Cell Environ. 2013, 36, 262–274. [Google Scholar] [CrossRef]
- Carevic, F.; Fernández, M.; Alejano, R.; Vázquez-Piqué, J.; Tapias, R.; Corral, E.; Domingo, J. Plant water relations and edaphoclimatic conditions affecting acorn production in a holm oak (Quercus ilex L. ssp. ballota) open woodland. Agrofor. Syst. 2010, 78, 299–308. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Tuffi Santos, L.D.; Iarema, L.; Thadeo, M.; Ferreira, F.A.; Meira, R.M.S.A. Características da epiderme foliar de eucalipto e seu envolvimento com a tolerância ao glyphosate. Planta Daninha 2006, 23, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Camargo, M.A.B.; Marenco, R.A. Density, size and distribution of stomata in 35 rainforest trees species in Central Amazonia. Acta Amazon. 2011, 41, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Carins Murphy, M.R.; Jordan, G.J.; Brodribb, T.J. Acclimation to humidity modifies the link between leaf size and the density of veins and stomata. Plant Cell Environ. 2014, 37, 124–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eksteen, A.B.; Grzeskowiak, V.; Jones, N.B.; Pammenter, N.W. Stomatal characteristics of Eucalyptus grandis clonal hybrids in response to water stress. S. For. 2013, 75, 105–111. [Google Scholar] [CrossRef]
- Gharun, M.; Turnbull, T.L.; Pfautsch, S.; Adams, M.A. Stomatal structure and physiology do not explain differences in water use among montane eucalypts. Oecologia 2015, 177, 1171–1181. [Google Scholar] [CrossRef]
- Araus, J.L.; Alegre, L.; Tapia, L.; Calafell, R.; Serret, M.D. Relationship between photosynthetic capacity and leaf structure in several shade plants. Am. J. Bot. 1986, 73, 1760–1770. [Google Scholar] [CrossRef]
- Pearce, D.W.; Millard, S.; Bray, D.F.; Rood, S.B. Stomatal characteristics of riparian poplar species in a semi-arid environment. Tree Physiol. 2006, 26, 211–218. [Google Scholar] [CrossRef]
- Bakker, J.C. Effects of humidity on stomatal density and its relation to leaf conductance. Sci. Hortic. 1991, 48, 205–212. [Google Scholar] [CrossRef]
- Toral, M.; Manríquez, A.; Navarro-Cerrillo, R.; Tersi, D.; Naulin, P. Características de los estomas, densidad e índice estomático en secuoya (Sequoia sempervirens) y su variación en diferentes plantaciones de Chile. Bosque 2010, 31, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhou, G. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. J. Exp. Bot. 2008, 59, 3317–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drake, P.L.; Froend, R.H.; Franks, P.J. Smaller, faster stomata: Scaling of stomatal size, rate of response, and stomatal conductance. J. Exp. Bot. 2013, 64, 495–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, D.; Douthe, C.; Flexas, J. Differential coordination of stomatal conductance, mesophyll conductance, and leaf hydraulic conductance in response to changing light across species. Plant Cell Environ. 2018, 41, 436–450. [Google Scholar] [CrossRef]
- Elliott-Kingston, C.; Haworth, M.; Yearsley, J.M.; Batke, S.P.; Lawson, T.; McElwain, J.C. Does size matter? Atmospheric CO2 may be a stronger driver of stomatal closing rate than stomatal size in taxa that diversified under low CO2. Front. Plant Sci. 2016, 7, 1253. [Google Scholar] [CrossRef] [Green Version]
- Kröber, W.; Bruelheide, H. Transpiration and stomatal control: A cross-species study of leaf traits in 39 evergreen and deciduous broadleaved subtropical tree species. Trees 2014, 28, 901–914. [Google Scholar] [CrossRef]
- Franks, P.J.; Drake, P.L.; Beerling, D.J. Plasticity in maximum stomatal conductance constrained by negative correlation between stomatal size and density: An analysis using Eucalyptus globulus. Plant Cell Environ. 2009, 32, 1737–1748. [Google Scholar] [CrossRef]
- Correia, B.; Pintó-Marijuan, M.; Neves, L.; Brossa, R.; Dias, M.C.; Costa, A.; Castro, B.B.; Araújo, C.; Santos, C.; Chaves, M.M.; et al. Water stress and recovery in the performance of two Eucalyptus globulus clones: Physiological and biochemical profiles. Physiol. Plant. 2014, 150, 580–592. [Google Scholar] [CrossRef]
- Hernandez, M.J.; Montes, F.; Ruiz, F.; Lopez, G.; Pita, P. The effect of vapour pressure deficit on stomatal conductance, sap pH and leaf-specific hydraulic conductance in Eucalyptus globulus clones grown under two watering regimes. Ann. Bot. 2016, 117, 1063–1071. [Google Scholar] [CrossRef] [Green Version]
- Merchant, A.; Callister, A.; Arndt, S.; Tausz, M.; Adams, M. Contrasting physiological responses of six Eucalyptus species to water deficit. Ann. Bot. 2007, 100, 1507–1515. [Google Scholar] [CrossRef]
- Mendes, H.S.J.; Paula, N.F.; Scarpinatti, E.A.; Paula, R.C. Respostas fisiológicas de genótipos de Eucalyptus grandis × E. urophylla à disponibilidade hídrica e adubação potássica. Cerne 2013, 19, 603–611. [Google Scholar] [CrossRef] [Green Version]
- Andivia, E.; Carevic, F.; Fernández, M.; Alejano, R.; Vázquez-Piqué, J.; Tapias, R. Seasonal evolution of water status after outplanting of two provenances of holm oak nursery seedlings. New For. 2012, 43, 815–824. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Water Deficits and Plant Growth, Volume III: Plant Responses and Control of Water Balance, 1st ed.; Academic Press: New York, NY, USA; London, UK, 1972; 382p. [Google Scholar]
- Fernández, M.; García-Albalá, J.; Andivia, E.; Alaejos, J.; Tapias, R.; Menéndez, J. Sickle bush (Dichrostachys cinerea L.) field performance and physical-chemical property assessment for energy purposes. Biomass Bioenergy 2015, 81, 483–489. [Google Scholar] [CrossRef]
- Eamus, D.; Taylor, D.T.; Macinnis-Ng, C.M.; Shanahan, S.; De Silva, L. Comparing model predictions and experimental data for the response of stomatal conductance and guard cell turgor to manipulations of cuticular conductance, leaf-to-air vapour pressure difference and temperature: Feedback mechanisms are able to account for all observations. Plant Cell Environ. 2008, 31, 269–277. [Google Scholar]
- Li, H.; Madden, J.L.; Potts, B.M. Variation in leaf waxes of the Tasmanian Eucalyptus species—I. Subgenus Symphyomyrtus. Biochem. Syst. Ecol. 1997, 25, 631–657. [Google Scholar] [CrossRef]
- Migacz, I.P.; Raeski, P.A.; Almeida, V.P.; Raman, V.; Nisgoski, S.; Muniz, G.I.B.; Farago, P.V.; Khan, I.A.; Budel, J.M. Comparative leaf morpho-anatomy of six species of Eucalyptus cultivated in Brazil. Rev. Bras. Farmacogn. 2018, 28, 273–281. [Google Scholar] [CrossRef]
- Fernández, M.; Alaejos, J.; Andivia, E.; Vázquez-Piqué, J.; Ruiz, F.; López, F.; Tapias, R. Eucalyptus × urograndis biomass production for energy purposes exposed to a Mediterranean climate under different irrigation and fertilisation regimes. Biomass Bioenergy 2018, 111, 22–30. [Google Scholar] [CrossRef]
- Sefton, C.A.; Montagu, K.D.; Atwell, B.J.; Conroy, J.P. Anatomical variation in juvenile eucalypt leaves accounts for differences in specific leaf area and CO2 assimilation rates. Aust. J. Bot. 2002, 50, 301–310. [Google Scholar] [CrossRef]
- Yao, R.L.; Glencross, K.; Nichols, J.D. Difference in shade tolerance affects foliage–sapwood response to thinning by two eucalypts. S. For. 2014, 76, 93–100. [Google Scholar] [CrossRef]
- Tyree, M.T.; Richter, H. Alternative Methods of Analysing Water Potential Isotherms: Some Cautions and Clarifications. J. Exp. Bot. 1981, 32, 643–653. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variables | Measurement Dates | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Feb 2015 | May 2015 | July 2015 | Nov 2015 | Feb 2016 | May 2016 | July 2016 | Nov 2016 | Feb 2017 | May 2017 | |
| T90 a (°C) | 16.3 | 19.9 | 30.0 | 24.9 | 19.8 | 19.3 | 28.6 | 27.3 | 17.9 | 20.9 |
| t90 b (°C) | 3.3 | 6.9 | 15.1 | 14.0 | 8.5 | 7.5 | 14.4 | 13.8 | 5.7 | 8.0 |
| RH90 a (%) | 102.1 | 100.1 | 89.6 | 95.8 | 95.2 | 93.6 | 86.7 | 86.9 | 89.8 | 91.6 |
| rh90 b (%) | 59.0 | 52.3 | 33.1 | 51.0 | 52.7 | 45.9 | 37.0 | 42.8 | 50.5 | 46.3 |
| R90 c (MJ m−2 d−1) | 9.3 | 16.2 | 26.3 | 16.0 | 9.1 | 16.8 | 26.2 | 17.1 | 9.5 | 16.2 |
| Clone | d (mm−2) | SL (μm) | SW/SL | RWCc (%) | tc (min) | Mc (%) | Ec (mmol m−2 s−1) | SLA (m2 kg−1) |
|---|---|---|---|---|---|---|---|---|
| 12€ | 279 ± 7 b | 18.5 ± 0.3 a | 0.77 ± 0.01 ab | 78.2 ± 1.3 ab | 70.2 ± 3.0 b | 61.8 ± 1.0 a | 0.12 ± 0.02 a | 13.4 ± 0.6 c |
| HE | 297 ± 9 b | 23.9 ± 0.3 d | 0.75 ± 0.01 a | 74.8 ± 1.4 a | 80.6 ± 3.7 b | 59.9 ± 0.9 a | 0.27 ± 0.02 b | 9.9 ± 0.3 ab |
| HG | 435 ± 11 c | 21.3 ± 0.3 b | 0.75 ± 0.01 a | 80.9 ± 1.3 ab | 64.5 ± 3.8 ab | 61.7 ± 0.8 a | 0.16 ± 0.02 a | 12.2 ± 0.5 c |
| HI | 431 ± 11 c | 19.1 ± 0.3 a | 0.75 ± 0.01 a | 83.4 ± 1.0 b | 55.5 ± 3.2 a | 61.4 ± 0.8 a | 0.12 ± 0.01 a | 11.0 ± 0.4 bc |
| 225 | 238 ± 6 a | 21.6 ± 0.3 bc | 0.77 ± 0.01 ab | 79.7 ± 1.1 ab | 73.4 ± 3.6 b | 61.0 ± 1.0 a | 0.16 ± 0.02 a | 9.2 ± 0.3 a |
| 227 | 225 ± 6 a | 23.0 ± 0.3 cd | 0.77 ± 0.01 ab | 79.4 ± 1.2 ab | 73.9 ± 4.7 b | 61.4 ± 0.9 a | 0.18 ± 0.03 ab | 9.7 ± 0.4 ab |
| C14 | 226 ± 6 a | 24.0 ± 0.3 d | 0.76 ± 0.01 ab | 79.3 ± 1.0 ab | 76.5 ± 3.6 b | 61.1 ± 0.8 a | 0.18 ± 0.02 ab | 9.0 ± 0.3 a |
| 358 | 204 ± 6 a | 22.2 ± 0.3 bc | 0.77 ± 0.01 ab | 80.4 ± 1.1 ab | 72.3 ± 2.8 b | 60.5 ± 0.8 a | 0.13 ± 0.01 a | 9.3 ± 0.3 a |
| 437 | 235 ± 7 a | 23.2 ± 0.3 cd | 0.78 ± 0.01 b | 80.5 ± 1.2 ab | 72.3 ± 3.5 b | 60.6 ± 0.9 a | 0.18 ± 0.03 ab | 9.1 ± 0.4 a |
| p | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.536 | <0.001 | <0.001 |
| Total | 286 ± 8 | 21.9 ± 0.3 | 0.76 ± 0.01 | 79.6 ± 0.4 | 70.9 ± 1.2 | 61.0 ± 0.3 | 0.17 ± 0.01 | 10.4 ± 0.2 |
| Measurement Date | d (mm−2) | SL (μm) | SW/SL | RWCc (%) | tc (min) | Mc (%) | Ec (mmol m−2 s−1) | SLA (m2 kg−1) |
|---|---|---|---|---|---|---|---|---|
| 01 Feb. 2015 | 278 ± 6 bcd | 21.6 ± 0.3 bcd | 0.74 ± 0.01 a | 73.2 ± 1.2 a | 86.3 ± 3.3 c | 58.7 ± 0.6 ab | 0.16 ± 0.01 abc | 11.5 ± 0.3 c |
| 01 May 2015 | 287 ± 7 cd | 22.8 ± 0.3 de | 0.78 ± 0.01 d | 82.9 ± 0.8 c | 81.3 ± 4.3 bc | 60.4 ± 0.6 bc | 0.12 ± 0.01 a | 10.1 ± 0.3 bc |
| 25 July 2015 | 276 ± 10 abcd | 20.7 ± 0.4 bc | 0.78 ± 0.01 bcd | 79.2 ± 1.7 abc | 50.0 ± 2.7 a | 59.4 ± 1.5 abcd | 0.22 ± 0.04 abc | 9.9 ± 0.7 abc |
| 01 Nov. 2015 | 247 ± 8 abc | 18.4 ± 0.4 a | 0.75 ± 0.01 abc | 81.4 ± 2.5 abc | 49.0 ± 1.5 a | 67.7 ± 1.1 f | 0.24 ± 0.03 abc | 14.4 ± 0.6 d |
| 01 Feb. 2016 | 247 ± 7 ab | 22.8 ± 0.3 cde | 0.76 ± 0.01 abcd | 80.8 ± 0.8 c | 66.3 ± 1.6 b | 62.4 ± 0.6 cd | 0.16 ± 0.02 abc | 10.6 ± 0.4 bc |
| 01 May 2016 | 276 ± 10 abcd | 24.2 ± 0.3 e | 0.77 ± 0.01 bcd | 82.6 ± 0.7 c | 64.1 ± 2.7 b | 56.5 ± 0.4 a | 0.14 ± 0.01 abc | 8.1 ± 0.2 a |
| 25 July 2016 | 299 ± 8 de | 26.1 ± 0.3 f | 0.78 ± 0.01cd | 80.5 ± 2.0 abc | 74.7 ± 2.5 bc | 60.8 ± 0.9 bce | 0.18 ± 0.05 abc | 9.2 ± 0.3 ab |
| 01 Nov. 2016 | 229 ± 6 a | 20.3 ± 0.3 ab | 0.75 ± 0.01 abc | 79.4 ± 1.2 bc | 70.2 ± 3.4 bc | 64.6 ± 0.9 def | 0.22 ± 0.02 c | 10.7 ± 0.4 bc |
| 01 Feb. 2017 | 283 ± 7 bcd | 20.2 ± 0.3 ab | 0.75 ± 0.01 ab | 75.5 ± 0.9 ab | 73.9 ± 3.0 bc | 64.5 ± 0.5 df | 0.18 ± 0.02 bc | 11.8 ± 0.4 cd |
| 01 May 2017 | 344 ± 9 e | 21.8 ± 0.3 bcd | 0.77 ± 0.01 bcd | 81.8 ± 0.9 c | 77.7 ± 4.8 bc | 57.3 ± 0.7 ab | 0.12 ± 0.01 ab | 8.3 ± 0.3 a |
| p | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Total | 286 ± 8 | 21.9 ± 0.3 | 0.76 ± 0.01 | 79.6 ± 0.4 | 70.9 ± 1.2 | 61.0 ± 0.3 | 0.17 ± 0.01 | 10.4 ± 0.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carignato, A.; Vázquez-Piqué, J.; Tapias, R.; Ruiz, F.; Fernández, M. Variability and Plasticity in Cuticular Transpiration and Leaf Permeability Allow Differentiation of Eucalyptus Clones at an Early Age. Forests 2020, 11, 9. https://doi.org/10.3390/f11010009
Carignato A, Vázquez-Piqué J, Tapias R, Ruiz F, Fernández M. Variability and Plasticity in Cuticular Transpiration and Leaf Permeability Allow Differentiation of Eucalyptus Clones at an Early Age. Forests. 2020; 11(1):9. https://doi.org/10.3390/f11010009
Chicago/Turabian StyleCarignato, André, Javier Vázquez-Piqué, Raúl Tapias, Federico Ruiz, and Manuel Fernández. 2020. "Variability and Plasticity in Cuticular Transpiration and Leaf Permeability Allow Differentiation of Eucalyptus Clones at an Early Age" Forests 11, no. 1: 9. https://doi.org/10.3390/f11010009
APA StyleCarignato, A., Vázquez-Piqué, J., Tapias, R., Ruiz, F., & Fernández, M. (2020). Variability and Plasticity in Cuticular Transpiration and Leaf Permeability Allow Differentiation of Eucalyptus Clones at an Early Age. Forests, 11(1), 9. https://doi.org/10.3390/f11010009