Timeline of Leaf and Cambial Phenology in Relation to Development of Initial Conduits in Xylem and Phloem in Three Coexisting Sub-Mediterranean Deciduous Tree Species


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Description
2.2. Xylem and Phloem Formation
2.3. Statistical Analysis
3. Results
3.1. Timing of Leaf Phenology and Seasonal Radial Growth
3.2. Development of Initial Conduits in Relation to Leaf Phenology
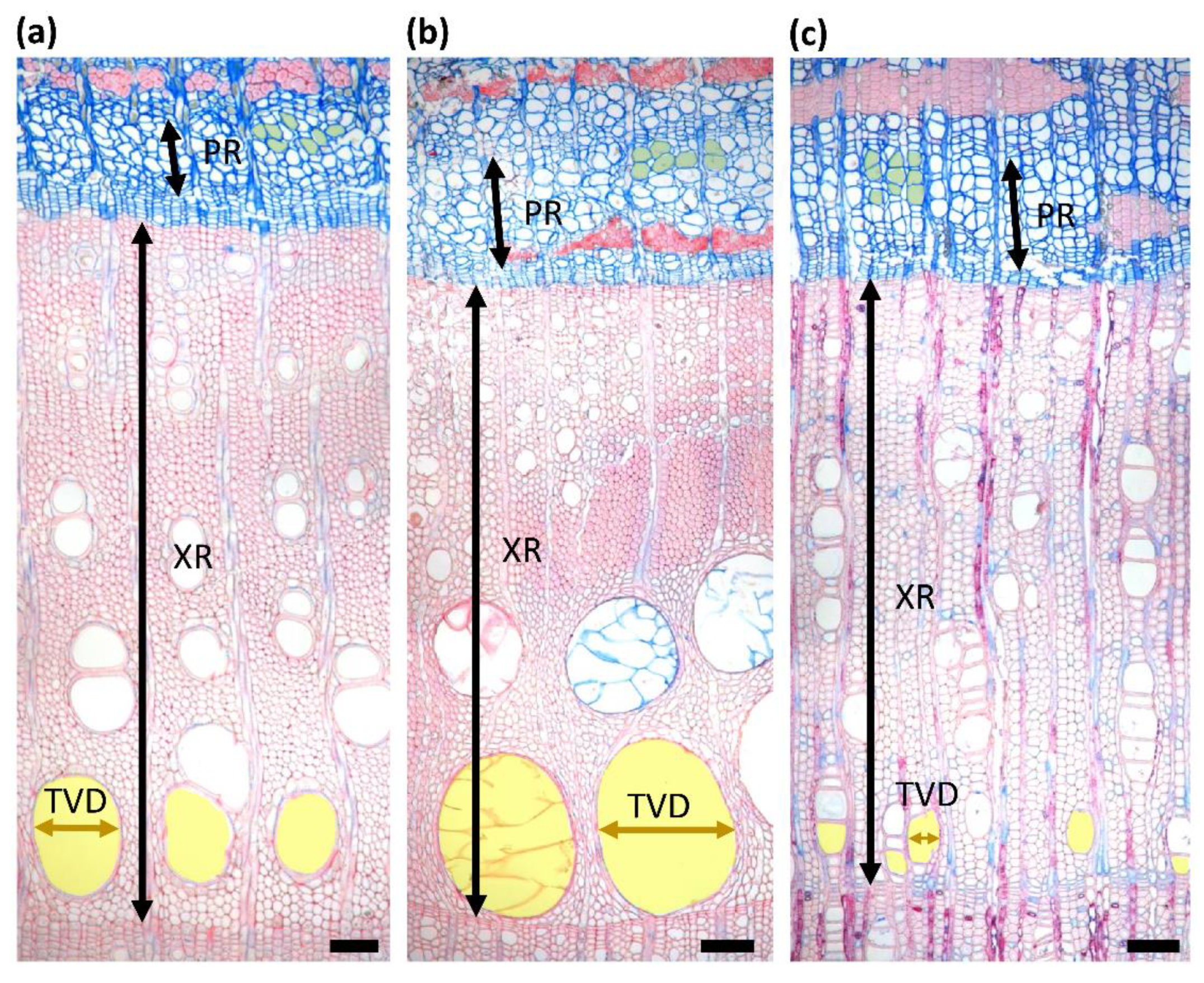
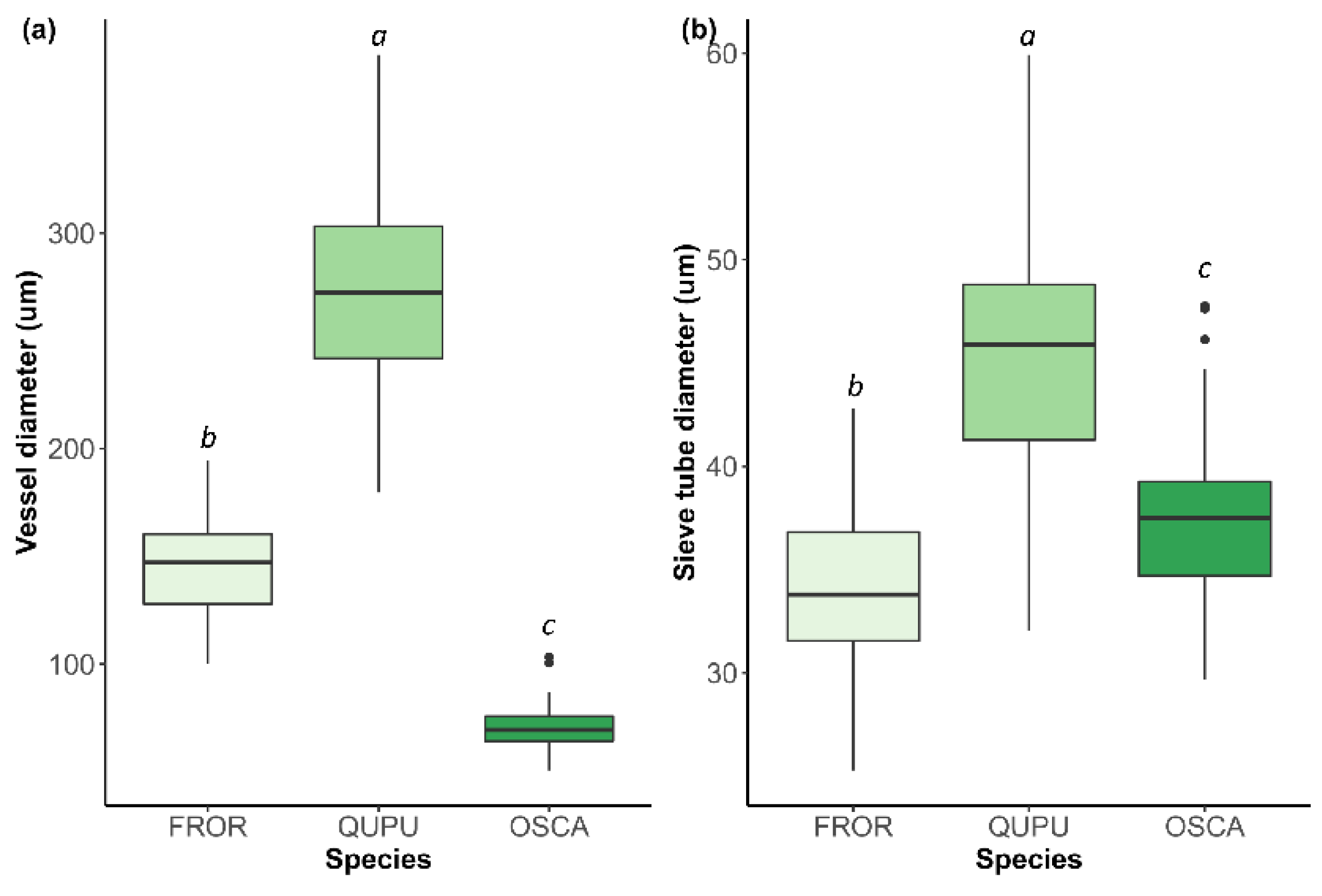
3.3. Size of Initial Earlywood Vessels and Early Phloem Sieve Tubes
4. Discussion
4.1. Widths of Annual Increments in Relation to Phenology and Weather Conditions
4.2. Linkage between Leaf Phenology and Development of Initial Earlywood Vessels
4.3. Characteristics of Initial Earlywood Vessels
4.4. Linkage between Leaf Phenology and Development and Size of Initial Early Phloem Sieve Tubes
4.5. Hormonal Control of Xylem and Phloem Development
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-KÜBler, K.; Bissolli, P.; BraslavskÁ, O.G.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Rossi, S.; Anfodillo, T.; Čufar, K.; Cuny, H.E.; Deslauriers, A.; Fonti, P.; Frank, D.; Gričar, J.; Gruber, A.; King, G.M.; et al. A meta-analysis of cambium phenology and growth: Linear and non-linear patterns in conifers of the northern hemisphere. Ann. Bot. 2013, 112, 1911–1920. [Google Scholar] [CrossRef] [PubMed]
- Chuine, I. Why does phenology drive species distribution? Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3149–3160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stucky, B.J.; Guralnick, R.; Deck, J.; Denny, E.G.; Bolmgren, K.; Walls, R. The Plant Phenology Ontology: A New Informatics Resource for Large-Scale Integration of Plant Phenology Data. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Delpierre, N.; Vitasse, Y.; Chuine, I.; Guillemot, J.; Bazot, S.; Rutishauser, T.; Rathgeber, C.B.K. Temperate and boreal forest tree phenology: From organ-scale processes to terrestrial ecosystem models. Ann. For. Sci. 2016, 73, 5–25. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Yang, B.; Shishov, V.; Rossi, S.; Bräuning, A.; Ljungqvist, F.C.; Grießinger, J. Relationships between Wood Formation and Cambium Phenology on the Tibetan Plateau during 1960–2014. Forests 2018, 9, 86. [Google Scholar] [CrossRef] [Green Version]
- Buras, A.; Menzel, A. Projecting Tree Species Composition Changes of European Forests for 2061–2090 Under RCP 4.5 and RCP 8.5 Scenarios. Front. Plant Sci. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decuyper, M.; Chávez, R.O.; Čufar, K.; Estay, S.A.; Clevers, J.G.P.W.; Prislan, P.; Gričar, J.; Črepinšek, Z.; Merela, M.; de Luis, M.; et al. Spatio-temporal assessment of beech growth in relation to climate extremes in Slovenia–An integrated approach using remote sensing and tree-ring data. Agric. For. Meteorol. 2020, 287, 107925. [Google Scholar] [CrossRef]
- Park, J.Y.; Muller-Landau, H.C.; Lichstein, J.W.; Rifai, S.W.; Dandois, J.P.; Bohlman, S.A. Quantifying Leaf Phenology of Individual Trees and Species in a Tropical Forest Using Unmanned Aerial Vehicle (UAV) Images. Remote Sens. 2019, 11, 1534. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Yoda, K.; Suzuki, H. Phenological comparison on the onset of vessel formation between ring-porous and diffuse-porous deciduous trees in a Japanese temperate forest. Iawa J. 1996, 17, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Sass-Klaassen, U.; Sabajo, C.R.; den Ouden, J. Vessel formation in relation to leaf phenology in pedunculate oak and European ash. Dendrochronologia 2011, 29, 171–175. [Google Scholar] [CrossRef]
- Takahashi, S.; Okada, N.; Nobuchi, T. Relationship between the timing of vessel formation and leaf phenology in ten ring-porous and diffuse-porous deciduous tree species. Ecol. Res. 2013, 28, 615–624. [Google Scholar] [CrossRef] [Green Version]
- Kitin, P.; Funada, R. Earlywood vessels in ring-porous trees become functional for water transport after bud burst and before the maturation of the current-year leaves. Int. Assoc. Wood Anat. 2016, 37, 315. [Google Scholar] [CrossRef]
- Puchałka, R.; Koprowski, M.; Gričar, J.; Przybylak, R. Does tree-ring formation follow leaf phenology in Pedunculate oak (Quercus robur L.)? Eur. J. For. Res. 2017, 136, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Dox, I.; Gričar, J.; Marchand, L.J.; Leys, S.; Zuccarini, P.; Geron, C.; Prislan, P.; Mariën, B.; Fonti, P.; Lange, H.; et al. Timeline of autumn phenology in temperate deciduous trees. Tree Physiol. 2020, 40, 1001–1013. [Google Scholar] [CrossRef] [PubMed]
- Marchand, L.J.; Dox, I.; Gričar, J.; Prislan, P.; Leys, S.; Van den Bulcke, J.; Fonti, P.; Lange, H.; Matthysen, E.; Peñuelas, J.; et al. Inter-individual variability in spring phenology of temperate deciduous trees depends on species, tree size and previous year autumn phenology. Agric. For. Meteorol. 2020, 290, 108031. [Google Scholar] [CrossRef]
- Pérez-de-Lis, G.; Rossi, S.; Vázquez-Ruiz, R.A.; Rozas, V.; García-González, I. Do changes in spring phenology affect earlywood vessels? Perspective from the xylogenesis monitoring of two sympatric ring-porous oaks. New Phytol. 2016, 209, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Prislan, P.; Gričar, J.; de Luis, M.; Smith, K.T.; Čufar, K. Phenological variation in xylem and phloem formation in Fagus sylvatica from two contrasting sites. Agric. For. Meteorol. 2013, 180, 142–151. [Google Scholar] [CrossRef]
- Gričar, J.; Lavrič, M.; Ferlan, M.; Vodnik, D.; Eler, K. Intra-annual leaf phenology, radial growth and structure of xylem and phloem in different tree parts of Quercus pubescens. Eur. J. For. Res. 2017, 136, 625–637. [Google Scholar] [CrossRef]
- Savage, J.A. It’s all about timing—or is it? Exploring the potential connection between phloem physiology and whole plant phenology. Am. J. Bot. 2020, 107, 848–851. [Google Scholar] [CrossRef]
- Ferlan, M.; Eler, K.; Simončič, P.; Batič, F.; Vodnik, D. Carbon and water flux patterns of a drought-prone mid-succession ecosystem developed on abandoned karst grassland. Agric. Ecosyst. Environ. 2016, 220, 152–163. [Google Scholar] [CrossRef]
- Rossi, S.; Anfodillo, T.; Menardi, R. Trephor: A new tool for sampling microcores from tree stems. Iawa J. 2006, 27, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Quinn, G.K.M. Experimental Design and Data Analysis for Biologists, 1st ed.; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Skene, D.S. The kinetics of tracheid development in Tsuga canadensis Carr and its relation to tree vigour. Ann. Bot. 1972, 36, 179–187. [Google Scholar] [CrossRef]
- Zweifel, R.; Zimmermann, L.; Zeugin, F.; Newbery, D.M. Intra-annual radial growth and water relations of trees: Implications towards a growth mechanism. J. Exp. Bot. 2006, 57, 1445–1459. [Google Scholar] [CrossRef] [Green Version]
- Gričar, J.; Zavadlav, S.; Jyske, T.; Lavrič, M.; Laakso, T.; Hafner, P.; Eler, K.; Vodnik, D. Effect of soil water availability on intra-annual xylem and phloem formation and non-structural carbohydrate pools in stem of Quercus pubescens. Tree Physiol. 2018, 39, 222–233. [Google Scholar] [CrossRef]
- Taneda, H.; Sperry, J.S. A case-study of water transport in co-occurring ring- versus diffuse-porous trees: Contrasts in water-status, conducting capacity, cavitation and vessel refilling. Tree Physiol. 2008, 28, 1641–1651. [Google Scholar] [CrossRef] [Green Version]
- Barbaroux, C.; Bréda, N.; Dufrêne, E. Distribution of above-ground and below-ground carbohydrate reserves in adult trees of two contrasting broad-leaved species (Quercus petraea and Fagus sylvatica). New Phytol. 2003, 157, 605–615. [Google Scholar] [CrossRef] [Green Version]
- Michelot, A.; Simard, S.; Rathgeber, C.; Dufrêne, E.; Damesin, C. Comparing the intra-annual wood formation of three European species (Fagus sylvatica, Quercus petraea and Pinus sylvestris) as related to leaf phenology and non-structural carbohydrate dynamics. Tree Physiol. 2012, 32, 1033–1045. [Google Scholar] [CrossRef] [Green Version]
- Tyree, M.T.; Zimmermann, M.H. Xylem Structure and the Ascent of Sap, 2nd ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2010. [Google Scholar]
- Cochard, H.; Tyree, M.T. Xylem dysfunction in Quercus: Vessel sizes, tyloses, cavitation and seasonal changes in embolism. Tree Physiol 1990, 6, 393–407. [Google Scholar] [CrossRef] [Green Version]
- Kiorapostolou, N.; Petit, G. Similarities and differences in the balances between leaf, xylem and phloem structures in Fraxinus ornus along an environmental gradient. Tree Physiol. 2018, 39, 234–242. [Google Scholar] [CrossRef]
- Sevanto, S.; Ryan, M.; Dickman, L.T.; Derome, D.; Patera, A.; Defraeye, T.; Pangle, R.E.; Hudson, P.J.; Pockman, W.T. Is desiccation tolerance and avoidance reflected in xylem and phloem anatomy of two coexisting arid-zone coniferous trees? Plantcell Environ. 2018, 41, 1551–1564. [Google Scholar] [CrossRef] [PubMed]
- Barbaroux, C.; Bréda, N. Contrasting distribution and seasonal dynamics of carbohydrate reserves in stem wood of adult ring-porous sessile oak and diffuse-porous beech trees. Tree Physiol. 2002, 22, 1201–1210. [Google Scholar] [CrossRef]
- Spicer, R. Symplasmic networks in secondary vascular tissues: Parenchyma distribution and activity supporting long-distance transport. J. Exp. Bot. 2014, 65, 1829–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aloni, R. Ecophysiological implications of vascular differentiation and plant evolution. Trees 2015, 29, 1–16. [Google Scholar] [CrossRef]
- Aloni, R. Differentiation of Vascular Tissues. Annu. Rev. Plant Physiol. 1987, 38, 179–204. [Google Scholar] [CrossRef]
- Aloni, R. Foliar and axial aspects of vascular differentiation: Hypotheses and evidence. J. Plant Growth Regul. 2001, 20, 12. [Google Scholar] [CrossRef] [Green Version]
- Aloni, R.; Alexander, J.D.; Tyree, M.T. Natural and experimentally altered hydraulic architecture of branch junctions in Acer saccharum Marsh. and Quercus velutina Lam. trees. Trees 1997, 11, 255–264. [Google Scholar] [CrossRef]
- Dayan, J.; Voronin, N.; Gong, F.; Sun, T.-p.; Hedden, P.; Fromm, H.; Aloni, R. Leaf-Induced Gibberellin Signaling Is Essential for Internode Elongation, Cambial Activity, and Fiber Differentiation in Tobacco Stems. Plant Cell 2012, 24, 66–79. [Google Scholar] [CrossRef] [Green Version]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.-J.; Nabuurs, G.-J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Todaro, L.; Rita, A.; Negro, F.; Moretti, N.; Saracino, A.; Zanuttini, R. Behavior of pubescent oak (Quercus pubescens Willd.) wood to different thermal treatments. Iforest Biogeosci. For. 2015, 8, 748–755. [Google Scholar] [CrossRef] [Green Version]
- Humar, M.; Balzano, A.; Grbec, S.; Gričar, J.; Kržišnik, D.; Lesar, B.; Vek, V. Investigation of the material resistance and moisture performance of pubescent oak (Quercus pubescens). Holzforschung 2020. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraxinus ornus | Quercus pubescens | Ostrya carpinifolia | |
|---|---|---|---|
| DBH (cm) 1 | 13.5 ± 5.4 | 18.9 ± 4.7 | 14.3 ± 2.8 |
| H (m) 2 | 8.9 ± 1.2 | 9.8 ± 1.0 | 10.0 ± 1.2 |
| Phenological Phases (DOY ± Standard Deviation) | |||
|---|---|---|---|
| Fraxinus ornus | Quercus pubescens | Ostrya carpinifolia | |
| Onset of cambial cell production | 59.33 ± 3.61 b | 67.50 ± 3.83 a | 71.00 ± 0.00 a |
| Swollen buds | 101.00 ± 3.10 a | 102.00 ± 2.45 a | 77.00 ± 0.00 b |
| Final size of initial early phloem sieve tube | 106.33 ± 3.27 a | 97.00 ± 3.27 b | 99.00 ± 6.57 a,b |
| Leaf emergence | 123.50 ± 6.12 a | 119.33 ± 4.08 b | 85.00 ± 0.00 c |
| Transition from early phloem to late phloem | 130.00 ± 6.26 b | 147.00 ± 3.29 a | 133.50 ± 3.84 b |
| Transition from earlywood to latewood | 152.50 ± 2.74 b | 134.67 ± 3.61 a | / |
| Full leaf unfolding | 146.00 ± 3.10 a | 140.83 ± 2.86 b | 131.50 ± 3.83 c |
| Mature initial earlywood vessel | 137.00 ± 0.00 b | 133.50 ± 3.84 b | 159.50 ± 4.93 a |
| End of cambial cell production | 191.83 ± 11.96 b | 208.33 ± 3.61 a | 205.33 ± 7.17 a,b |
| Autumn leaf coloring | 298.33 ± 8.26 c | 309.00 ± 0.00 b | 322.00 ± 0.00 a |
| Xylem and Phloem Characteristics | |||
|---|---|---|---|
| Fraxinus ornus | Quercus pubescens | Ostrya carpinifolia | |
| Xylem ring width (µm) | 1136.7 ± 550.8 b | 737.2 ± 269.9 a | 814.41 ± 526.09 a |
| Phloem ring width (µm) | 158.3 ± 30.4 b | 236.8 ± 37.4 a | 213.1 ± 73.3 b |
| IEV diameter (µm) | 144.1 ± 20.7 b | 271.9 ± 44.8 a | 70.3 ± 10.2 c |
| IEV area (µm2) | 18,011.1 ± 4629.5 b | 61,007.9 ± 15,713.7 a | 5216.7 ± 1406.7 c |
| Sieve tube diameter (µm) | 34.5 ± 4.1 b | 45.4 ± 6.4 a | 37.5 ± 4.3 c |
| Sieve tube area (µm2) | 852.7 ± 166.6 b | 1213.4 ± 222.8 a | 1038.4 ± 151.5 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gričar, J.; Vedenik, A.; Skoberne, G.; Hafner, P.; Prislan, P. Timeline of Leaf and Cambial Phenology in Relation to Development of Initial Conduits in Xylem and Phloem in Three Coexisting Sub-Mediterranean Deciduous Tree Species. Forests 2020, 11, 1104. https://doi.org/10.3390/f11101104
Gričar J, Vedenik A, Skoberne G, Hafner P, Prislan P. Timeline of Leaf and Cambial Phenology in Relation to Development of Initial Conduits in Xylem and Phloem in Three Coexisting Sub-Mediterranean Deciduous Tree Species. Forests. 2020; 11(10):1104. https://doi.org/10.3390/f11101104
Chicago/Turabian StyleGričar, Jožica, Andreja Vedenik, Gregor Skoberne, Polona Hafner, and Peter Prislan. 2020. "Timeline of Leaf and Cambial Phenology in Relation to Development of Initial Conduits in Xylem and Phloem in Three Coexisting Sub-Mediterranean Deciduous Tree Species" Forests 11, no. 10: 1104. https://doi.org/10.3390/f11101104
APA StyleGričar, J., Vedenik, A., Skoberne, G., Hafner, P., & Prislan, P. (2020). Timeline of Leaf and Cambial Phenology in Relation to Development of Initial Conduits in Xylem and Phloem in Three Coexisting Sub-Mediterranean Deciduous Tree Species. Forests, 11(10), 1104. https://doi.org/10.3390/f11101104